Contents
What is adiponectin
Adiponectin, the most abundant hormone secreted by fat (adipose) tissue, exhibits insulin-sensitizing, antiinflammatory, antiatherogenic, proapoptotic, and antiproliferative properties 1. Adiponectin, a 244-amino acid protein secreted predominantly by white adipose tissue, is also known as AdipoQ 2, Acrp30 (adipocyte complement-related protein of 30 kDa) 3, apM1 (gene product of the adipose most abundant gene transcript-1) 4, and GBP28 (gelatin-binding protein-28) 5. Adiponectin is encoded by the APM1 gene (chromosome 3q27). In humans, adiponectin plasma levels range from 3 to 30 mcg/mL, which is among the highest plasma concentrations of a circulating protein 6. The adiponectin molecule is a 247-amino acid polypeptide and is secreted into circulation in three oligomeric isoforms: a low- molecular-weight trimer, an intermediate-molecular- weight hexamer and a high-molecular-weight complex 7. Some studies suggest that the high-molecular- weight isoform is most biologically active, and that lower levels of this form are associated with diabetes and coronary artery disease 8. Adiponectin acts through two receptors, AdipoR1 and AdipoR2; the former is expressed at higher levels in muscle tissue, and the latter, in liver tissue. Studies have demonstrated that the AdipoR1 receptor is also present in endothelial cells 9, cardiomyocytes (heart muscle cells) 10, and pancreatic beta cells 11, while AdipoR2 is present in endothelial cells 12, and both receptors are present in the hypothalamus 13. Resistance to the action of insulin resulting from obesity causes downregulation of adiponectin receptors in muscle and liver 14. Furthermore, adiponectin expression blunts increases in insulin, TNF-α, endothelin-1, and glucocorticoids, factors implicated in the pathogenesis of insulin resistance, subclinical inflammation, endothelial dysfunction, and regulation of energy metabolism, and closely related to the development of metabolic syndrome, type 2 diabetes, and cardiovascular disease 15. Accordingly, extensive research has shown that adiponectin levels are reduced in obesity 16, type 2 diabetes 16, and coronary artery disease 17.
Figure 1. Mechanisms whereby a reduction in adiponectin levels is associated with the development of metabolic syndrome

Footnote: 1: Accumulation of visceral fat reduces production of adiponectin. Tissue inflammation increases levels of C-reactive protein (CRP) and inflammatory cytokines (TNF-α and interleukin-6), activating hepatic gluconeogenesis. 2: Hepatic gluconeogenesis is activated and insulin sensitivity in muscle and liver is further reduced, resulting in increased circulating glucose levels. 3 and 4: Reduction of triglyceride oxidation from adipose tissue and dietary lipids by the liver increases levels of free fatty acids (FFA) and production of VLDL, generating an imbalance in lipid profile (increased LDL cholesterol, triglycerides [TG], and reduced HDL cholesterol). 5: Increased serum level of procollagen type I carboxy-terminal propeptide (PICP) intensifies arterial stiffness, and reduced nitric oxide production contributes to reduced vasodilation. These mechanisms, along with the pro-inflammatory environment, promote changes in blood pressure homeostasis, which contribute to the development of systemic arterial hypertension.
[Source 6 ]Circulating adiponectin levels, which are determined predominantly by genetic factors, diet, physical activity, and abdominal adiposity, are decreased in patients with diabetes, cardiovascular disease, and several obesity-associated cancers 1. Also, adiponectin levels are inversely associated with the risk of developing diabetes, cardiovascular disease, and several malignancies later in life 1. Many cancer cell lines express adiponectin receptors, and adiponectin in vitro limits cell proliferation and induces apoptosis 1. Recent in vitro studies demonstrate the antiangiogenic and tumor growth-limiting properties of adiponectin 1. Studies in both animals and humans have investigated adiponectin and adiponectin receptor regulation and expression in several cancers. Current evidence supports a role of adiponectin as a novel risk factor and potential diagnostic and prognostic biomarker in cancer. In addition, either adiponectin per se or medications that increase adiponectin levels or up-regulate signaling pathways downstream of adiponectin may prove to be useful anticancer agents.
In addition to its fat-storing capacity, adipose tissue is the largest endocrine organ regulating energy homeostasis, metabolism, inflammation, immunity, endocrine balance, and bone remodeling 18. Adipose tissue is responsible for the biosynthesis and secretion of more than 50 hormones and cytokines, known as adipocytokines or adipokines 19. Adiponectin represents the most abundant adipose-tissue protein with insulin-sensitizing, antiinflammatory, and antiatherogenic properties 18. Hypoadiponectinemia is associated not only with insulin resistance, type 2 diabetes, atherosclerosis, and coronary heart disease 20, but also with malignancies 21. Very recently, studies have shown that adiponectin is a key mediator in the development and possible progression of several types of obesity-associated cancers 22, yet the mechanism of association is still poorly understood.
What is resistin
Resistin is a 12-Da polypeptide that was initially linked to insulin resistance in animal models 23. The association between resistin and insulin resistance in humans has been controversial and inconsistent, showing positive associations in some studies 24 but not in others 25. Many studies did not find an association between resistin and measures of insulin resistance 25, 26, whereas others found a relationship that was attenuated when adjusting for adiposity 27. A lot of those studies included small numbers of participants, which limited their power. In human adipose tissue, resistin seems to be produced mainly by infiltrating macrophages 28. Circulating resistin is positively related to adiposity and may be implicated in proinflammatory signaling associated with excess adiposity 29.
What is leptin
Leptin is a hormone that helps regulate appetite by signaling hunger satisfaction (satiety). Leptin is produced primarily by fat cells (adipocytes) and to a lesser degree by other tissues, including the placenta in pregnant women. Leptin is transported in the blood to receptors in the hypothalamus in the brain. When sufficient food has been consumed, leptin tells the body that it is no longer hungry. In a normal feedback response, a low level of leptin triggers hunger and an increase in food consumption. As the level of leptin rises from an increase in fat cells, hunger diminishes and food consumption drops off.
Insufficient leptin can cause persistent hunger as the body attempts to protect itself from perceived underfeeding (starvation). Very rare inherited leptin deficiencies can cause severe obesity through constant hunger and constant eating that starts in early childhood. Leptin replacement therapy has been shown to be successful in treating some of those affected.
Obesity is most commonly associated with elevated leptin levels. This is thought to be due to a resistance to leptin that is similar to the insulin resistance often seen with obesity. People who are affected are resistant to the action of leptin—they continue to experience hunger even after consuming sufficient food. The body continues to produce more leptin in an attempt to compensate and in response to the perceived hunger. However, about 10% of those who are obese are estimated to have some degree of leptin deficiency.
There is significant interest in better understanding leptin’s ties to obesity. Obesity is a major health concern in the U.S. because it increases the risk of many conditions, such as high blood pressure (hypertension), dyslipidemias (high cholesterol and/or high triglycerides), type 2 diabetes, joint problems, sleep apnea, coronary heart disease, stroke, and some cancers. The rate of obesity has increased steadily over the last 20 years in all age ranges and, according to the Centers for Disease Control and Prevention, more than one-third of adults and 17% of children and teens in this country are currently classified as obese. Classification is based on body mass index or BMI.
A recent study found that in some people a leptin level might be more accurate than the traditional body mass index in gauging how much excess fat a person is carrying. In general, the higher the level of leptin in the bloodstream, the more fat tissue a person has. In the study, this was especially true with older women and in those with large muscles or dense bones where the results of the BMI score could be misleading.
Research is ongoing to evaluate leptin’s roles in the body and the links between leptin and obesity, and between leptin and successful weight loss. There is also continued interest in determining whether a leptin-based treatment might be useful for those who are obese and leptin-deficient.
Adiponectin function
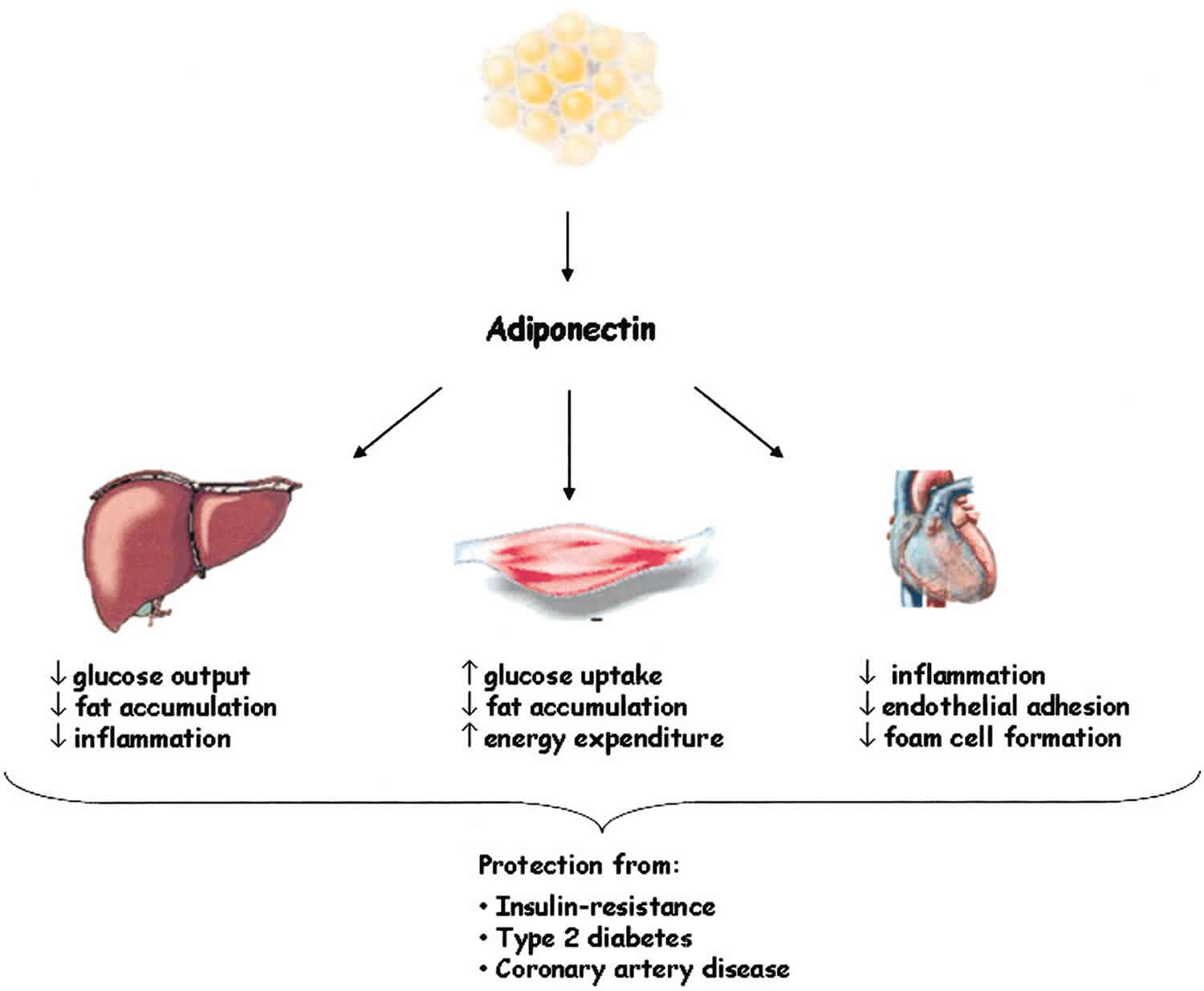
Adponectin has pleiotropic effects on a plethora of tissues and organs, with the various isoforms presenting different biological effects on different target tissues 18. Adiponectin has insulin-sensitizing, antiinflammatory, antiatherogenic, cardioprotective effects as well as distinct effects on lipid metabolism 30. Key metabolic actions include regulation of glucose and lipid metabolism through stimulation of fatty acid oxidation, suppression of hepatic glucose output, and increased insulin sensitivity in liver and skeletal muscle. High molecular weight adiponectin may be the major mediator in adiponectin’s effects, especially in the liver 31. Adiponectin can redirect fatty acids to the muscles for their oxidation, decreasing the influx of fatty acids to the liver and the total triglyceride content and leading to an improved insulin signal transduction and a higher insulin sensitivity 32. Interestingly, adiponectin reduces triglyceride content in skeletal muscle through mechanisms that involve increased expression of fatty acid transport molecules, combustion of fatty acid by acetyl-coenzyme A carboxylase, and energy dissipation by uncoupling protein 2 33. Furthermore, adiponectin enhances insulin-induced phosphorylation of the insulin receptor and the ability of insulin to activate the phosphorylation of the adaptor protein insulin receptor substrate 1 (IRS-1) 34. Adiponectin regulates pancreatic β-cell proliferation in conjunction with leptin, suggesting that adiponectin may present a direct effect on insulin secretion 35. Finally, adiponectin presents potent protective effects against insulin resistance and chronic inflammation due to its ability to improve systemic carbohydrate and lipid profiles 36.
Adiponectin has been reported to present direct antiatherogenic actions by inhibiting atherosclerosis and plaque formation. It suppresses neointimal formation by strongly inhibiting the expressions of adhesion molecules such as intracellular adhesion molecule-1, vascular cellular adhesion molecule-1, E-selectin, and the TNF-α-induced NF-κB 37. It also suppresses the uptake of cholesterol by inhibiting the expression of scavenger receptors and the foam cell formation 38. Adiponectin activates endothelial nitric oxide synthase, enhancing nitric acid production 39, and modulates vascular remodeling by suppressing smooth cell migration 40. Collectively, the antiatherogenic actions of adiponectin can be condensed as follows: adiponectin acts in the injured vascular wall by reducing the ability of macrophages to transform into foamy cells, inhibits subendothelial lipid accumulation, and stimulates vasodilatation and increased blood flow.
Aside from its peripheral actions, adiponectin may act centrally to modulate food intake and energy expenditure 41. A growing body of evidence suggests that adiponectin is an important regulator of reproductive events, with beneficial actions in ovulation, implantation process, and fetal growth and development 42. Adiponectin also plays an important role in bone homeostasis, although the precise mechanism of action remains unclear 43. Further innovative research is needed to identify and clarify the different and tissue-specific effects of this adipose tissue hormone.
Adiponectin levels
Normal plasma adiponectin levels range from 2 to 30 mcg/mL (depending on the assay methodology) 44, thus accounting for 0.01% of total plasma proteins in humans, and increase slightly with age 30. Adiponectin concentration values are 1000 times higher than leptin and cortisol levels (in the order of nanograms per milliliter) and 106 times greater than other cytokines such as IL-6 and TNF-α (in the order of picograms per milliliter) 44.
Circulating adiponectin levels are determined by various genetic, anthropometric, hormonal, inflammatory, dietary, and pharmacological factors. Unlike most of the other adipose tissue-derived proteins, serum adiponectin is reduced in obesity and generally correlates negatively with body mass index (BMI), waist and hip circumference, waist-to-hip ratio, and visceral (intraabdominal) fat rather than sc, independently of age and menopausal status 44. In addition to overall obesity, central fat distribution is an independent negative predictor of circulating adiponectin, suggesting that adiponectin may represent a link between central obesity and insulin resistance 45. Therefore, a negative correlation between circulating adiponectin and obesity, especially central obesity, insulin resistance, and type 2 diabetes has been well established 46. One possible explanation of the reduced adiponectin levels in obesity may be due to cytokines increased in obesity that contribute to the decreased adiponectin production such as TNF-α 43. Another potential mechanism indicates a negative feedback of adiponectin on its own production during the development of obesity. It has also been hypothesized that the diminished expression of sirtuin 1 (a nicotinamide adenine dinucleotide+-dependent protein deacetylase involved in adipogenesis) and Forkhead box O1 in fat tissues of obese mouse models could be implicated in the decreased adiponectin expression associated with obesity 47. Indeed, adiponectin may control its own production and probably the expression of its receptors via a regulatory feedback loop 48. Higher adiponectin is associated with weight loss 46 but predicts higher weight gain in healthy women, suggesting that elevated adiponectin production might be a sign of “healthy” adipose tissue with further capacity to accumulate fat 49. Weiss et al. 50 showed that a 10% or higher weight loss is associated with a significantly greater increase in adiponectin levels. In obese women, bariatric surgery, which leads to an average weight loss of 15 to 25%, is significantly associated with a higher increase in circulating adiponectin and a reduction in breast cancer risk 51. Moderate aerobic exercise in the absence of fat mass reduction and significant weight loss has little effect on adiponectin levels 52. Only high-intensity endurance training may improve plasma adiponectin concentrations 53.
Table 1. Reference Ranges for Adiponectin
| Body Mass Index (BMI) | Males (mcg/mL) | Females (mcg/mL) |
| <25 kg/m2 (Normal weight) | 4 to 26 | 5 to 37 |
| 25 – 30 kg/m2 (Overweight) | 4 to 20 | 5 to 28 |
| >30 kg/m2 (Obese) | 2 to 20 | 4 to 22 |
Adiponectin levels in the circulation display a diurnal variation, reaching nadir at night and peak in the morning 54, and are higher in women than men, independent of fat mass and/or fat distribution 55, most likely due to differences in circulating estrogens or androgens 56. It was demonstrated that testosterone suppresses serum total adiponectin levels in mice and men, whereas exogenous estrogen treatment or ovariectomy does not affect adiponectin concentration 57. In women without a history of diabetes, research have shown that serum adiponectin is also independently and negatively correlated with estradiol levels, but not free testosterone, cortisol, and leptin levels 45. Neither short-term fasting nor leptin administration alters serum adiponectin levels 45. In the same study, postmenopausal women presented higher adiponectin levels with lower estradiol levels compared with premenopausal women 45.
Plasma adiponectin is closely related to hormonal markers of insulin sensitivity and fasting insulinemia 58, and hypoadiponectinemia at baseline precedes a decrease in insulin sensitivity 59. Regarding other hormonal factors, adiponectin production may be down-regulated by prolactin, growth hormone, and glucocorticoids 60. Nutritional parameters can also modulate plasma adiponectin concentrations 61. Generally, chronic caloric restriction leading to weight loss increases plasma adiponectin 62. Beyond any effect of specific food item, research have shown that adherence to a Mediterranean-style dietary pattern or the Alternate Healthy Eating Index is positively associated with plasma adiponectin levels 63. The mechanisms through which the Mediterranean diet has impacts on circulating levels of adiponectin is still unknown, but several hypotheses have been raised. The omega-3 type polyunsaturated fatty acids (n-3 PUFA) found in this diet can modulate adiponectin levels by interacting with the peroxisome proliferator-activated receptors alpha (PPAR-α) and gamma (PPAR-γ) 64. Activation of peroxisome proliferator-activated receptors alpha (PPAR-α) stimulated by consumption of omega-3 PUFA (polyunsaturated fatty acid) increases expression of AdipoR1 and AdipoR2 in muscle and liver, improving sensitivity to adiponectin hormone in these tissues 65. Adiponectin then acts by reducing inflammation and oxidative stress, which ultimately leads to improved insulin sensitivity 66.
Adiponectin levels are inversely associated with glycemic load in a dose-response manner 62. Higher intakes of fiber and magnesium as well as coffee consumption have been associated with increased plasma adiponectin 62, whereas fruit consumption is related to high molecular weight adiponectin levels 67. Also, short-term walnut consumption (4 days) may increase circulating total adiponectin in obese subjects with the metabolic syndrome 68. Circulating adiponectin levels positively correlate with high-density lipoprotein (HDL) “good” cholesterol and negatively associate with triglyceride and apolipoprotein-B 69. Hypoadiponectinemia is usually associated with an atherosclerotic lipid profile 70. Adiponectin has also been linked to several inflammatory markers such as C-reactive protein (CRP) and fibrinogen 71. Several lipid-lowering drugs, such as fibrates, hydrophilic statins, and omega-3 fatty acids as well as antihypertensive drugs including angiotensin-converting enzyme inhibitors, angiotensin 2 receptor blockers, etc., may increase circulating adiponectin or improve adiponectin signaling through its receptors 72. In vitro and in vivo studies in mice and humans have indicated that adiponectin expression and secretion is up-regulated by some antidiabetic drugs such as thiazolidinediones and/or selective PPAR-γ, predominantly the high molecular weight adiponectin isoform 73. On the contrary, metformin, which significantly improves insulin sensitivity, is not capable of modulating adiponectin 74.
Finally, genetic factors such as certain polymorphisms of the ADIPOQ gene have been associated with hypoadiponectinemia in diabetic individuals 75 and posttransplantation diabetes mellitus (ADIPOQ rs1501299) in male patients receiving kidney transplants without a history of diabetes 76. In a recent large-scale meta-analysis of three genome-wide association studies for circulating adiponectin levels from population-based cohorts (n = 14,733 participants), it was reported that five single nucleotide polymorphism were genome-wide significant, whereas SNP at the adiponectin-encoding ADIPOQ locus (rs266717) demonstrated the strongest association with adiponectin levels 77. A novel variant in the ARL-15 (ADP-ribosylation factor-like 15) gene (rs4311394-G) was associated with hypoadiponectinemia, cardiovascular disease, and diabetes type 2 77 supporting a role of this novel protein ARL-15 in determining adiponectin levels. Whether these polymorphisms are also associated with risk for malignancies remains to be studied further.
Adiponectin levels in relation to disease states
Hypoadiponectinemia caused by genetic or environmental factors such as obesity and diet may play a crucial causal role in the pathogenesis of insulin resistance 20, metabolic syndrome, type 2 diabetes 78, gestational diabetes 79, hypertension, cardiovascular disease 80, and several malignancies 30. Interestingly, hypoadiponectinemia is the common denominator of the constellation of risk factors that constitute the metabolic syndrome such as hypertension, dyslipidemia, obesity, hyperglycemia, and insulin resistance. A recent meta-analysis of prospective studies including a total of 14,598 subjects and 2,623 cases of type 2 diabetes showed that higher adiponectin levels were associated with a lower risk for type 2 diabetes 81. Higher adiponectin levels were also associated with a moderate decrease in risk for coronary artery disease in diabetic men 82 and with improved glycemic control and lipid concentrations as well as reduced inflammation in diabetic women 83. The association of hypoadiponectinemia and liver disease is well documented, and a protective effect of adiponectin against fatty liver disease was proposed. Lower adiponectin levels were observed in patients suffering from chronic hepatitis with liver steatosis, correlating inversely with the grade of steatosis 84.
On the contrary, higher serum adiponectin levels were found in patients with anorexia nervosa 85 and chronic inflammatory joint diseases such as rheumatoid arthritis, whereas serum adiponectin correlated with the severity 86 of joint damage 87. Elevated adiponectin is associated with more severe proteinuria in chronic kidney disease patients, possibly underlying a protective response aimed at countering the high renal and cardiovascular risk of high proteinuria 88. Further studies are needed to elucidate the role of adiponectin in other diseases, such as ischemic cerebrovascular diseases 89, preeclampsia 90, and polycystic ovary syndrome (PCOS) 37, where there is no clear evidence of a relationship with adiponectin.
How to increase adiponectin
Only high-intensity endurance training may improve plasma adiponectin concentrations 53. Generally, chronic caloric restriction leading to weight loss increases plasma adiponectin 62. Beyond any effect of specific food item, research have shown that adherence to a Mediterranean-style dietary pattern or the Alternate Healthy Eating Index is positively associated with plasma adiponectin levels 63. Adiponectin levels are inversely associated with glycemic load in a dose-response manner 62. Higher intakes of fiber and magnesium as well as coffee consumption have been associated with increased plasma adiponectin 62, whereas fruit consumption is related to high molecular weight adiponectin levels 67. Also, short-term walnut consumption (4 days) may increase circulating total adiponectin in obese subjects with the metabolic syndrome 68. Circulating adiponectin levels positively correlate with high-density lipoprotein (HDL) “good” cholesterol and negatively associate with triglyceride and apolipoprotein-B 69. Hypoadiponectinemia is usually associated with an atherosclerotic lipid profile 70. Adiponectin has also been linked to several inflammatory markers such as C-reactive protein (CRP) and fibrinogen 71. Several lipid-lowering drugs, such as fibrates, hydrophilic statins, and omega-3 fatty acids as well as antihypertensive drugs including angiotensin-converting enzyme inhibitors, angiotensin 2 receptor blockers, etc., may increase circulating adiponectin or improve adiponectin signaling through its receptors 72. In vitro and in vivo studies in mice and humans have indicated that adiponectin expression and secretion is up-regulated by some antidiabetic drugs such as thiazolidinediones and/or selective PPAR-γ, predominantly the high molecular weight adiponectin isoform 73. On the contrary, metformin, which significantly improves insulin sensitivity, is not capable of modulating adiponectin 74.
In obese women, bariatric surgery, which leads to an average weight loss of 15 to 25%, is significantly associated with a higher increase in circulating adiponectin and a reduction in breast cancer risk 51. Moderate aerobic exercise in the absence of fat mass reduction and significant weight loss has little effect on adiponectin levels 52.
- Dalamaga M, Diakopoulos KN, Mantzoros CS. The role of adiponectin in cancer: a review of current evidence. Endocr Rev. 2012;33(4):547-94. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3410224/[↩][↩][↩][↩][↩]
- AdipoQ is a novel adipose-specific gene dysregulated in obesity. Hu E, Liang P, Spiegelman BM. J Biol Chem. 1996 May 3; 271(18):10697-703.[↩]
- Scherer PE, Williams S, Fogliano M, Baldini G, Lodish HF. 1995. A novel serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem 270:26746–26749[↩]
- Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y, Matsubara K. 1996. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem Biophys Res Commun 221:286–289[↩]
- Nakano Y, Tobe T, Choi-Miura NH, Mazda T, Tomita M. 1996. Isolation and characterization of GBP28, a novel gelatin-binding protein purified from human plasma. J Biochem 120:803–812[↩]
- Relationships between adiponectin levels, the metabolic syndrome, and type 2 diabetes: a literature review. Arch. Endocrinol. Metab. vol.61 no.6 São Paulo Dec. 2017 http://www.scielo.br/pdf/aem/v61n6/2359-3997-aem-61-06-0614.pdf[↩][↩]
- Pajvani UB, Du X, Combs TP, Berg AH, Rajala MW, Schulthess T, et al. Structure-function studies of the adipocyte-secreted hormone Acrp30/adiponectin. Implications fpr metabolic regulation and bioactivity. J Biol Chem. 2003;278(11):9073-85.[↩]
- Aso Y, Yamamoto R, Wakabayashi S, Uchida T, Takayanagi K, Takebayashi K, et al. Comparison of serum high-molecular weight (HMW) adiponectin with total adiponectin concentrations in type 2 diabetic patients with coronary artery disease using a novel enzyme-linked immunosorbent assay to detect HMW adiponectin. Diabetes. 2006;55(7):1954-60.[↩]
- Motoshima H, Wu X, Mahadev K, Goldstein BJ. Adiponectin suppresses proliferation and superoxide generation and enhances eNOS activity in endothelial cells treated with oxidized LDL. Biochem Biophys Res Commun. 2004;315(2):264-71.[↩]
- Pineiro R, Iglesias MJ, Gallego R, Raghay K, Eiras S, Rubio J, et al. Adiponectin is synthesized and secreted by human and murine cardiomyocytes. FEBS Lett. 2005;579(23):5163-9.[↩]
- Kharroubi I, Rasschaert J, Eizirik DL, Cnop M. Expression of adiponectin receptors in pancreatic beta cells. Biochem Biophys Res Commun. 2003;312(4):1118-22[↩]
- Tan KC, Xu A, Chow WS, Lam MC, Ai VH, Tam SC, et al. Hypoadiponectinemia is associated with impaired endothelium-dependent vasodilation. J Clin Endocrinol Metab. 2004;89(2):765-9.[↩]
- Coope A, Milanski M, Araujo EP, Tambascia M, Saad MJ, Geloneze B, et al. AdipoR1 mediates the anorexigenic and insulin/leptin-like actions of adiponectin in the hypothalamus. FEBS letters. 2008;582(10):1471-6.[↩]
- Tsuchida A, Yamauchi T, Ito Y, Hada Y, Maki T, Takekawa S, et al. Insulin/Foxo1 pathway regulates expression levels of adiponectin receptors and adiponectin sensitivity. J Biol Chem. 2004;279(29):30817-22.[↩]
- Gil-Campos M, Canete RR, Gil A. Adiponectin, the missing link in insulin resistance and obesity. Clin Nutr. 2004;23(5):963-74.[↩]
- Heid IM, Wagner SA, Gohlke H, Iglseder B, Mueller JC, Cip P, et al. Genetic architecture of the APM1 gene and its influence on adiponectin plasma levels and parameters of the metabolic syndrome in 1,727 healthy Caucasians. Diabetes. 2006;55(2):375-84.[↩][↩]
- Oliveira CS, Saddi-Rosa P, Crispim F, Canani LH, Gerchman F, Giuffrida FM, et al. Association of ADIPOQ variants, total and high molecular weight adiponectin levels with coronary artery disease in diabetic and non-diabetic Brazilian subjects. J Diabetes Complications. 2012;26(2):94-8.[↩]
- Ziemke F, Mantzoros CS. 2010. Adiponectin in insulin resistance: lessons from translational research. Am J Clin Nutr 91:258S–261S[↩][↩][↩]
- MacDougald OA, Burant CF. 2007. The rapidly expanding family of adipokines. Cell Metab 6:159–161[↩]
- Hivert MF, Sullivan LM, Fox CS, Nathan DM, D’Agostino RB, Sr, Wilson PW, Meigs JB. 2008. Associations of adiponectin, resistin, and tumor necrosis factor-α with insulin resistance. J Clin Endocrinol Metab 93:3165–3172[↩][↩]
- Kelesidis I, Kelesidis T, Mantzoros CS. 2006. Adiponectin and cancer: a systematic review. Br J Cancer 94:1221–1225[↩]
- Paz-Filho G, Lim EL, Wong ML, Licinio J. 2011. Associations between adipokines and obesity-related cancer. Front Biosci 16:1634–1650[↩]
- The hormone resistin links obesity to diabetes. Steppan CM, Bailey ST, Bhat S, Brown EJ, Banerjee RR, Wright CM, Patel HR, Ahima RS, Lazar MA. Nature. 2001 Jan 18; 409(6818):307-12.[↩]
- Osawa H, Tabara Y, Kawamoto R, Ohashi J, Ochi M, Onuma H, Nishida W, Yamada K, Nakura J, Kohara K, Miki T, Makino H 2007 Plasma resistin, associated with single nucleotide polymorphism-420, is correlated with insulin resistance, lower HDL cholesterol, and high-sensitivity C-reactive protein in the Japanese general population. Diabetes Care 30:1501–1506[↩]
- Vozarova de Court, Degawa-Yamauchi M, Considine RV, Tataranni PA 2004 High serum resistin is associated with an increase in adiposity but not a worsening of insulin resistance in Pima Indians. Diabetes 53:1279–1284[↩][↩]
- Lee JH, Chan JL, Yiannakouris N, Kontogianni M, Estrada E, Seip R, Orlova C, Mantzoros CS 2003 Circulating resistin levels are not associated with obesity or insulin resistance in humans and are not regulated by fasting or leptin administration: cross-sectional and interventional studies in normal, insulin-resistant, and diabetic subjects. J Clin Endocrinol Metab 88:4848–4856[↩]
- Degawa-Yamauchi M, Bovenkerk JE, Juliar BE, Watson W, Kerr K, Jones R, Zhu Q, Considine RV 2003 Serum resistin (FIZZ3) protein is increased in obese humans. J Clin Endocrinol Metab 88:5452–5455[↩]
- Curat CA, Wegner V, Sengenes C, Miranville A, Tonus C, Busse R, Bouloumie A 2006 Macrophages in human visceral adipose tissue: increased accumulation in obesity and a source of resistin and visfatin. Diabetologia 49:744–747[↩]
- McTernan PG, Kusminski CM, Kumar S 2006 Resistin. Curr Opin Lipidol 17:170–175[↩]
- Barb D, Williams CJ, Neuwirth AK, Mantzoros CS. 2007. Adiponectin in relation to malignancies: a review of existing basic research and clinical evidence. Am J Clin Nutr 86:s858–s866[↩][↩][↩]
- Hada Y, Yamauchi T, Waki H, Tsuchida A, Hara K, Yago H, Miyazaki O, Ebinuma H, Kadowaki T. 2007. Selective purification and characterization of adiponectin multimer species from human plasma. Biochem Biophys Res Commun 356:487–493[↩]
- Yamauchi T, Kamon J, Waki H, Terauchi Y, Kubota N, Hara K, Mori Y, Ide T, Murakami K, Tsuboyama-Kasaoka N, Ezaki O, Akanuma Y, Gavrilova O, Vinson C, Reitman ML, Kagechika H, Shudo K, Yoda M, Nakano Y, Tobe K, Nagai R, Kimura S, Tomita M, Froguel P, Kadowaki T. 2001. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med 7:941–946[↩]
- Kadowaki T, Yamauchi T, Kubota N, Hara K, Ueki K, Tobe K. 2006. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J Clin Invest 116:1784–1792[↩]
- Wang C, Mao X, Wang L, Liu M, Wetzel MD, Guan KL, Dong LQ, Liu F. 2007. Adiponectin sensitizes insulin signaling by reducing p70 S6 kinase-mediated serine phosphorylation of IRS-1. J Biol Chem 282:7991–7996[↩]
- Kharroubi I, Rasschaert J, Eizirik DL, Cnop M. 2003. Expression of adiponectin receptors in pancreatic β cells. Biochem Biophys Res Commun 312:1118–1122[↩]
- Park J, Euhus DM, Scherer PE. 2011. Paracrine and endocrine effects of adipose tissue on cancer development and progression. Endocr Rev 32:550–570[↩]
- Brochu-Gaudreau K, Rehfeldt C, Blouin R, Bordignon V, Murphy BD, Palin MF. 2010. Adiponectin action from head to toe. Endocrine 37:11–32[↩][↩]
- Ouchi N, Kihara S, Arita Y, Nishida M, Matsuyama A, Okamoto Y, Ishigami M, Kuriyama H, Kishida K, Nishizawa H, Hotta K, Muraguchi M, Ohmoto Y, Yamashita S, Funahashi T, Matsuzawa Y. 2001. Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocyte-derived macrophages. Circulation 103:1057–1063[↩]
- Chen H, Montagnani M, Funahashi T, Shimomura I, Quon MJ. 2003. Adiponectin stimulates production of nitric oxide in vascular endothelial cells. J Biol Chem 278:45021–45026[↩]
- Arita Y, Kihara S, Ouchi N, Maeda K, Kuriyama H, Okamoto Y, Kumada M, Hotta K, Nishida M, Takahashi M, Nakamura T, Shimomura I, Muraguchi M, Ohmoto Y, Funahashi T, Matsuzawa Y. 2002. Adipocyte-derived plasma protein adiponectin acts as a platelet-derived growth factor-BB-binding protein and regulates growth factor-induced common postreceptor signal in vascular smooth muscle cell. Circulation 105:2893–2898[↩]
- Kubota N, Yano W, Kubota T, Yamauchi T, Itoh S, Kumagai H, Kozono H, Takamoto I, Okamoto S, Shiuchi T, Suzuki R, Satoh H, Tsuchida A, Moroi M, Sugi K, Noda T, Ebinuma H, Ueta Y, Kondo T, Araki E, Ezaki O, Nagai R, Tobe K, Terauchi Y, Ueki K, Minokoshi Y, Kadowaki T. 2007. Adiponectin stimulates AMP-activated protein kinase in the hypothalamus and increases food intake. Cell Metab 6:55–68[↩]
- Corbetta S, Bulfamante G, Cortelazzi D, Barresi V, Cetin I, Mantovani G, Bondioni S, Beck-Peccoz P, Spada A. 2005. Adiponectin expression in human fetal tissues during mid- and late gestation. J Clin Endocrinol Metab 90:2397–2402[↩]
- Tilg H, Moschen AR. 2006. Adipocytokines: mediators linking adipose tissue, inflammation and immunity. Nat Rev Immunol 6:772–783[↩][↩]
- Ziemke F, Mantzoros CS. 2010. Adiponectin in insulin resistance: lessons from translational research. Am J Clin Nutr 91:258S–261S https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2793112/[↩][↩][↩]
- Gavrila A, Chan JL, Yiannakouris N, Kontogianni M, Miller LC, Orlova C, Mantzoros CS. 2003. Serum adiponectin levels are inversely associated with overall and central fat distribution but are not directly regulated by acute fasting or leptin administration in humans: cross-sectional and interventional studies. J Clin Endocrinol Metab 88:4823–4831[↩][↩][↩][↩]
- Chandran M, Phillips SA, Ciaraldi T, Henry RR. 2003. Adiponectin: more than just another fat cell hormone? Diabetes Care 26:2442–2450[↩][↩]
- Qiao L, Shao J. 2006. SIRT1 regulates adiponectin gene expression through Foxo1-C/enhancer-binding protein α transcriptional complex. J Biol Chem 281:39915–39924[↩]
- Bauche IB, Ait El Mkadem S, Rezsohazy R, Funahashi T, Maeda N, Miranda LM, Brichard SM. 2006. Adiponectin downregulates its own production and the expression of its AdipoR2 receptor in transgenic mice. Biochem Biophys Res Commun 345:1414–1424[↩]
- Hivert MF, Sun Q, Shrader P, Mantzoros CS, Meigs JB, Hu FB. 2011. Higher adiponectin levels predict greater weight gain in healthy women in the Nurses’ Health Study. Obesity (Silver Spring) 19:409–415[↩]
- Weiss EP, Racette SB, Villareal DT, Fontana L, Steger-May K, Schechtman KB, Klein S, Holloszy JO. 2006. Improvements in glucose tolerance and insulin action induced by increasing energy expenditure or decreasing energy intake: a randomized controlled trial. Am J Clin Nutr 84:1033–1042[↩]
- Sjöström L, Gummesson A, Sjöström CD, Narbro K, Peltonen M, Wedel H, Bengtsson C, Bouchard C, Carlsson B, Dahlgren S, Jacobson P, Karason K, Karlsson J, Larsson B, Lindroos AK, Lönroth H, Näslund I, Olbers T, Stenlöf K, Torgerson J, Carlsson LM. 2009. Effects of bariatric surgery on cancer incidence in obese patients in Sweden (Swedish Obese Subjects Study): a prospective, controlled intervention trial. Lancet Oncol 10:653–662[↩][↩]
- Friedenreich CM, Neilson HK, Woolcott CG, McTiernan A, Wang Q, Ballard-Barbash R, Jones CA, Stanczyk FZ, Brant RF, Yasui Y, Irwin ML, Campbell KL, McNeely ML, Karvinen KH, Courneya KS. 2011. Changes in insulin resistance indicators, IGFs, and adipokines in a year-long trial of aerobic exercise in postmenopausal women. Endocr Relat Cancer 18:357–369[↩][↩]
- Moghadasi M, Mohebbi H, Rahmani-Nia F, Hassan-Nia S, Noroozi H, Pirooznia N. 2012. High-intensity endurance training improves adiponectin mRNA and plasma concentrations. Eur J Appl Physiol 112:1207–1214[↩][↩]
- Gavrila A, Peng CK, Chan JL, Mietus JE, Goldberger AL, Mantzoros CS. 2003. Diurnal and ultradian dynamics of serum adiponectin in healthy men: comparison with leptin, circulating soluble leptin receptor, and cortisol patterns. J Clin Endocrinol Metab 88:2838–2843[↩]
- Cnop M, Havel PJ, Utzschneider KM, Carr DB, Sinha MK, Boyko EJ, Retzlaff BM, Knopp RH, Brunzell JD, Kahn SE. 2003. Relationship of adiponectin to body fat distribution, insulin sensitivity and plasma lipoproteins: evidence for independent roles of age and sex. Diabetologia 46:459–469[↩]
- Nishizawa H, Shimomura I, Kishida K, Maeda N, Kuriyama H, Nagaretani H, Matsuda M, Kondo H, Furuyama N, Kihara S, Nakamura T, Tochino Y, Funahashi T, Matsuzawa Y. 2002. Androgens decrease plasma adiponectin, an insulin-sensitizing adipocyte-derived protein. Diabetes 51:2734–2741[↩]
- Sieminska L, Wojciechowska C, Niedziolka D, Marek B, Kos-Kudla B, Kajdaniuk D, Nowak M. 2005. Effect of postmenopause and hormone replacement therapy on serum adiponectin levels. Metabolism 54:1610–1614[↩]
- Hroussalas G, Kassi E, Dalamaga M, Delimaris I, Zachari A, Dionyssiou-Asteriou A. 2008. Leptin, soluble leptin receptor, adiponectin and resistin in relation to OGTT in overweight/obese postmenopausal women. Maturitas 59:339–349[↩]
- Stefan N, Vozarova B, Funahashi T, Matsuzawa Y, Weyer C, Lindsay RS, Youngren JF, Havel PJ, Pratley RE, Bogardus C, Tataranni PA. 2002. Plasma adiponectin concentration is associated with skeletal muscle insulin receptor tyrosine phosphorylation, and low plasma concentration precedes a decrease in whole-body insulin sensitivity in humans. Diabetes 51:1884–1888[↩]
- Swarbrick MM, Havel PJ. 2008. Physiological, pharmacological, and nutritional regulation of circulating adiponectin concentrations in humans. Metab Syndr Relat Disord 6:87–102[↩]
- Pischon T, Girman CJ, Rifai N, Hotamisligil GS, Rimm EB. 2005. Association between dietary factors and plasma adiponectin concentrations in men. Am J Clin Nutr 81:780–786[↩]
- Qi L, Rimm E, Liu S, Rifai N, Hu FB. 2005. Dietary glycemic index, glycemic load, cereal fiber, and plasma adiponectin concentration in diabetic men. Diabetes Care 28:1022–1028[↩][↩][↩][↩][↩][↩]
- Fargnoli JL, Fung TT, Olenczuk DM, Chamberland JP, Hu FB, Mantzoros CS. 2008. Adherence to healthy eating patterns is associated with higher circulating total and high-molecular-weight adiponectin and lower resistin concentrations in women from the Nurses’ Health Study. Am J Clin Nutr 88:1213–1224[↩][↩]
- Neschen S, Morino K, Rossbacher JC, Pongratz RL, Cline GW, Sono S, et al. Fish oil regulates adiponectin secretion by a peroxisome proliferator-activated receptor-gamma-dependent mechanism in mice. Diabetes. 2006;55(4):924-8.[↩]
- Yamauchi T, Kadowaki T Physiological and pathophysiological roles of adiponectin and adiponectin receptors in the integrated regulation of metabolic and cardiovascular diseases. Int J Obes (Lond). 2008;32 Suppl 7:S13-8.[↩]
- Tsuchida A, Yamauchi T, Takekawa S, Hada Y, Ito Y, Maki T, et al. Peroxisome proliferator-activated receptor (PPAR)alpha activation increases adiponectin receptors and reduces obesity-related inflammation in adipose tissue: comparison of activation of PPARalpha, PPARgamma, and their combination. Diabetes. 2005;54(12):3358-70.[↩]
- Yannakoulia M, Yiannakouris N, Melistas L, Fappa E, Vidra N, Kontogianni MD, Mantzoros CS. 2008. Dietary factors associated with plasma high molecular weight and total adiponectin levels in apparently healthy women. Eur J Endocrinol 159:R5–R10[↩][↩]
- Aronis KN, Vamvini MT, Chamberland JP, Sweeney LL, Brennan AM, Magkos F, Mantzoros CS. 2012. Short-term walnut consumption increases circulating total adiponectin and apolipoprotein A concentrations, but does not affect markers of inflammation or vascular injury in obese humans with the metabolic syndrome: data from a double-blinded, randomized, placebo-controlled study. Metabolism 61:577–582[↩][↩]
- Kassi E, Dalamaga M, Hroussalas G, Kazanis K, Merantzi G, Zachari A, Giamarellos-Bourboulis EJ, Dionyssiou-Asteriou A. 2010. Adipocyte factors, high-sensitive C-reactive protein levels and lipoxidative stress products in overweight postmenopausal women with normal and impaired OGTT. Maturitas 67:72–77[↩][↩]
- Okada T, Saito E, Kuromori Y, Miyashita M, Iwata F, Hara M, Harada K. 2006. Relationship between serum adiponectin level and lipid composition in each lipoprotein fraction in adolescent children. Atherosclerosis 188:179–183[↩][↩]
- Schulze MB, Rimm EB, Shai I, Rifai N, Hu FB. 2004. Relationship between adiponectin and glycemic control, blood lipids, and inflammatory markers in men with type 2 diabetes. Diabetes Care 27:1680–1687[↩][↩]
- Wanders D, Plaisance EP, Judd RL. 2010. Pharmacological effects of lipid-lowering drugs on circulating adipokines. World J Diabetes 1:116–128[↩][↩]
- Combs TP, Wagner JA, Berger J, Doebber T, Wang WJ, Zhang BB, Tanen M, Berg AH, O’Rahilly S, Savage DB, Chatterjee K, Weiss S, Larson PJ, Gottesdiener KM, Gertz BJ, Charron MJ, Scherer PE, Moller DE. 2002. Induction of adipocyte complement-related protein of 30 kilodaltons by PPARγ agonists: a potential mechanism of insulin sensitization. Endocrinology 143:998–1007[↩][↩]
- Hanefeld M, Pfützner A, Forst T, Kleine I, Fuchs W. 2011. Double-blind, randomized, multicentre, and active comparator controlled investigation of the effect of pioglitazone, metformin, and the combination of both on cardiovascular risk in patients with type 2 diabetes receiving stable basal insulin therapy: the PIOCOMB study. Cardiovasc Diabetol 10:65.[↩][↩]
- Waki H, Yamauchi T, Kamon J, Ito Y, Uchida S, Kita S, Hara K, Hada Y, Vasseur F, Froguel P, Kimura S, Nagai R, Kadowaki T. 2003. Impaired multimerization of human adiponectin mutants associated with diabetes. Molecular structure and multimer formation of adiponectin. J Biol Chem 278:40352–40363[↩]
- Kang ES, Magkos F, Kim BS, Zhai R, Su L, Kim YS, Christiani DC, Lee HC, Mantzoros CS. 2012. Variants of the adiponectin and adiponectin receptor-1 Genes and posttransplantation diabetes mellitus in renal allograft recipients. J Clin Endocrinol Metab 97:E129–E135[↩]
- Richards JB, Waterworth D, O’Rahilly S, Hivert MF, Loos RJ, Perry JR, Tanaka T, Timpson NJ, Semple RK, Soranzo N, Song K, Rocha N, Grundberg E, Dupuis J, Florez JC, Langenberg C, Prokopenko I, Saxena R, Sladek R, Aulchenko Y, Evans D, Waeber G, Erdmann J, Burnett MS, Sattar N, Devaney J, Willenborg C, Hingorani A, Witteman JC, Vollenweider P, Glaser B, Hengstenberg C, Ferrucci L, Melzer D, Stark K, Deanfield J, Winogradow J, Grassl M, Hall AS, Egan JM, Thompson JR, Ricketts SL, König IR, Reinhard W, Grundy S, Wichmann HE, Barter P, Mahley R, Kesaniemi YA, Rader DJ, Reilly MP, Epstein SE, Stewart AF, Van Duijn CM, Schunkert H, Burling K, Deloukas P, Pastinen T, Samani NJ, McPherson R, Davey Smith G, Frayling TM, Wareham NJ, Meigs JB, Mooser V, Spector TD. 2009. A genome-wide association study reveals variants in ARL15 that influence adiponectin levels. PLoS Genet 5:e1000768[↩][↩]
- Heidemann C, Sun Q, van Dam RM, Meigs JB, Zhang C, Tworoger SS, Mantzoros CS, Hu FB. 2008. Total and high-molecular-weight adiponectin and resistin in relation to the risk for type 2 diabetes in women. Ann Intern Med 149:307–316[↩]
- Mazaki-Tovi S, Romero R, Vaisbuch E, Erez O, Mittal P, Chaiworapongsa T, Kim SK, Pacora P, Yeo L, Gotsch F, Dong Z, Yoon BH, Hassan SS, Kusanovic JP. 2009. Maternal serum adiponectin multimers in gestational diabetes. J Perinat Med 37:637–650[↩]
- Trujillo ME, Scherer PE. 2005. Adiponectin—journey from an adipocyte secretory protein to biomarker of the metabolic syndrome. J Intern Med 257:167–175[↩]
- Li S, Shin HJ, Ding EL, van Dam RM. 2009. Adiponectin levels and risk of type 2 diabetes: a systematic review and meta-analysis. JAMA 302:179–188[↩]
- Schulze MB, Shai I, Rimm EB, Li T, Rifai N, Hu FB. 2005. Adiponectin and future coronary heart disease events among men with type 2 diabetes. Diabetes 54:534–539[↩]
- Mantzoros CS, Li T, Manson JE, Meigs JB, Hu FB. 2005. Circulating adiponectin levels are associated with better glycemic control, more favorable lipid profile, and reduced inflammation in women with type 2 diabetes. J Clin Endocrinol Metab 90:4542–4548[↩]
- Durante-Mangoni E, Zampino R, Marrone A, Tripodi MF, Rinaldi L, Restivo L, Cioffi M, Ruggiero G, Adinolfi LE. 2006. Hepatic steatosis and insulin resistance are associated with serum imbalance of adiponectin/tumour necrosis factor-α in chronic hepatitis C patients. Aliment Pharmacol Ther 24:1349–1357[↩]
- Delporte ML, Brichard SM, Hermans MP, Beguin C, Lambert M. 2003. Hyperadiponectinaemia in anorexia nervosa. Clin Endocrinol (Oxf) 58:22–29[↩]
- Ebina K, Fukuhara A, Ando W, Hirao M, Koga T, Oshima K, Matsuda M, Maeda K, Nakamura T, Ochi T, Shimomura I, Yoshikawa H, Hashimoto J. 2009. Serum adiponectin concentrations correlate with severity of rheumatoid arthritis evaluated by extent of joint destruction. Clin Rheumatol 28:445–451[↩]
- Senolt L, Pavelka K, Housa D, Haluzík M. 2006. Increased adiponectin is negatively linked to the local inflammatory process in patients with rheumatoid arthritis. Cytokine 35:247–252[↩]
- Zoccali C, Mallamaci F. 2011. Adiponectin and leptin in chronic kidney disease: causal factors or mere risk markers? J Ren Nutr 21:87–91[↩]
- Chen MP, Tsai JC, Chung FM, Yang SS, Hsing LL, Shin SJ, Lee YJ. 2005. Hypoadiponectinemia is associated with ischemic cerebrovascular disease. Arterioscler Thromb Vasc Biol 25:821–826[↩]
- Dalamaga M, Srinivas SK, Elovitz MA, Chamberland J, Mantzoros CS. 2011. Serum adiponectin and leptin in relation to risk for preeclampsia: results from a large case-control study. Metabolism 60:1539–1544[↩]



{kind=link}