Contents
What is parthenogenesis
Parthenogenesis (parthenos = virgin, genesis = creation) can be defined as the production of an embryo from a female gamete (female egg) without any genetic contribution from a male gamete (male sperm), with or without the eventual development into an adult. Parthenogenesis is the development of offspring from unfertilized eggs. Parthenogenesis is the artificial activation of an egg in the absence of a sperm; the egg begins to divide as if it has been fertilized. Parthenogenesis naturally occurs in a variety of plant and animal species, particularly in lower plants such as mosses and algae and species-rich invertebrate groups such as insects, nematodes, and crustaceans, but also in about 10% of the fern and 1% of the flowering plant species, and as rare examples in vertebrates 1. In plants, parthenogenesis usually is found in combination with apomeiosis (the omission of meiosis) and pseudogamous or autonomous (with or without central cell fertilization) endosperm formation, together known as apomixis (clonal seed production).
Parthenogenesis is a reproduction strategy common in some jawed vertebrate species such as the whiptail lizard (Aspidoscelis uniparens) 2, in which no sperm is involved to trigger embryonic development from the oocyte and the female generates an offspring with no paternal inheritance. In mammals, parthenogenesis is not a natural form of reproduction, as the birth of an offspring is considered not possible 3.
Parthenogenesis is distinct from asexual reproduction since it involves the production of egg cells. Parthenogenesis is a normal method of reproduction in many lower organisms, but does not lead to viable mammalian offspring. Parthenogenetic development can proceed by various routes depending on whether meiosis has occurred or has been supressed, in which case the egg develops as as result of mitotic divisions. Whenever sex is determined by chromosome constitution, parthenogenetic offspring, in the absence of effective meiosis, all will be, mostly female. In birds, however, the offspring are male as in this case females are the heterogametic sex. In bees, males originate by haploid parthenogenesis while diploid females are produced by fertilization in the normal way. Other aphids, such as greenfly (Hemiptera) have generations which alternate between parthenogenesis and fertilization, so called cyclical parthenogenesis. The formation of female parthenogenetic offspring is widespread among many order of insects. For example in Drosophila parthenogenetica, a small proportion of eggs laid by virgin females develop to produce viable adults. Another example is the parthenogenetic grasshopper Warramaba virgo, a species which consists of females only. Parthenogenesis is also successful in some Crustaceae such as the brine shrimp, Artemia salina.
There are a number of genetically different types of parthenogenesis, ranging from gamete duplication, which immediately eliminates all heterozygosity, to apomixis and prezygotic doubling, which both preserve all the heterozygosity that was present in the mother. The existence of successful parthenogenetic lineages demonstrates that sex is not necessary for reproduction and raises the question of why sex exists. Parthenogenetic lineages are often successful in the short term but almost never persist in the evolutionary long term. An influential hypothesis holds that sexual populations persist longer than parthenogenetic populations because they evolve more rapidly in response to environmental challenges such as rapidly evolving parasites and pathogens.
However, parthenogenetic activation of mammalian eggs (oocytes) by artificial manipulation results in early embryonic development and in some instances fetal early development can be achieved (mouse forelimb stage E9.5) stage 4. No further development is achieved due to the lack of expression of imprinted paternal genes necessary for the establishment of a functional placenta 5. On the other hand, genetic modification technologies have made the creation and birth of fertile animals from mouse parthenogenetically activated oocytes possible 6.
Parthenogenetic activation of human eggs (oocytes) obtained from infertility treatments has gained new interest in recent years as an alternative approach to create human parthenogenetic embryos as a source of cells for research in areas such as assisted reproduction technologies (ART) itself, somatic cell, and nuclear transfer experiments and for derivation of clinical grade pluripotent embryonic stem cells for regenerative medicine 3. Different activating methods have been tested on human and nonhuman oocytes, with varying degrees of success in terms of parthenote generation rates, embryo development stem cell derivation rates. Success in achieving a standardized artificial activation methodology for human oocytes and the subsequent potential therapeutic gain obtained from these embryos depends mainly on the availability of gametes donated from infertility treatments.
Renewed interest in parthenogenetic activation of mammalian oocytes has come about for scientific, medical, and economic reasons. First of all, experimental work involving parthenogenetic embryos circumvents the ethical and legal problems concerning the use of human embryos generated for reproductive purposes. Parthenotes may be employed in different research areas related to ART, studies on human pluripotent stem cells or basic science uncovering regulatory mechanisms that command human embryonic development and cloning experiments using somatic cell or nuclear transfer in mammalian oocytes.
Last but not least, the creation of clinical grade parthenogenetic human embryonic stem cell (hpESC) lines have the potential to benefit a considerable large number of patients, when used in cell therapies, with a reduced risk of transplant rejection. However, to this end, efforts should be directed to clinicians to participate in research projects. The establishment of parthenogenetic clinical grade human embryonic stem cell lines for use in cell or tissue therapies will only be possible, if research laboratories obtain enough biological material from assisted reproduction technologies (ART) centers to evaluate activation strategies, derivation, and culture media adequate for future therapeutic use of parthenogenetic human embryonic stem cell (hpESC) lines. Furthermore, well trained committed staff, with good practice and knowledge of basic embryology, is paramount for the conscious and successful use of spare oocytes and embryos obtained from assisted reproduction centers.
Parthenogenetic activation methodologies
Biochemistry of Oocyte Activation
Arrested nonfertilized metaphase II (MII) oocytes will remain at this stage until a stimulus, which may come from the fertilizing spermatozoon or from an artificial agent (Figure 1), triggers intracytoplasmic Ca2+ rises and meiosis resumption. Intracellular Ca2+ oscillations will inhibit the action of the metaphase promoting factor (MPF) and the cytostatic factor (CSF) and lead to metaphase/anaphase transition, segregation of sister chromatids, and extrusion of second polar body. The importance of these Ca2+ transients for oocyte activation was shown by the prevention of the intracellular elevation of Ca2+ after sperm penetration by preloading the oocytes with the Ca2+ chelator BAPTA 1-AM 7. In the absence of any intracellular Ca2+ increase, activation and subsequent embryo development failed to occur. In mammals, intracellular Ca2+ transients are triggered by a putative sperm factor, the testis specific phospholipase C-zeta (plc-zeta), or the newly described postacrosomal sheath WW domain-binding protein PAWP released by the sperm during the normal fertilization process 8. Regardless of the exact nature of the sperm factor responsible for eliciting the Ca2+ oscillations, its presence is paramount for successful oocyte activation and embryonic development. The activating agents used for parthenogenetic activation may mimic the sperm bound stimuli to trigger Ca2+ release from the endoplasmic reticulum, exit from meiotic arrest, and start embryonic development.
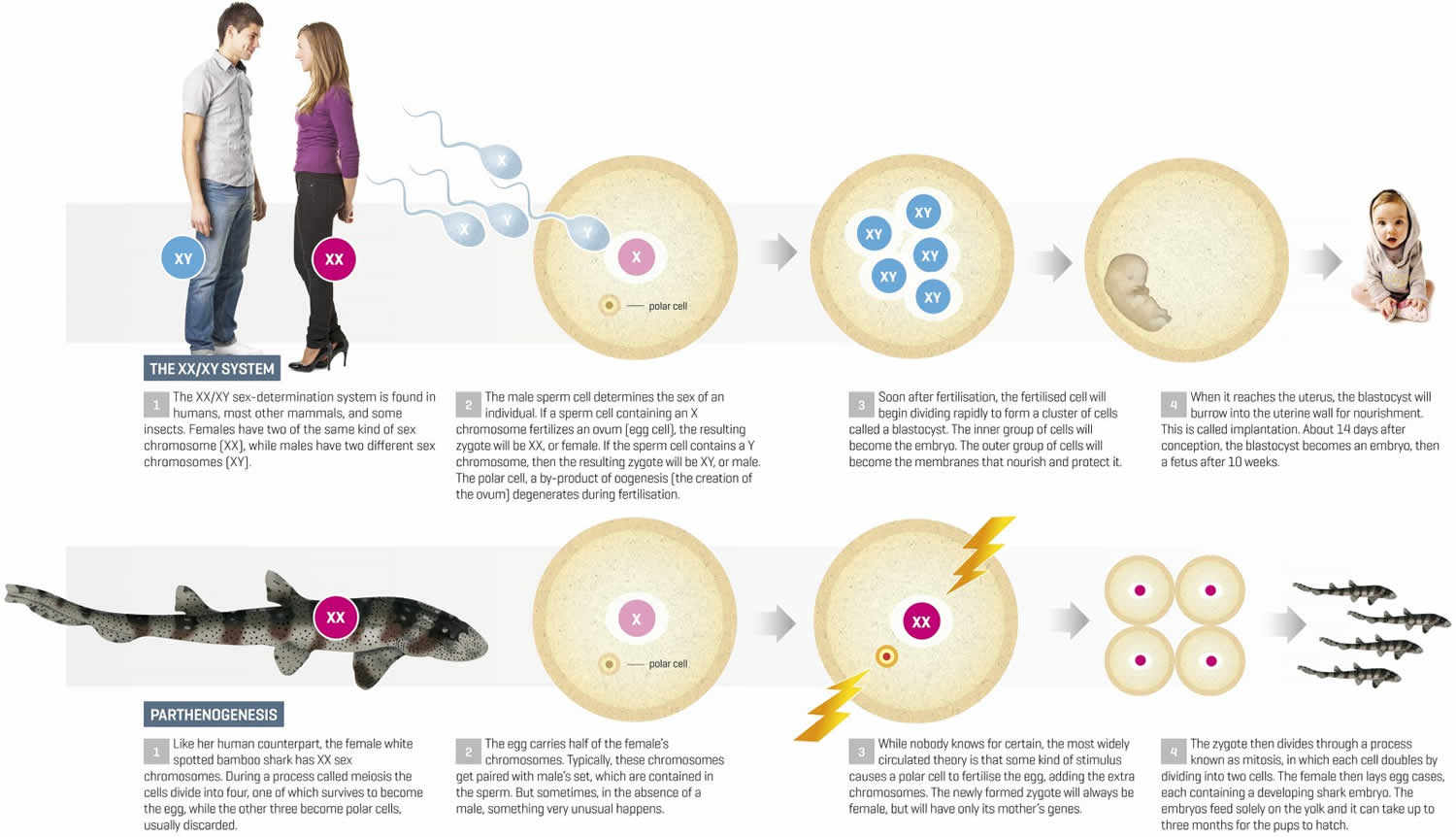
Figure 1. Types of parthenogenesis
Techniques of Parthenogenetic Activation
A wide range of artificial stimuli have been employed as agents to trigger the activation process in oocytes of several animal species. All the techniques which have been described for artificial activation can be divided into two major groups, depending on the nature of the activating stimulus, that is, chemical or physical agents. Except for Sr2+ 7, all other artificial agents that have been studied do not produce repetitive Ca2+ oscillations normally observed during fertilization. However, the single rise in intracellular Ca2+ that is produced by agents such as Ca2+ ionophore, ionomycin, or ethanol is adequate to trigger the meiotic resumption and cortical granule release. Specific oocyte activation protocols using strontium chloride (SrCl2) to induce continuous Ca2+ oscillations during exit from second meiosis and first embryonic mitosis seem to have a role in long-term embryonic events, such as the number of cells in the inner cell mass (ICM) and trophectoderm of the resulting blastocyst 9.
Despite the fact that various methods have been investigated for parthenogenetically activating oocytes through intracytoplasmic Ca2+ elevations, it seems that there is not a single method that activates oocytes from every species studied. Thus, parthenogenetic activation strategies have presented varying degrees of success, regarding activation rates and subsequent embryonic development according to the protocol employed and the species.
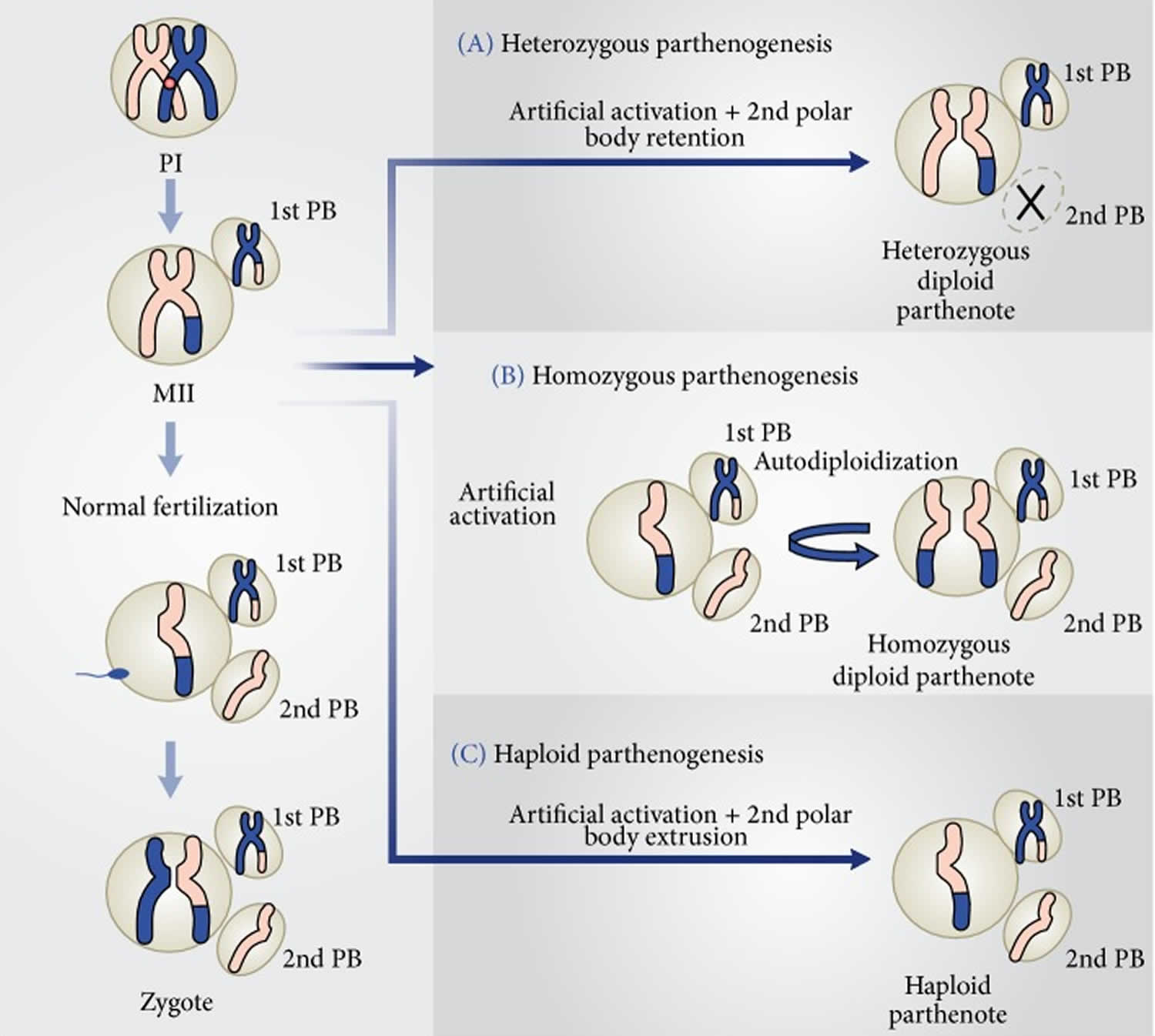
Figure 1 shows how embryos are generated after fertilization of an MII oocyte and the three possible parthenotes that may result according to the activation protocol employed to stimulate exit from MII, in the absence of a fertilizing spermatozoon.
In the first instance (Figure 1A), treatment with an activating agent such as SrCl2, ethanol, Ca2+ ionophore, or ionomycin is followed by another chemical, for instance, 6-DMAP (broad protein synthesis inhibitor) or cytochalasin B or D (inhibitors of actin filaments polymerization), which blocks second polar body (PB2) extrusion. Thus, the resulting parthenote is a “pseudodiploid” heterozygous embryo containing the two sister chromatids of each maternal chromosome present in the MII oocyte.
Alternative activation methodology may allow PB2 extrusion and the resulting parthenote containing a single copy of the sister chromatids undergoes spontaneous diploidization giving rise to a completely homozygous embryo (Figure 1B).
Last, the activating agent induces exit from MII and PB2 extrusion, without diploidization (Figure 1C). The result of this strategy is a haploid zygote, which develops into a haploid parthenote. However, in this instance, embryo development to the blastocyst stage is generally very low 3.
High oocyte activation and blastocyst formation rates were achieved, when mouse oocytes were exposed to Ca2+-free medium supplemented with 10 mM SrCl2 for 2 hours 10. However, similar success rates were not achieved when the same protocol was employed to activate human and bovine oocytes 11.
On the other hand, bovine oocytes presented high activation rates and development to the blastocyst stage, when exposed to the Ca2+ salt ionomycin for 5 minutes, followed by 2 mM 6-dimethylaminopurine (6-DMAP) for 3.5 hours 12. This last activating method has been employed to generate parthenogenetic bovine pluripotent embryonic stem cell, resulting in high rates of derivation and proliferation of primary embryonic stem cell colonies 13.
Activating Protocols for Fresh Human Oocytes
Human oocytes have been artificially activated mainly with the aim to generate blastocysts for the creation of parthenogenetic human embryonic stem cell (phESC) lines 14. Notably, parthenogenetic embryo development and embryonic stem cell derivation efficiencies are similar to those of sperm fertilized counterparts 15.
The first phESC lines were created using Ca2+ ionomycin together with 6-DMAP 16. In that study, fresh oocytes were obtained from shared oocyte donation cycles and the activation procedure took place few hours (3-4 hours) after collection. Twenty-three blastocysts (50%) were generated, from which six phESC lines were established. Mai et al. 15 described an activation methodology that combines electrical and chemical stimuli using ionomycin and 6-DMAP, for the artificial activation of fresh oocytes. Despite the fact that their parthenogenetic blastocyst rate was lower (21%) than the previous report 16, the inner cell mass of three blastocysts was used for the successful derivation of two new parthenogenetic stem cell lines. Blastocyst formation rates also vary considerably in ART cycles, depending on several factors such as the age of the patient, ovarian stimulation protocol, and culture conditions, among others. Thus, it is not surprising that reports on developmental rates of human parthenogenetic embryos vary between studies, as the age of the donors and the stimulation protocols may differ considerably, affecting the final blastocyst rates and consequently their inner cell mass quality.
Activation of Failure to Fertilize Human Oocytes
The detection of nonfertilized oocytes after insemination is not uncommon in ART treatments, both due to gamete immaturity and unknown factor that led to a failure to fertilize (FF). Most frequently, failure to fertilize occurs after in vitro fertilization (IVF). In this situation, the oocyte will remain arrested in MII. Rescue insemination by intracytoplasmic sperm injection (ICSI), few hours after detection of FF, has been proposed as a valid alternative to induce the creation of zygotes from the nonfertilized oocytes 17. There is, however, the underlying risk of chromosome abnormalities in the resulting embryo, mainly because of oocyte aging from the collection time to the moment of reinsemination by intracytoplasmic sperm injection (ICSI), which may occur 30 hours (or more) after oocyte pick-up. Also, the resulting clinical gestation rates vary considerably from center to center, which hampers rescue insemination to be used as a routine procedure in infertility treatments.
On the other hand, it has been shown that unwanted, failure to fertilize human oocytes retain their developmental potential and may generate viable blastocysts by exposure to ionomycin followed by 6-DMAP and a protein synthesis inhibitor, cyclohexamide 18. The embryos presented a similar gene expression profiling and developmental potential as normally fertilized ones making the unwanted artificially activated oocytes an alternative route for the generation of human embryonic stem cell lines.
A recent study 19 compared the efficacy of SrCl2 or Ca2+ ionophore on the activation rates of 3-day-old failure to fertilize human oocytes and observed that the treatment with the ionophore was more efficient in activating aged unfertilized oocytes and embryo development to the blastocyst stage. However, these reports on the activation of failure to fertilize oocytes do not reflect the real parthenogenetic embryonic development potential of the human oocyte as the studies mentioned earlier 20, because, in these last instances, there is a high probability that a paternal genome is present in the ooplasm, considering that IVF or ICSI was previously performed.
As an adjunct procedure in ART cycles, two intracellular Ca2+ releasing agents, SrCl2, and ionomycin have been used aiming at activating oocytes during fertilization. In a series of nine cases of couples who have had total fertilization failure or very low fertilization rates in previous cycles, Kyono et al. 21 exposed oocytes soon after insemination by ICSI to chemical stimulation using SrCl2. Fertilization rates increased from 21.7% in the previous cycles to 64.5% in the cycles using SrCl2 artificial activation. Six pregnancies were established, four of which went to term. Five healthy children were born and their 1-year followup did not show any physical or neurological abnormality. In another study, the same group of authors employed the Ca2+ ionophore to activate zygotes generated by ICSI using globozoospermic sperm 22. The alternative use of Ca2+ ionophore to activate the oocytes during fertilization and induce embryo development and gestation in cases of previous failure to fertilize of human oocytes was also reported with relatively good success rates by other groups 23.
The exposure to SrCl2 or Ca2+ ionophore may have induced the release and oscillations of intracellular Ca2+ needed for oocyte exit from the meiotic arrest and embryonic development 24. However, the putative epigenetic effects of this procedure demands caution and more research on the procedure. Also, the long-term followup of the children born by this methodology is necessary, before it can be considered as a routine methodology in human ART programs.
Oocyte Activation after In Vitro Maturation
In classical, stimulated ART cycles, immature germinal vesicle (GV) or metaphase I (MI) oocytes are commonly collected together with the mature MII. The immature oocytes may be submitted to in vitro maturation and ICSI to produce embryos for reproductive purposes. Resulting implantation and pregnancy rates are generally poor and there are controversies on whether to use these immature oocytes and embryos for reproductive purposes 25. However, attempts to in vitro mature GV or MI oocytes obtained from stimulated ovaries may yield MII oocytes and potentially parthenotes, under suitable in vitro maturation conditions. Liu et al. 26 have shown that cryopreserved immature germinal vesicle (GV) or metaphase I (MI) oocytes collected from stimulated cycles yield good rates of high quality blastocysts, after insemination by ICSI and exposure to the activating agent ethanol. Thus, immature oocytes collected from stimulated ovaries should not be neglected as an additional source of gametes for parthenogenetic activation. Research should focus on protocols devised specifically to improve their cytoplasmic and nuclear maturation to generate blastocysts with high quality inner cell masses.
On the other hand, IVM protocols to promote maturation of oocytes collected from unstimulated ovaries exist and they are employed in several fertility centers around the world, as an alternative to classical, stimulated cycles 27. The present results show that implantation and gestation rates after IVM are close to those obtained in classical stimulated cycles. Thus, surplus immature oocytes obtained from in vitro maturation cycles or from oophorectomy, in patients who undergo sex reassignment surgery 28, for instance, may represent an important source of gametes for artificial parthenogenetic activation and blastocyst formation after exposure to adequate in vitro maturation conditions.
- Grusz A. L. (2016). A current perspective on apomixis in ferns. J. Syst. Evol. 54 656–665. 10.1111/jse.12228[↩]
- Behavioral facilitation of reproduction in sexual and unisexual whiptail lizards. Crews D, Grassman M, Lindzey J. Proc Natl Acad Sci U S A. 1986 Dec; 83(24):9547-50.[↩]
- Bos-Mikich A, Bressan FF, Ruggeri RR, Watanabe Y, Meirelles FV. Parthenogenesis and Human Assisted Reproduction. Stem Cells Int. 2015;2016:1970843. doi:10.1155/2016/1970843 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4655294[↩][↩][↩][↩]
- Normal postimplantation development of mouse parthenogenetic embryos to the forelimb bud stage. Kaufman MH, Barton SC, Surani MA. Nature. 1977 Jan 6; 265(5589):53-5.[↩]
- Completion of mouse embryogenesis requires both the maternal and paternal genomes. McGrath J, Solter D. Cell. 1984 May; 37(1):179-83.[↩]
- Birth of parthenogenetic mice that can develop to adulthood. Kono T, Obata Y, Wu Q, Niwa K, Ono Y, Yamamoto Y, Park ES, Seo JS, Ogawa H. Nature. 2004 Apr 22; 428(6985):860-4.[↩]
- Repetitive calcium transients and the role of calcium in exocytosis and cell cycle activation in the mouse egg. Kline D, Kline JT. Dev Biol. 1992 Jan; 149(1):80-9.[↩][↩]
- PLCζ or PAWP: revisiting the putative mammalian sperm factor that triggers egg activation and embryogenesis. Kashir J, Nomikos M, Swann K, Lai FA. Mol Hum Reprod. 2015 May; 21(5):383-8.[↩]
- Meiotic and mitotic Ca2+ oscillations affect cell composition in resulting blastocysts. Bos-Mikich A, Whittingham DG, Jones KT. Dev Biol. 1997 Feb 1; 182(1):172-9.[↩]
- Calcium oscillations and protein synthesis inhibition synergistically activate mouse oocytes. Bos-Mikich A, Swann K, Whittingham DG. Mol Reprod Dev. 1995 May; 41(1):84-90.[↩]
- Bos-Mikich A., Ruggeri R., Rodrigues J. L., Oliveira N. P., Croco M., Prak P. H. Employment oocyte disposal as a source of blastocysts parthenotes to generate stem cell lines with clinical grade human embryonic. Jornal Brasileiro de Reprodução Assistida. 2011;15(3):38–40.[↩]
- The use of parthenotegenetic and IVF bovine blastocysts as a model for the creation of human embryonic stem cells under defined conditions. Ruggeri RR, Watanabe Y, Meirelles F, Bressan FF, Frantz N, Bos-Mikich A. J Assist Reprod Genet. 2012 Oct; 29(10):1039-43.[↩]
- Ruggeri R. R., Bressan F. F., Siqueira N. M., et al. Derivation and culture of putative parthenogenetic embryonic stem cells in new gelatin substrates modified with galactomannan. Macromolecular Research. 2014;22(10):1053–1058. doi: 10.1007/s13233-014-2151-5[↩]
- Brevini T. A. L., Gandolfi F. Parthenotes as a source of embryonic stem cells. Cell Proliferation. 2008;41(supplement 1):20–30. doi: 10.1111/j.1365-2184.2008.00485.x[↩]
- Mai Q., Yu Y., Li T., et al. Derivation of human embryonic stem cell lines from parthenogenetic blastocysts. Cell Research. 2007;17(12):1008–1019. doi: 10.1038/cr.2007.102[↩][↩]
- Revazova E. S., Turovets N. A., Kochetkova O. D., et al. Patient-specific stem cell lines derived from human parthenogenetic blastocysts. Cloning and Stem Cells. 2007;9(3):432–449. doi: 10.1089/clo.2007.0033[↩][↩]
- Amarin Z. O., Obeidat B. R., Rouzi A. A., Jallad M. F., Khader Y. S. Intracytoplasmic sperm injection after total conventional in-vitro fertilization failure. Saudi Medical Journal. 2005;26(3):411–415.[↩]
- Sneddon S. F., Desousa P. A., Arnesen R. E., Lieberman B. A., Kimber S. J., Brison D. R. Gene expression analysis of a new source of human oocytes and embryos for research and human embryonic stem cell derivation. Fertility and Sterility. 2011;95(4):1410–1415. doi: 10.1016/j.fertnstert.2010.08.050[↩]
- Liu Y., Han X.-J., Liu M.-H., et al. Three-day-old human unfertilized oocytes after in vitro fertilization/intracytoplasmic sperm injection can be activated by calcium ionophore A23187 or strontium chloride and develop to blastocysts. Cellular Reprogramming. 2014;16(4):276–280. doi: 10.1089/cell.2013.0081[↩]
- Ruggeri R. R., Watanabe Y., Meirelles F., Bressan F. F., Frantz N., Bos-Mikich A. The use of parthenotegenetic and IVF bovine blastocysts as a model for the creation of human embryonic stem cells under defined conditions. Journal of Assisted Reproduction and Genetics. 2012;29(10):1039–1043. doi: 10.1007/s10815-012-9866-z[↩]
- Kyono K., Kumagai S., Nishinaka C., et al. Birth and follow-up of babies born following ICSI using SrCl2 oocyte activation. Reproductive BioMedicine Online. 2008;17(1):53–58. doi: 10.1016/s1472-6483(10)60293-3[↩]
- Kyono K., Nakajo Y., Nishinaka C., et al. A birth from the transfer of a single vitrified-warmed blastocyst using intracytoplasmic sperm injection with calcium activation in a globozoospermic patient. Fertility and Sterility. 2009;91(3):e7–e11[↩]
- Check J. H., Summers-Chase D., Cohen R., Brasile D. Artificial oocyte activation with calcium ionophore allowed fertilization and pregnancy in a couple with long-term unexplained infertility where the female partner had diminished EGG reserve and failure to fertilize oocytes despite intracytoplasmic sperm injection. Clinical and Experimental Obstetrics & Gynecology. 2010;37(4):263–265[↩]
- Bos-Mikich A., Swann K., Whittingham D. G. Calcium oscillations and protein synthesis inhibition synergistically activate mouse oocytes. Molecular Reproduction and Development. 1995;41(1):84–90. doi: 10.1002/mrd.1080410113[↩]
- Shu Y., Gebhardt J., Watt J., Lyon J., Dasig D., Behr B. Fertilization, embryo development, and clinical outcome of immature oocytes from stimulated intracytoplasmic sperm injection cycles. Fertility and Sterility. 2007;87(5):1022–1027. doi: 10.1016/j.fertnstert.2006.08.110[↩]
- Liu Y., Cao Y. X., Zhang Z. G., Xing Q. Artificial oocyte activation and human failed-matured oocyte vitrification followed by in vitro maturation. Zygote. 2013;21(1):71–76. doi: 10.1017/s0967199411000530[↩]
- Strowitzki T. In vitro maturation (IVM) of human oocytes. Archives of Gynecology and Obstetrics. 2013;288(5):971–975. doi: 10.1007/s00404-013-3033-3[↩]
- Imesch P., Scheiner D., Xie M., et al. Developmental potential of human oocytes matured in vitro followed by vitrification and activation. Journal of Ovarian Research. 2013;6(1, article 30) doi: 10.1186/1757-2215-6-30[↩]

{kind=link}