Spinal cord
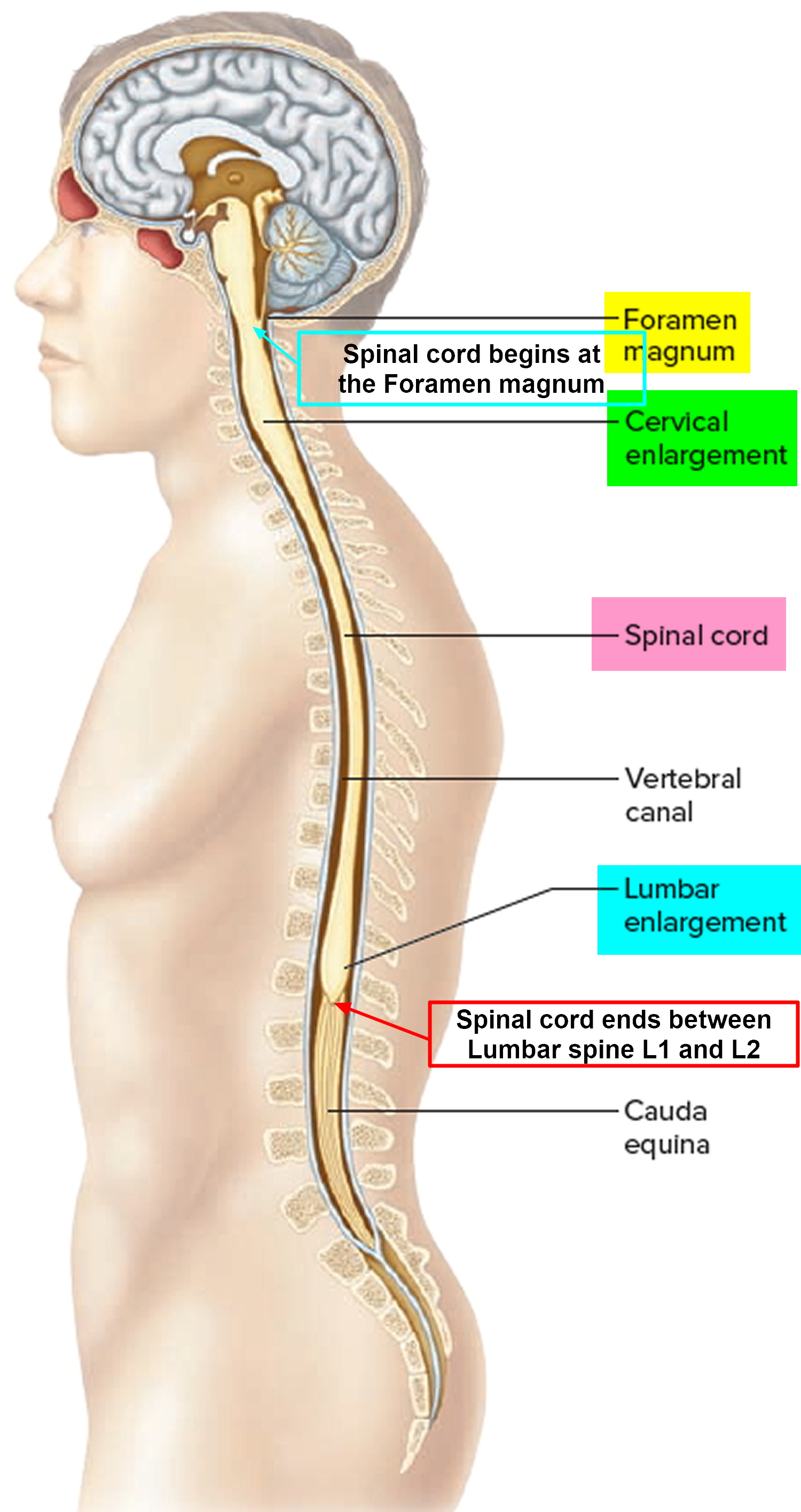
The spinal cord is a slender column of nervous tissue that passes downward from the brain into the vertebral canal. Although continuous with the brain, the spinal cord begins where nervous tissue leaves the cranial cavity at the level of the foramen magnum.
The spinal cord is not uniform in diameter along its length. In the neck region, a thickening in the spinal cord, called the cervical enlargement, occurs in the region associated with the origins of spinal nerves from the cervical spines to thoracic spine T1 giving rise to nerves to the upper limbs. A similar thickening in the lower back, the lumbosacral enlargement, occurs in the region associated with the origins of spinal nerves thoracic spine T 11 to Sacral S3 giving rise to nerves to the lower limbs. The spinal cord tapers to a point and ends near the intervertebral disc that separates the first (L1) and second lumbar (L2) vertebrae, adults, although it can end as high as thoracic vertebra T12 or as low as the disc between vertebrae lumbar vertebrae L2 and L3. From this point, nervous tissue, including axons of both motor and sensory neurons, extends downward to become spinal nerves at the remaining lumbar and sacral levels forming a structure called the cauda equina (horse’s tail).
The distal end of the cord (the conus medullaris) is cone shaped. A fine filament of connective tissue (the pial part of the filum terminale) continues inferiorly from the apex of the conus medullaris.
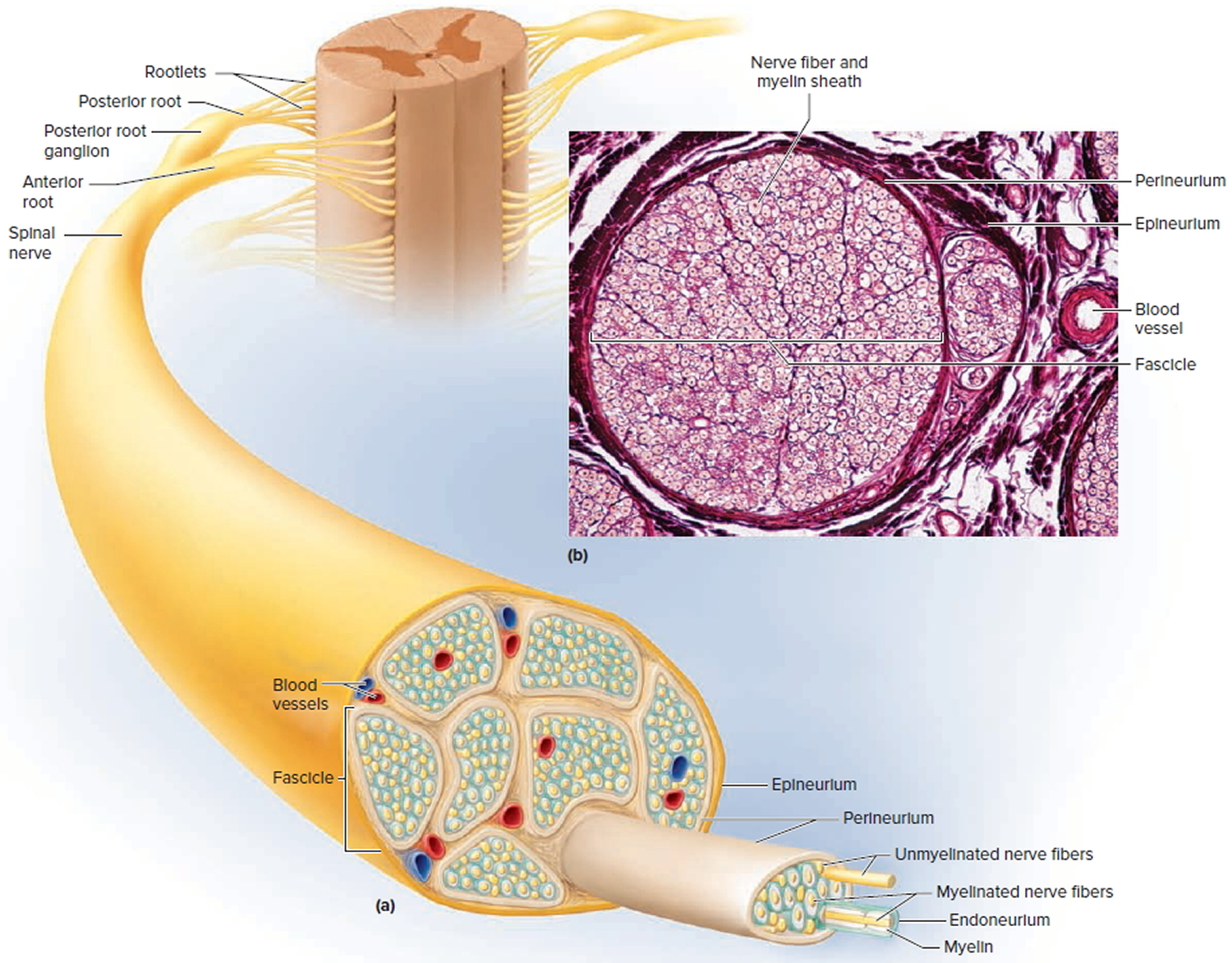
A nerve is a cordlike organ composed of numerous nerve fibers (axons) bound together by connective tissue. If you compare a nerve fiber to a wire carrying an electrical current in one direction, a nerve would be comparable to an electrical cable composed of thousands of wires carrying currents in opposite directions. A nerve contains anywhere from a few nerve fibers to (in the optic nerve) a million. Nerves usually have a pearly white color and resemble frayed string as they divide into smaller and smaller branches. As we move away from the spinal nerves proper, the smaller branches are called peripheral nerves, and their disorders are collectively called peripheral neuropathy.
Figure 1. Spinal cord
Spinal cord parts
The spinal cord consists of thirty-one segments, each of which gives rise to a pair of spinal nerves. Although the spinal cord is not visibly segmented, the part supplied by each pair of nerves is called a segment. The cord exhibits longitudinal grooves on its anterior and posterior sides—the anterior median fissure and posterior median sulcus, respectively. These nerves (part of the peripheral nervous system) branch to various body parts and connect them with the central nervous system.
The spinal cord is divided into cervical, thoracic, lumbar, and sacral regions. It may seem odd that it has a sacral region when the cord itself ends well above the sacrum. These regions, however, are named for the level of the vertebral column from which the spinal nerves emerge, not for the vertebrae that contain the cord itself.
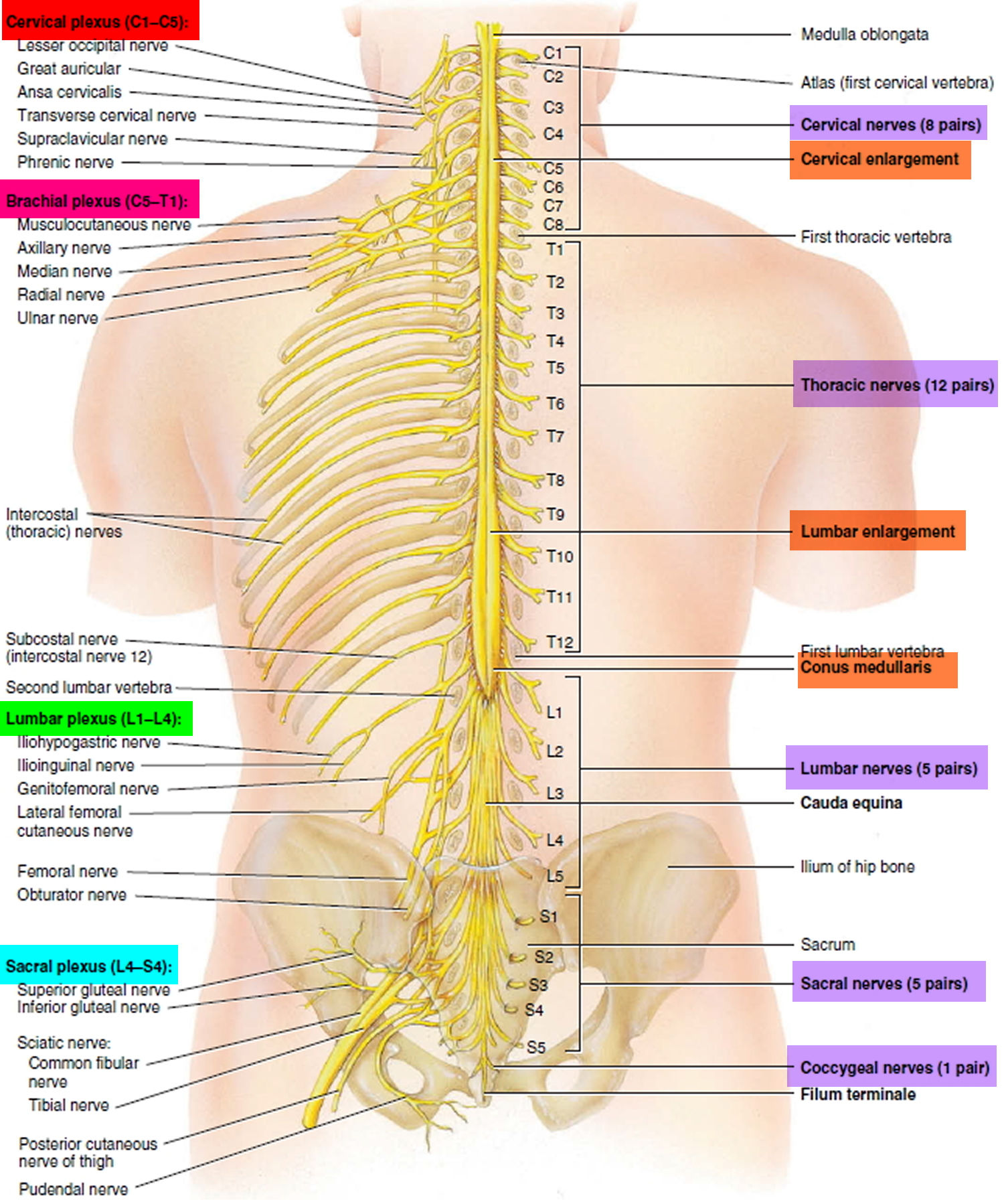
In two areas, the spinal cord is a little thicker than elsewhere. In the inferior cervical region, a cervical enlargement gives rise to nerves of the upper limbs. In the lumbosacral region, there is a similar lumbar enlargement that issues nerves to the pelvic region and lower limbs. Inferior to the lumbar enlargement, the cord tapers to a point called the medullary cone. Arising from the lumbar enlargement and medullary cone is a bundle of nerve roots that occupy the vertebral canal from L2 (lumbar vertbra L2) to S5 (sacral vertbra S5). This bundle, named the cauda equina for its resemblance to a horse’s tail, innervates the pelvic organs and lower limbs.
Figure 2. Spinal cord segments
Spinal Nerves
There are 31 pairs of spinal nerves: 8 cervical (C1–C8), 12 thoracic (T1–T12), 5 lumbar (L1–L5), 5 sacral (S1–S5), and 1 coccygeal (Co1). The first cervical nerve emerges between the skull and atlas, and the others emerge through intervertebral foramina, including the anterior and posterior foramina of the sacrum and the sacral hiatus. Thus, spinal nerves C1 through C7 emerge superior to the correspondingly numbered vertebrae (nerve C5 above vertebra C5, for example); nerve C8 emerges inferior to vertebra C7; and below this, all the remaining nerves emerge inferior to the correspondingly numbered vertebrae (nerve L3 inferior to vertebra L3, for example).
Proximal Branches
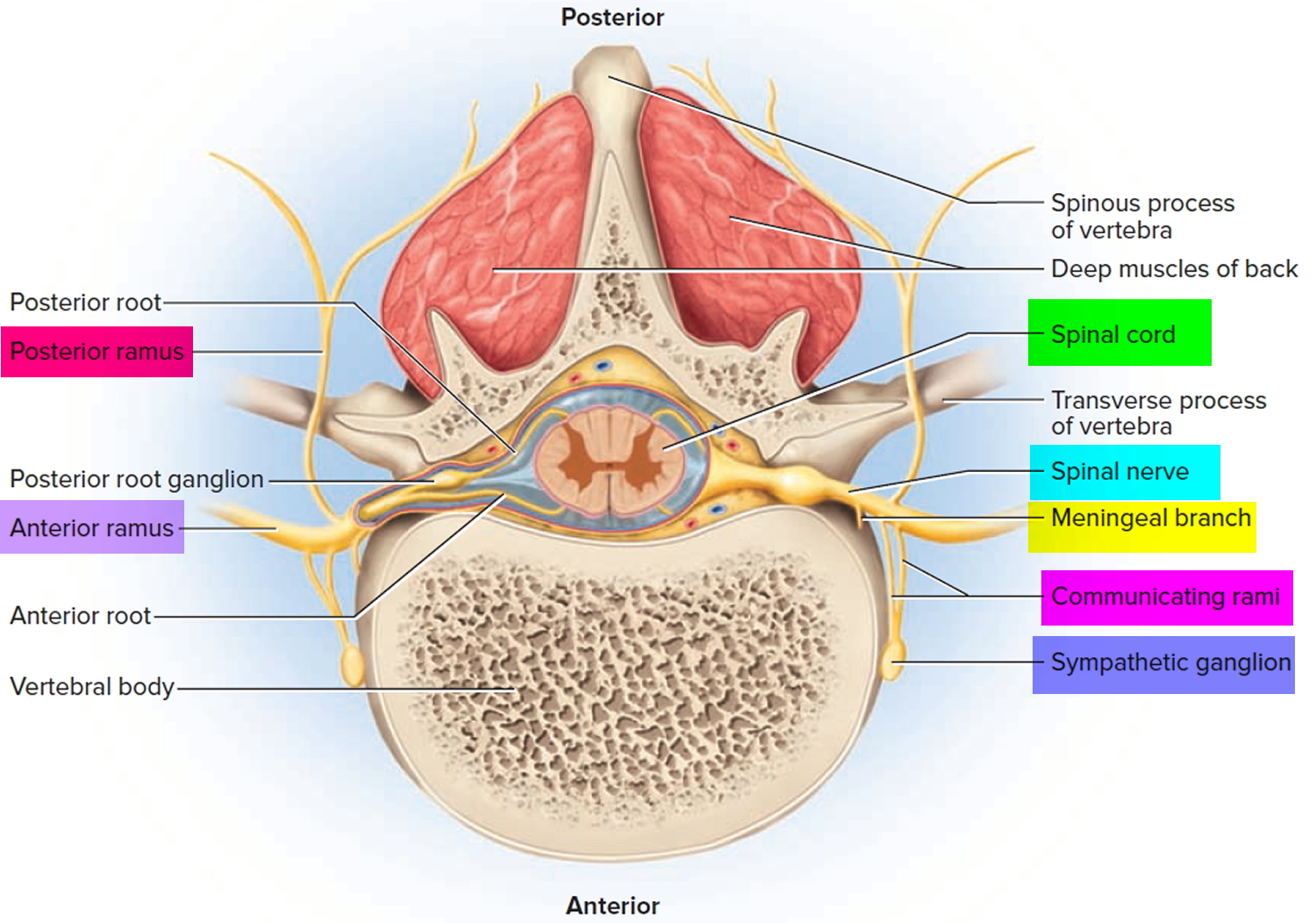
Each spinal nerve arises from two points of attachment to the spinal cord. In each segment of the cord, six to eight nerve rootlets emerge from the anterior surface and converge to form the anterior (ventral) root of the spinal nerve. Another six to eight rootlets emerge from the posterior surface and converge to form the posterior (dorsal) root. A short distance away from the spinal cord, the posterior root swells into a posterior (dorsal) root ganglion, which contains the somas (neuron bodies) of sensory neurons. There is no corresponding ganglion on the anterior root.
Slightly distal to the ganglion, the anterior and posterior roots merge, leave the dural sheath, and form the spinal nerve proper. The nerve then exits the vertebral canal through the intervertebral foramen. The spinal nerve is a mixed nerve, carrying sensory signals to the spinal cord by way of the posterior root and ganglion, and motor signals out to more distant parts of the body by way of the anterior root.
The anterior and posterior roots are shortest in the cervical region and become longer inferiorly. The roots that arise from segments L2 to Co1 of the cord form the cauda equina. Some viruses can invade the CNS by way of the spinal nerve roots (e.g varicella-zoster virus of shingles and herpes simplex virus of core sores or genital herpes).
Figure 3. Spinal nerve
Distal Branches
Distal to the vertebrae, the branches of a spinal nerve are more complex. Immediately after emerging from the intervertebral foramen, the nerve divides into an anterior ramus, posterior ramus, and a small meningeal branch. Thus, each spinal nerve branches on both ends—into anterior and posterior roots approaching the spinal cord, and anterior and posterior rami leading away from the vertebral column.
The meningeal branch reenters the vertebral canal and innervates the meninges, vertebrae, and spinal ligaments with sensory and motor fibers. The posterior ramus innervates the muscles and joints in that region of the spine and the skin of the back. The larger anterior ramus innervates the anterior and lateral skin and muscles of the trunk, and gives rise to nerves of the limbs.
The anterior ramus differs from one region of the trunk to another. In the thoracic region, it forms an intercostal nerve, which travels along the inferior margin of a rib and innervates the skin and intercostal muscles (thus contributing to breathing). Sensory fibers of the intercostal nerve branches to the skin are the most common routes of viral migration in the painful disease known as shingles. Motor fibers of the intercostal nerves innervate the internal oblique, external oblique, and transverse abdominal muscles. All other anterior rami form the nerve plexuses.
The anterior ramus also gives off a pair of communicating rami, which connect with a string of sympathetic chain ganglia alongside the vertebral column. These are seen only in spinal nerves T1 through L2. They are components of the sympathetic nervous system.
Figure 4. Rami of the spinal nerve
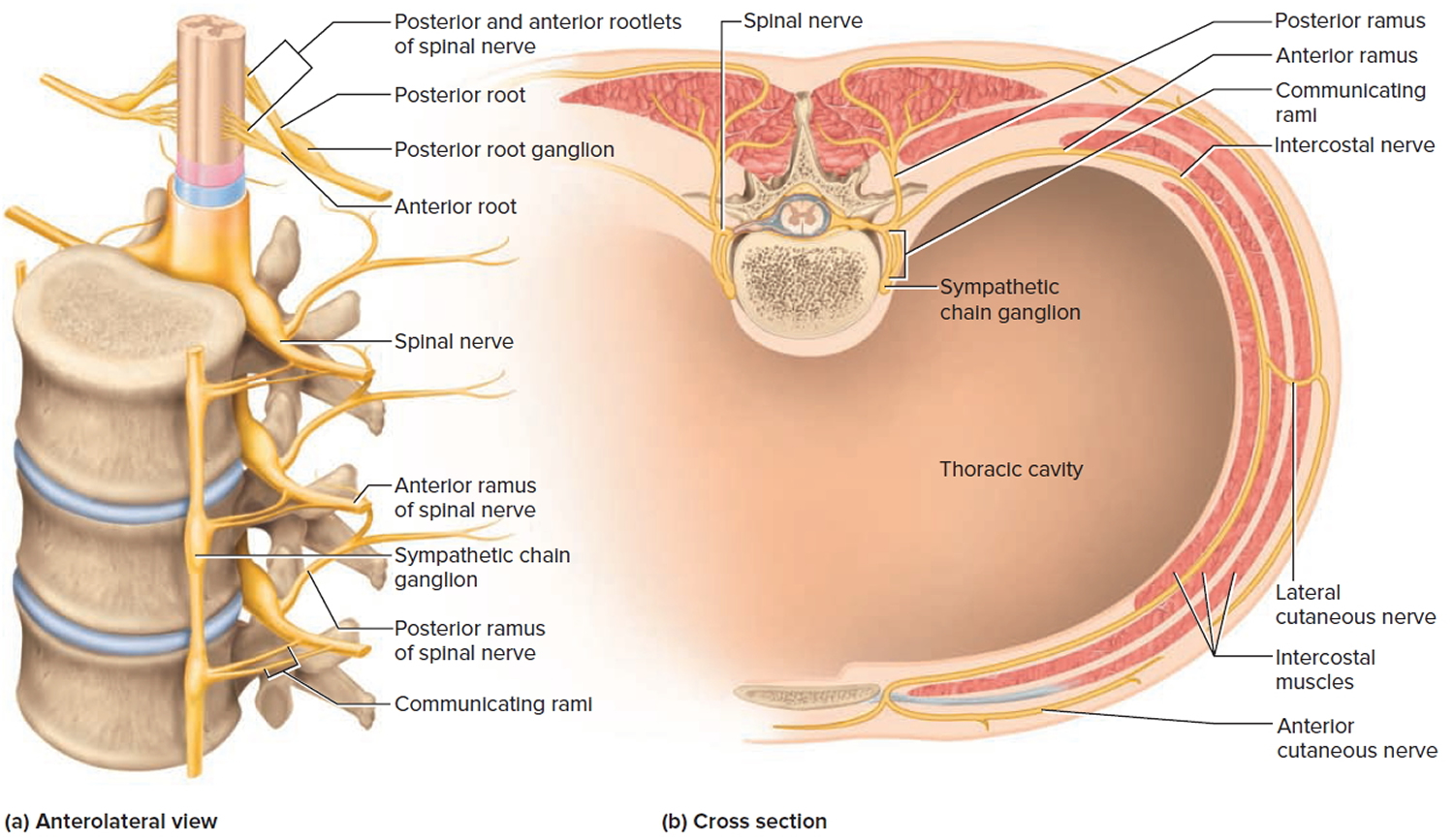
Figure 5. Spinal nerve fiber anatomy
If a nerve resembles a thread, a ganglion resembles a knot in the thread. A ganglion is a cluster of neurosomas outside the central nervous system. It is enveloped in an epineurium continuous with that of the nerve. Among the neurosomas are bundles of nerve fibers leading into and out of the ganglion. Figure 9 shows a type of ganglion associated with the spinal nerves.
Figure 6. Spinal nerve ganglion
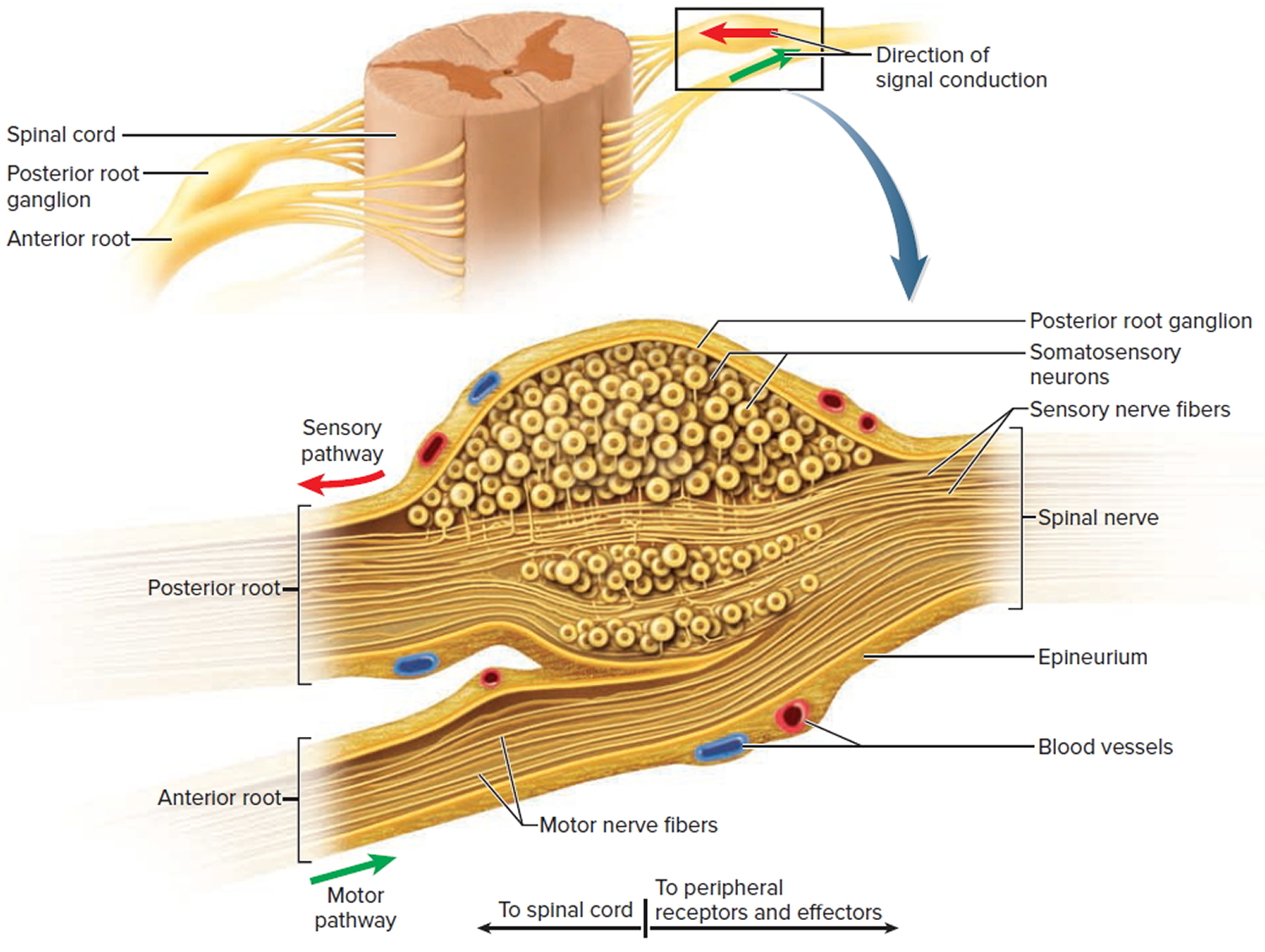 Note: The posterior root ganglion contains the somas of unipolar sensory neurons conducting signals from peripheral sense organs toward the spinal cord. Below this is the anterior root of the spinal nerve, which conducts motor signals away from the spinal cord, toward peripheral effectors. Note that the anterior root is not part of the ganglion.
Note: The posterior root ganglion contains the somas of unipolar sensory neurons conducting signals from peripheral sense organs toward the spinal cord. Below this is the anterior root of the spinal nerve, which conducts motor signals away from the spinal cord, toward peripheral effectors. Note that the anterior root is not part of the ganglion.Nerve Plexuses
Except in the thoracic region, the anterior rami branch and merge repeatedly to form five webs called nerve plexuses: the small cervical plexus in the neck, the brachial plexus near the shoulder, the lumbar plexus of the lower back, the sacral plexus immediately inferior to this, and finally, the tiny coccygeal plexus adjacent to the lower sacrum and coccyx.
Two of the nerves arising from these plexuses, the radial and sciatic, are sites of unique nerve injuries. Some nerves tabulated have somatosensory and motor
functions. Somatosensory means that they carry sensory signals from bones, joints, muscles, and the skin, in contrast to sensory input from the viscera or from special sense organs such as the eyes and ears. Somatosensory signals are for touch, heat, cold, stretch, pressure, pain, and other sensations. One of the most important sensory roles of these nerves is proprioception, in which the brain receives information about body position and movements from nerve endings in the muscles, tendons, and joints. The brain uses this information to adjust muscle actions and thereby maintain equilibrium (balance) and coordination.
The motor function of these nerves is primarily to stimulate the contraction of skeletal muscles. They also innervate the bones of the corresponding regions, and carry autonomic fibers to some viscera and blood vessels, thus adjusting blood flow to local needs. You may assume that for each muscle, these nerves also carry sensory fibers from its proprioceptors.
Cutaneous Innervation and Dermatomes
Each spinal nerve except C1 receives sensory input from a specific area of skin called a dermatome. A dermatome map is a diagram of the cutaneous regions innervated by each spinal nerve. Such a map is oversimplified, however, because the dermatomes overlap at their edges by as much as 50%. Therefore, severance of one sensory nerve root does not entirely deaden sensation from a dermatome. It is necessary to sever or anesthetize three sequential spinal nerves to produce a total loss of sensation from one dermatome. Spinal nerve damage is assessed by testing the dermatomes with pinpricks and noting areas in which the patient has no sensation.
Figure 7. Dermatome (spinal nerves sensory innvervation)
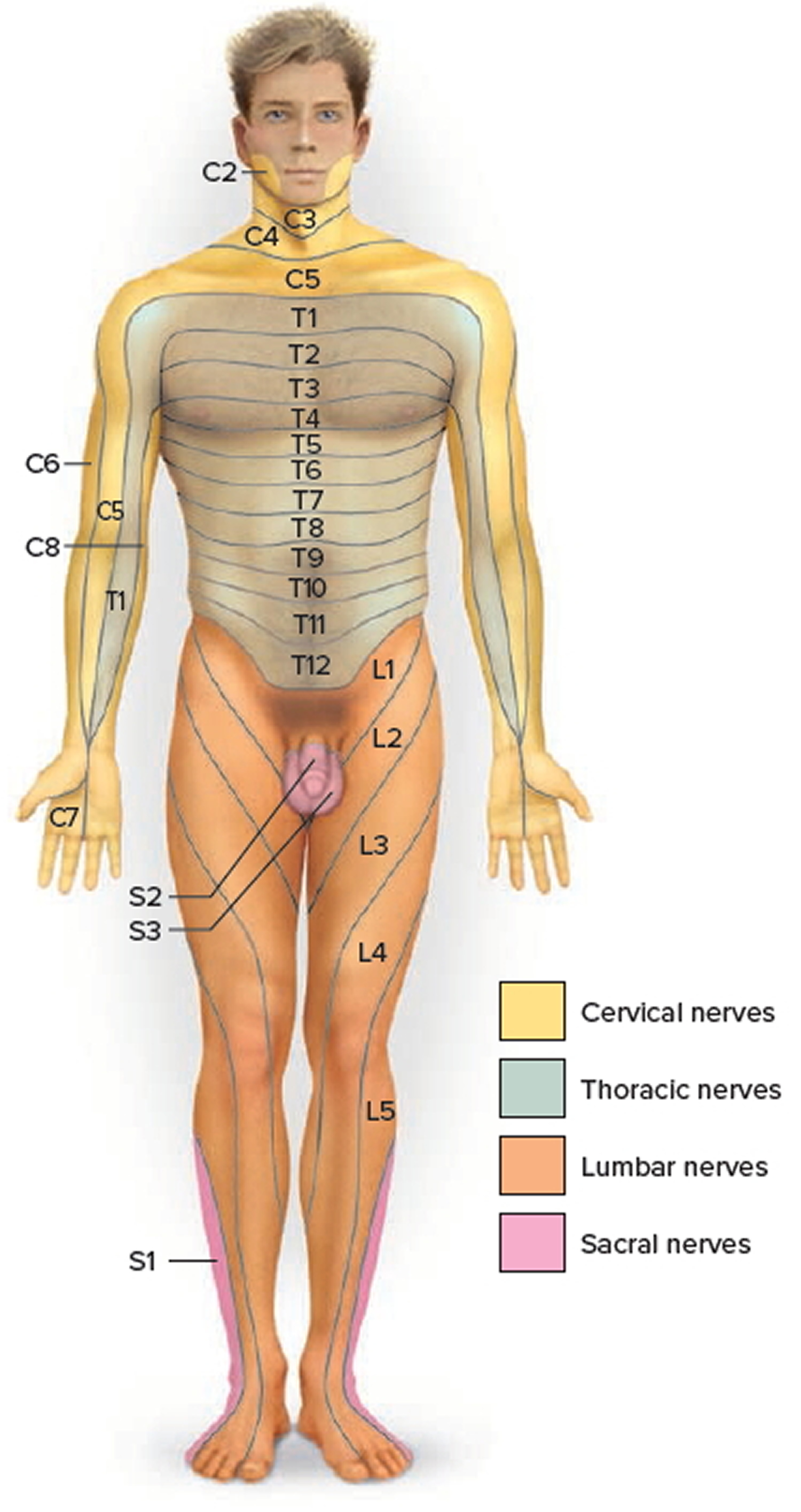
Note: Each zone of the skin is innervated by sensory branches of the spinal nerves indicated by the labels. Nerve C1 does not innervate the skin.
What does the spinal cord do ?
The spinal cord serves four principal functions:
- Conduction. It contains bundles of nerve fibers that conduct information up and down the cord, connecting different levels of the trunk with each other and with the brain. This enables sensory information to reach the brain, motor commands to reach the effectors, and input received at one level of the cord to affect output from another level.
- Neural integration. Pools of spinal neurons receive input from multiple sources, integrate the information, and execute an appropriate output. For example, the spinal cord can integrate the stretch sensation from a full bladder with cerebral input concerning the appropriate time and place to urinate and execute control of the bladder accordingly.
- Locomotion. Walking involves repetitive, coordinated contractions of several muscle groups in the limbs. Motor neurons in the brain initiate walking and determine its speed, distance, and direction, but the simple repetitive muscle contractions that put one foot in front of another, over and over, are coordinated by groups of neurons called central pattern generators in the cord. These neural circuits produce the sequence of outputs to the extensor and flexor muscles that cause alternating movements of the lower limbs.
- Reflexes. Spinal reflexes play vital roles in posture, motor coordination, and protective responses to pain or injury.
Figure 8. Spinal cord anatomy
Protective Structures of the Spinal cord
The nervous tissue of the central nervous system (CNS) is very delicate and does not respond well to injury or damage. Accordingly, nervous tissue requires considerable protection. The first layer of protection for the central nervous system is the hard bony skull and vertebral column. The skull encases the brain and the vertebral column surrounds the spinal cord, providing strong protective defenses against damaging blows or bumps. The spinal cord is located within the vertebral canal of the vertebral column. The surrounding vertebrae provide a sturdy shelter for the enclosed spinal cord. The vertebral ligaments, meninges, and cerebrospinal fluid provide additional protection. The second protective layer is the meninges, three membranes that lie between the bony encasement and the nervous tissue in both the brain and spinal cord. Finally, a space between two of the meningeal membranes contains cerebrospinal fluid, a buoyant liquid that suspends the central nervous tissue in a weightless environment while surrounding it with a shock-absorbing, hydraulic cushion.
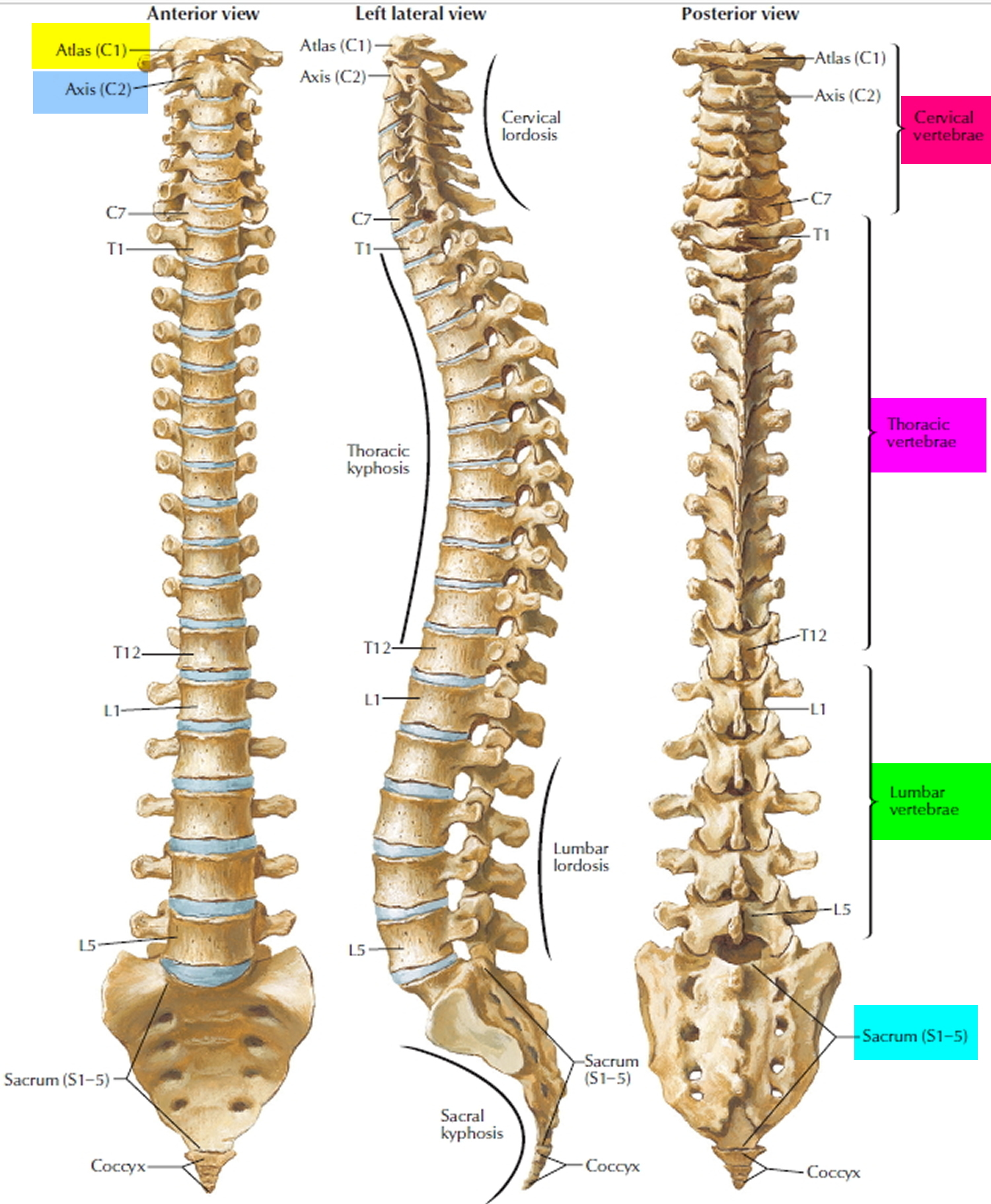
The spinal column is not all bone. Between the vertebrae are discs of semi-rigid cartilage and narrow spaces called foramen that act as passages through which the spinal nerves travel to and from the rest of the body. These are places where the spinal cord is particularly vulnerable to direct injury.
Figure 9. Vertebral column
Meninges of the Spinal Cord
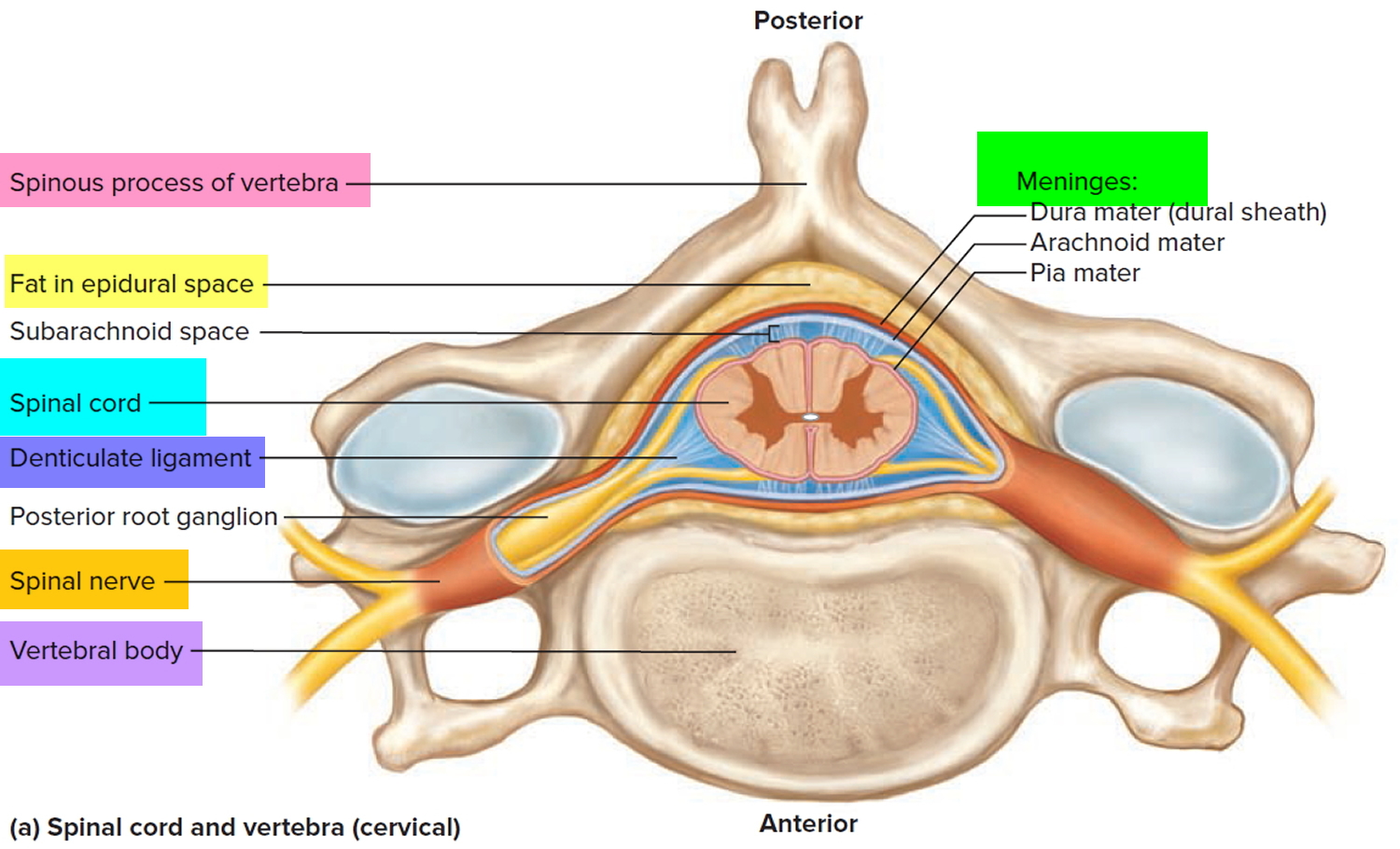
The spinal cord and brain are enclosed in three fibrous membranes called meninges (singular meninx). These membranes separate the soft tissue of the central nervous system from the bones of the vertebrae and skull. From superficial to deep, they are the dura mater, arachnoid mater, and pia mater. The dura mater forms a loose-fitting sleeve called the dural sheath around the spinal cord. It is a tough membrane about as thick as a rubber kitchen glove, composed of multiple layers of dense irregular connective tissue. The space between the sheath and vertebral bones, called the epidural space, is occupied by blood vessels, adipose tissue, and loose connective tissue. Anesthetics are sometimes introduced to this space to block pain signals during childbirth or surgery; this procedure is called epidural anesthesia.
The arachnoid mater consists of the arachnoid membrane—five or six layers of squamous to cuboidal cells adhering to the inside of the dura—and a looser array of cells and collagenous and elastic fibers spanning the gap between the arachnoid membrane and the pia mater. This gap, the subarachnoid space, is filled with cerebrospinal fluid. Inferior to the medullary cone, the subarachnoid space is called the lumbar cistern and is occupied by the cauda equina and cerebrospinal fluid.
The pia mater is a delicate, transparent membrane composed of one or two layers of squamous to cuboidal cells and delicate collagenous and elastic fibers. It closely follows the contours of the spinal cord. It continues beyond the medullary cone as a fibrous strand, the terminal filum, within the lumbar cistern. At the level of vertebra S2, it exits the lower end of the cistern and fuses with the dura mater, and the two form a coccygeal ligament that anchors the cord and meninges to vertebra Co1. At regular intervals along the cord, extensions of the pia called denticulate ligaments extend through the arachnoid to the dura, anchoring the cord and limiting side-to-side movements. When a sample of cerebrospinal fluid is needed for clinical purposes, it is taken from the lumbar cistern by a procedure called lumbar puncture (or colloquially, spinal tap). A spinal needle is inserted between two vertebrae at level L3/L4 or L4/L5, where there is no risk of accidental injury to the spinal cord (which ends at L1 to L2).
Cross-Sectional Anatomy of Spinal Cord
The spinal cord, like the brain, consists of two kinds of nervous tissue called gray and white matter. Gray matter has a relatively dull color because it contains little myelin. It contains the somas, dendrites, and proximal parts of the axons of neurons. It is the site of synaptic contact between neurons, and therefore the site of all neural integration in the spinal cord. White matter, by contrast, is whitish containing a mixture of proteins and fat-like substances called myelin covering the axons and allows electrical signals to flow quickly and freely. Myelin is much like the insulation around electrical wires. The white matter is composed of bundles of axons, called tracts, that carry signals from one level of the central nervous system (CNS) to another. It is formed by axon-insulating cells called oligodendrocytes. Because of its whitish color, the outer section of the spinal cord—which is formed by bundles of myelinated axons—is called white matter. Both gray and white matter also have an abundance of glial cells.
Figure 10. Spinal cord cross section
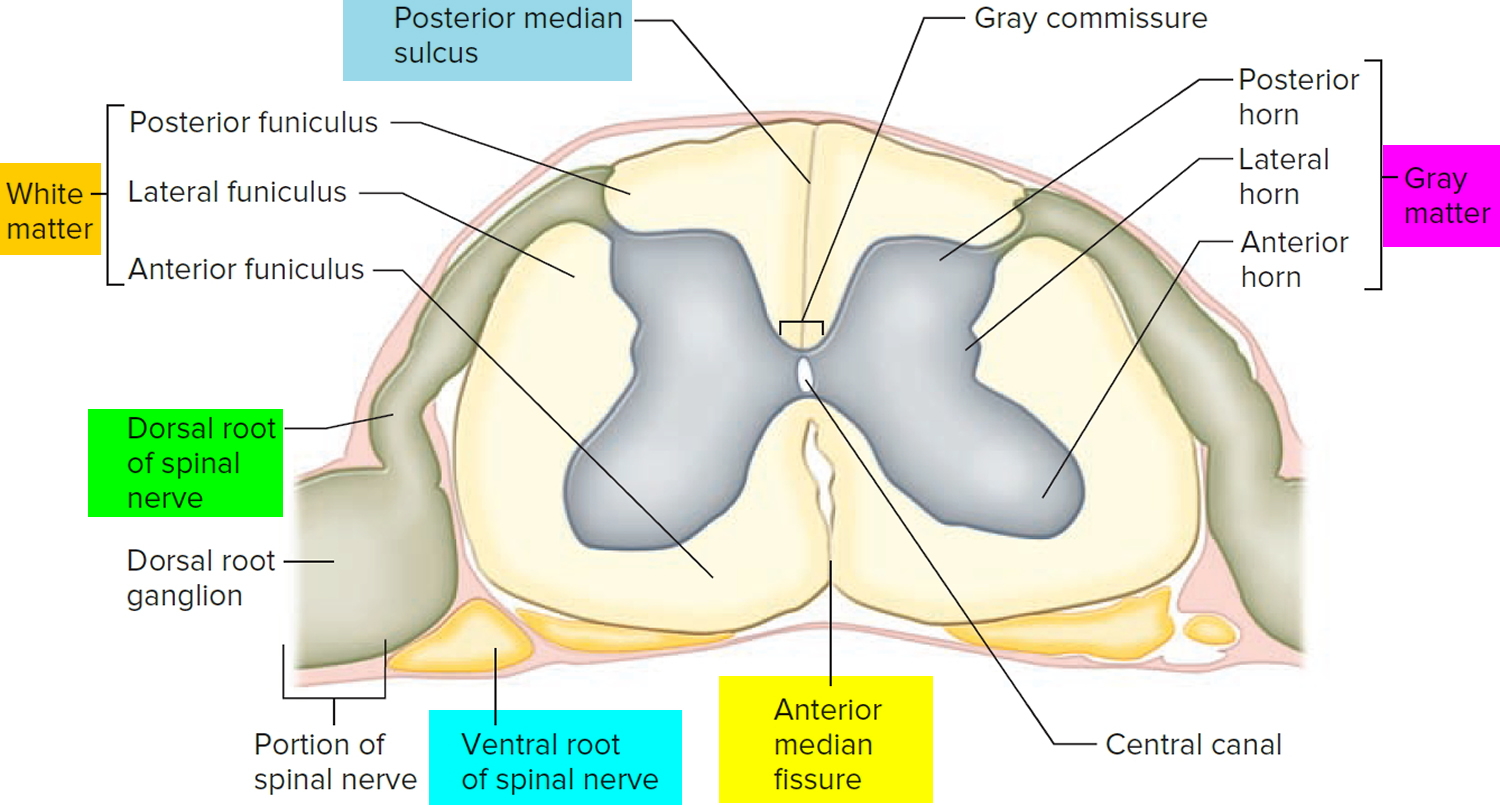
Gray Matter
The spinal cord has a central core of gray matter that looks somewhat butterfly- or H-shaped in cross sections. The core consists mainly of two posterior (dorsal) horns, which extend toward the posterolateral surfaces of the cord, and two thicker anterior (ventral) horns, which extend toward the anterolateral surfaces.
The right and left sides of the gray matter are connected by a median bridge called the gray commissure. In the middle of the commissure is the central canal, which is collapsed in most areas of the adult spinal cord, but in some places (and in young children) remains open, lined with ependymal cells, and filled with cerebrospinal fluid.
The posterior horn receives sensory nerve fibers from the spinal nerves, which usually synapse with networks of interneurons in the horn. The anterior horn contains the large neurosomas of motor neurons whose axons lead out to the skeletal muscles. The interneurons and motor neurons are especially abundant in the cervical and lumbar enlargements. The high density of neurons in these regions is related to motor control and sensation in the upper and lower limbs.
An additional lateral horn is visible on each side of the gray matter from segments T2 through L1 of the cord. It contains neurons of the sympathetic nervous system, which send their axons out of the cord by way of the anterior root along with the somatic efferent fibers.
White Matter
The white matter of the spinal cord surrounds the gray matter. It consists of bundles of axons that course up and down the cord and provide avenues of communication between different levels of the CNS. These bundles are arranged in three pairs called columns or funiculi—a posterior (dorsal), lateral, and anterior (ventral) column on each side. Each column consists of subdivisions called tracts or fasciculi.
Spinal Cord Tracts
Knowledge of the locations and functions of the spinal tracts is essential in diagnosing and managing spinal cord injuries.
Ascending tracts carry sensory information up the cord, and descending tracts conduct motor impulses down. All nerve fibers in a given tract have a similar origin, destination, and function. Many of these fibers have their origin or destination in a region called the brainstem. Described more fully in the human brain article.
Several of these tracts undergo decussation as they pass up or down the brainstem and spinal cord—meaning that they cross over from the left side of the body to the right, or vice versa. As a result, the left side of the brain receives sensory information from the right side of the body and sends motor commands to that side, while the right side of the brain senses and controls the left side of the body. Therefore, a stroke that damages motor centers of the right side of the brain can cause paralysis of the left limbs and vice versa.
When the origin and destination of a tract are on opposite sides of the body, anatomists say they are contralateral to each other. When a tract does not decussate, its origin and destination are on the same side of the body and anatomists say they are ipsilateral. Bear in mind that each tract is repeated on the right and left sides of the spinal cord.
Figure 11. Spinal cord tracts
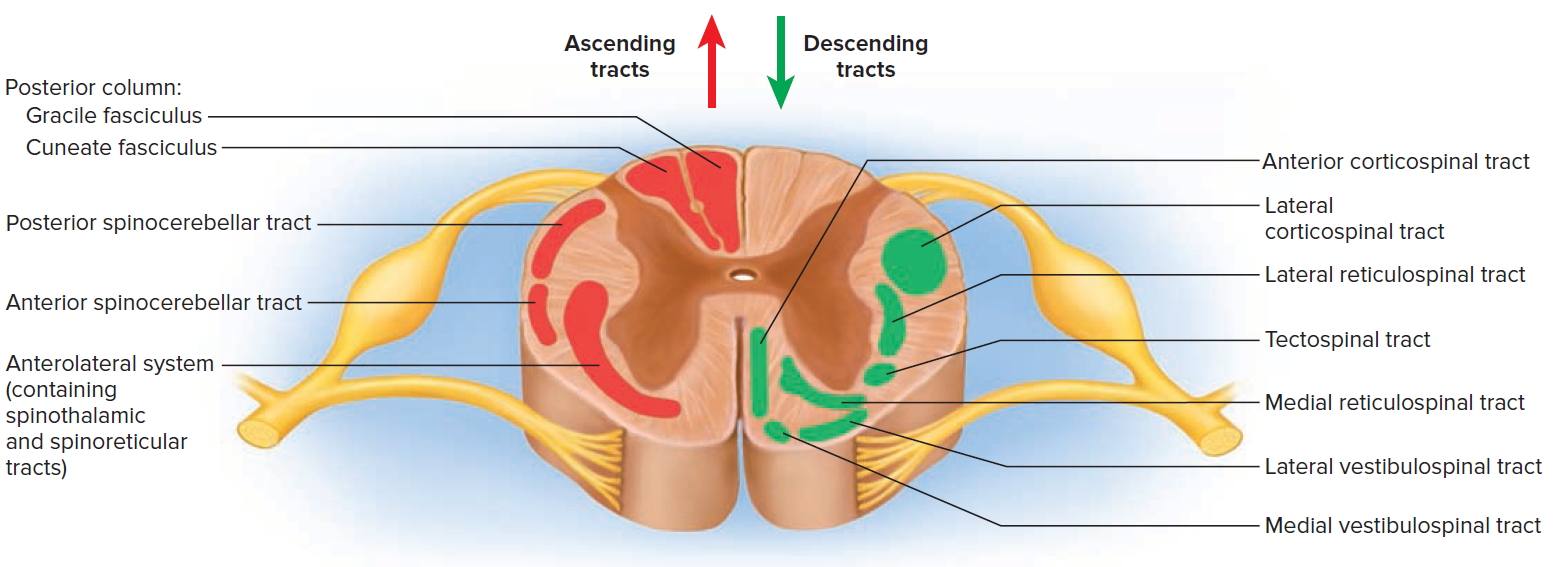
Figure 12. Processing of sensory input and motor output by the spinal cord
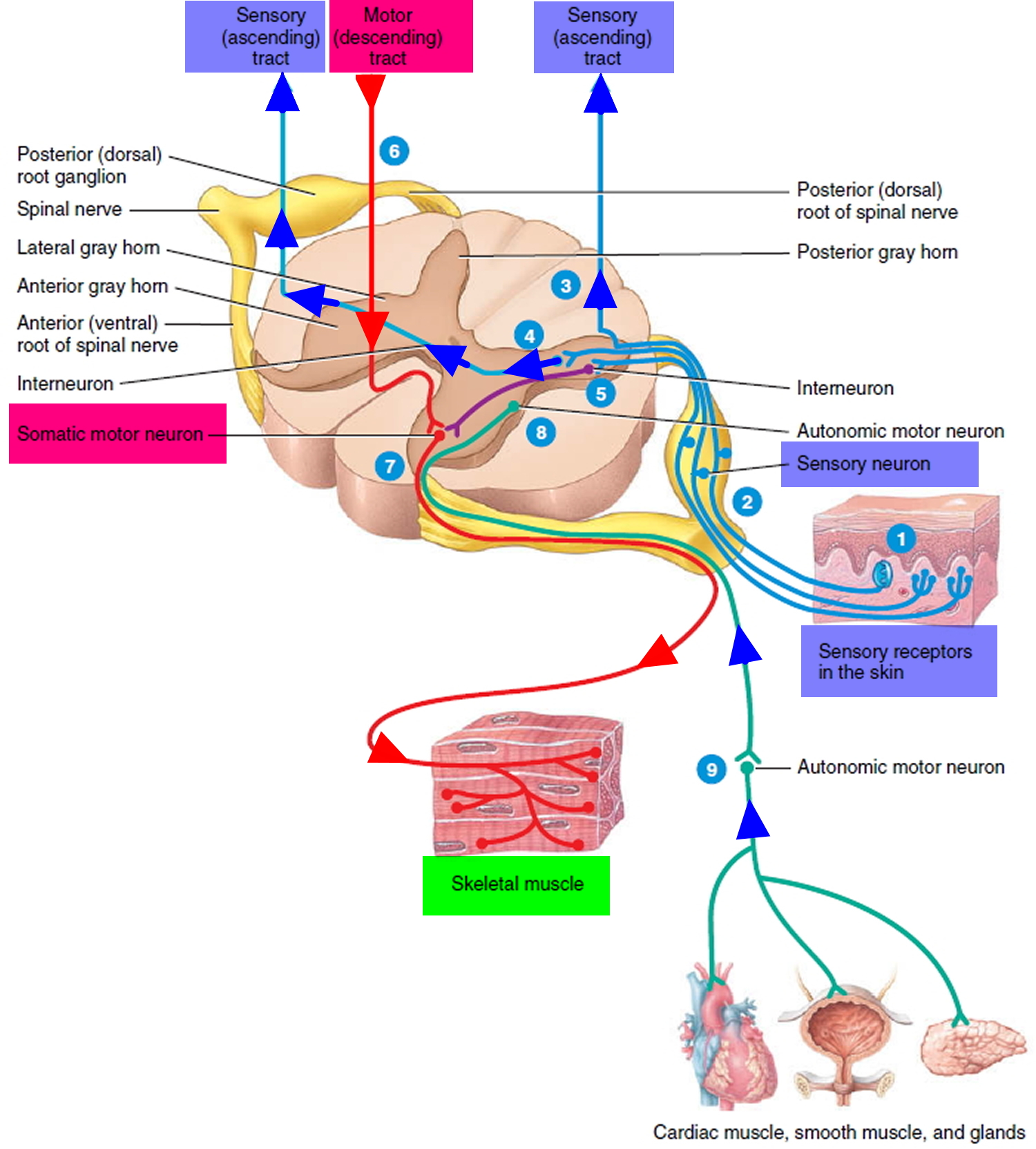
Note: Sensory input is conveyed from sensory receptors to the posterior gray horns of the spinal cord, and motor output is conveyed from the anterior and lateral gray horns of the spinal cord to effectors (muscles and glands).
Ascending Tracts
Ascending tracts carry sensory signals up the spinal cord. Sensory signals typically travel across three neurons from their origin in the receptors to their destination in the brain: a first-order neuron that detects a stimulus and transmits a signal to the spinal cord or brainstem; a second-order neuron that continues as far as a “gateway” called the thalamus at the upper end of the brainstem; and a third-order neuron that carries the signal the rest of the way to the cerebral cortex. The axons of these neurons are called the first- through third-order nerve fibers.
Figure 13. Spinal cord ascending tracts to the brain
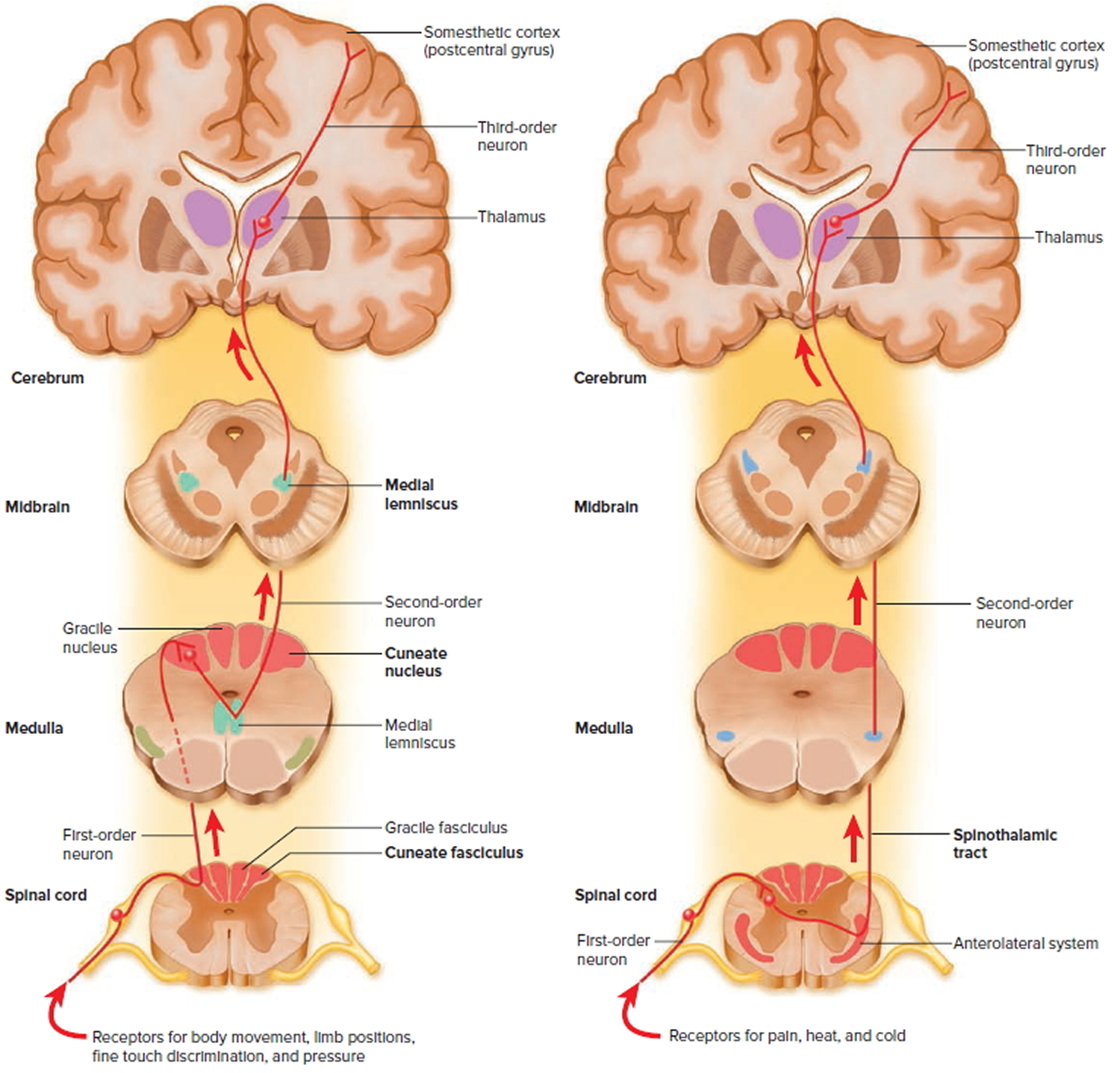
The major ascending tracts are as follows. The names of most of them consist of the prefix spino- followed by a root denoting the destination of its fibers in the brain, although this naming system does not apply to the first two.
Gracile fasciculus
The gracile fasciculus carries signals from the midthoracic and lower parts of the body. Below vertebra T6, it composes the entire posterior column. At T6, it is joined by the cuneate fasciculus, discussed next. It consists of first-order nerve fibers that travel up the ipsilateral side of the spinal cord and terminate at the gracile nucleus in the medulla oblongata of the brainstem. These fibers carry signals for vibration, visceral pain, deep and discriminative touch (touch whose location one can precisely identify), and especially proprioception from the lower limbs and lower trunk. Proprioception is the nonvisual sense of the position and movements of the body.
Cuneate fasciculus
The cuneate fasciculus joins the gracile fasciculus at the T6 level. It occupies the lateral portion of the posterior column and forces the gracile fasciculus medially. It carries the same type of sensory signals, originating from T6 and up (from the upper limbs and chest). Its fibers end in the cuneate nucleus on the ipsilateral side of the medulla oblongata. In the medulla, second-order fibers of the gracile and cuneate systems decussate and form the medial lemniscus, a tract of nerve fibers that leads the rest of the way up the brainstem to the thalamus. Third-order fibers go from the thalamus to the cerebral cortex. Because of decussation, the signals carried by the gracile and cuneate fasciculi ultimately go to the contralateral cerebral hemisphere.
Spinothalamic tract
The spinothalamic tract and some smaller tracts form the anterolateral system, which passes up the anterior and lateral columns of the spinal cord. The spinothalamic tract carries signals for pain, temperature, pressure, tickle, itch, and light or crude touch. Light touch is the sensation produced by stroking hairless skin with a feather or cotton wisp, without indenting the skin; crude touch is touch whose location one can only vaguely identify.
In this pathway, first-order neurons end in the posterior horn of the spinal cord near the point of entry. Here they synapse with second-order neurons, which decussate and form the contralateral ascending spinothalamic tract. These fibers lead all the way to the thalamus. Third-order neurons continue from there to the cerebral cortex. Because of decussation, sensory signals in this tract arrive in the cerebral hemisphere contralateral to their point of origin.
Spinoreticular tract
The spinoreticular tract also travels up the anterolateral system. It carries pain signals resulting from tissue injury. The first-order sensory neurons enter the posterior horn and immediately synapse with second-order neurons. These decussate to the opposite anterolateral system, ascend the cord, and end in a loosely organized core of gray matter called the reticular formation in the medulla and pons. Third-order neurons continue from the pons to the thalamus, and fourth-order neurons complete the path from there to the cerebral cortex.
Posterior and anterior spinocerebellar tracts
The posterior and anterior spinocerebellar tracts travel through the lateral column and carry proprioceptive signals from the limbs and trunk to the cerebellum at the rear of the brain. Their first-order neurons originate in muscles and tendons and end in the posterior horn of the spinal cord. Second-order neurons send their fibers up the spinocerebellar tracts and end in the cerebellum.
Fibers of the posterior tract travel up the ipsilateral side of the spinal cord. Those of the anterior tract cross over and travel up the contralateral side but then cross back in the brainstem to enter the ipsilateral side of the cerebellum. Both tracts provide the cerebellum with feedback needed to coordinate muscle action.
Descending Tracts
Descending tracts carry motor signals down the brainstem and spinal cord. A descending motor pathway typically involves two neurons called the upper and lower motor neurons. The upper motor neuron begins with a soma in the cerebral cortex or brainstem and has an axon that terminates on a lower motor
neuron in the brainstem or spinal cord. The axon of the lower motor neuron then leads the rest of the way to the muscle or other target organ. The names of most descending tracts consist of a word root denoting the point of origin in the brain, followed by the suffix -spinal.
Figure 14. Spinal cord descending tracts from the brain
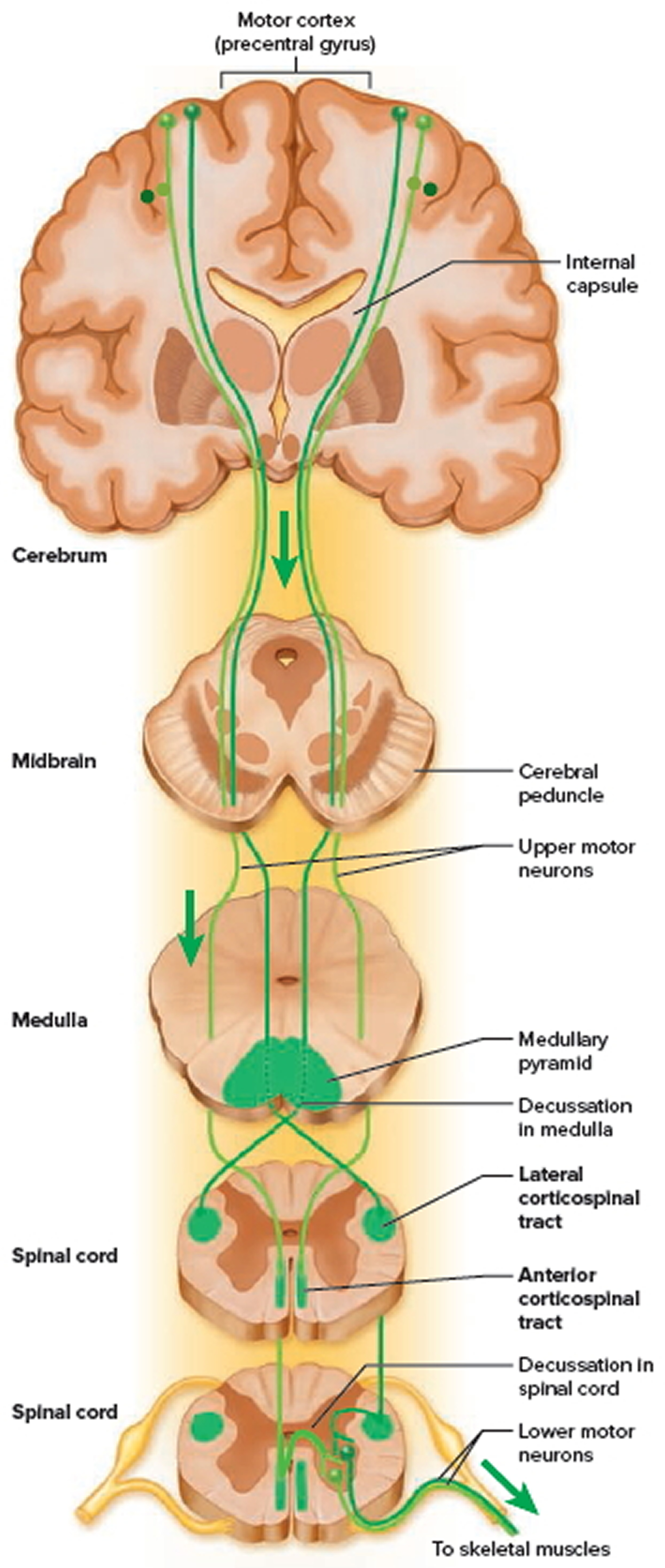
Corticospinal tracts
The corticospinal tracts carry motor signals from the cerebral cortex for precise, finely coordinated limb movements. The fibers of this system form ridges called pyramids on the anterior surface of the medulla oblongata, so these tracts were once called pyramidal tracts. Most corticospinal fibers decussate in the lower medulla and form the lateral corticospinal tract on the contralateral side of the spinal cord. A few fibers remain uncrossed and form the anterior corticospinal tract on the ipsilateral side. Fibers of the anterior tract decussate lower in the cord, however, so even they control contralateral muscles. This tract gets smaller as it descends and gives off nerve fibers, and usually disappears by the midthoracic level.
Tectospinal tract
The tectospinal tract begins in a midbrain region called the tectum and crosses to the contralateral side of the midbrain. It descends through the brainstem to the upper spinal cord on that side, going only as far as the neck. It is involved in reflex turning of the head, especially in response to sights and sounds.
Lateral and medial reticulospinal tracts
The lateral and medial reticulospinal tracts originate in the reticular formation of the brainstem. They control muscles of the upper and lower limbs, especially to maintain posture and balance. They also contain descending analgesic pathways that reduce the transmission of pain signals to the brain.
Lateral and medial vestibulospinal tracts
The lateral and medial vestibulospinal tracts begin in the brainstem vestibular nuclei, which receive signals for balance from the inner ear. The lateral vestibulospinal tract passes down the anterior column of the spinal cord and facilitates neurons that control extensor muscles of the limbs, thus inducing the limbs to stiffen and straighten. This is an important reflex in responding to body tilt and keeping one’s balance. The medial vestibulospinal tract splits into ipsilateral and contralateral fibers that descend through the anterior column on both sides of the cord and terminate in the neck. It plays a role in the control of head position.
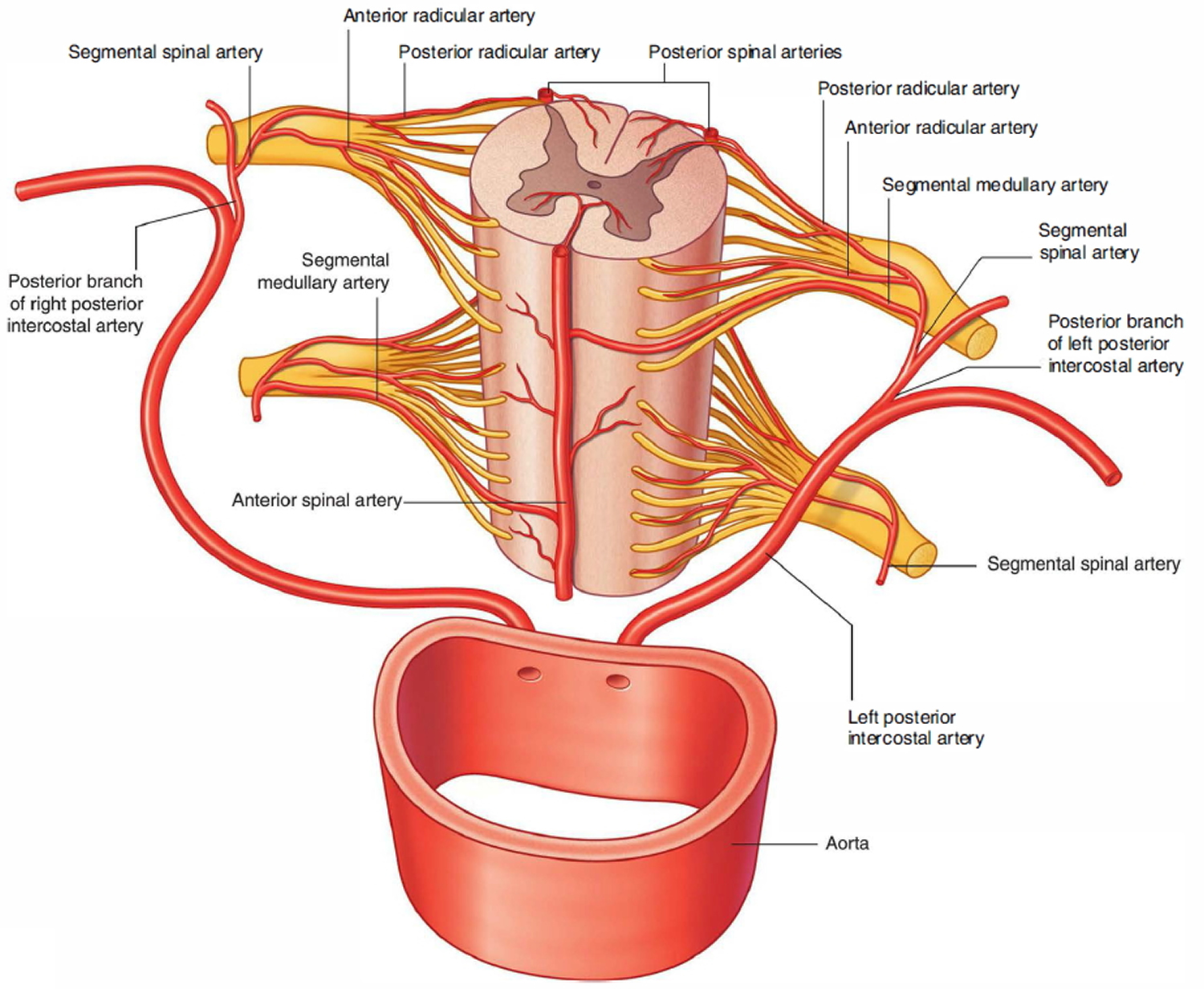
Spinal cord blood supply
The arterial supply to the spinal cord comes from two sources. It consists of:
- The longitudinal vessels.
- Segmental spinal arteries.
The longitudinal vessels consist of:
- A single anterior spinal artery, which originates within the cranial cavity as the union of two vessels that arise from the vertebral arteries-the resulting single anterior spinal artery passes inferiorly, approximately parallel to the anterior median fissure, along the surface of the spinal cord; and
- Two posterior spinal arteries, which also originate in the cranial cavity, usually arising directly from a terminal branch of each vertebral artery (the posterior inferior cerebellar artery)-the right and left posterior spinal arteries descend along the spinal cord, each as two branches that bracket the posterolateral sulcus and the connection of posterior roots with the spinal cord.
The anterior and posterior spinal arteries are reinforced along their length by eight to ten segmental medullary arteries. The largest of these is the arteria radicularis magna or the artery of Adamkiewicz. This vessel arises in the lower thoracic or upper lumbar region, usually on the left side, and reinforces the arterial supply to the lower portion of the spinal cord, including the lumbar enlargement.
Segmental spinal arteries
Segmental spinal arteries arise predominantly from the vertebral and deep cervical arteries in the neck, the posterior intercostal arteries in the thorax, and the lumbar arteries in the abdomen.
After entering an intervertebral foramen, the segmental spinal arteries give rise to anterior and posterior radicular arteries. This occurs at every vertebral level. The radicular arteries follow, and supply, the anterior and posterior roots. At various vertebral levels, the segmental spinal arteries also give off segmental medullary arteries. These vessels pass directly to the longitudinally oriented vessels, reinforcing these.
Figure 15. Spinal cord blood supply
{kind=link}