Contents
What is basophils
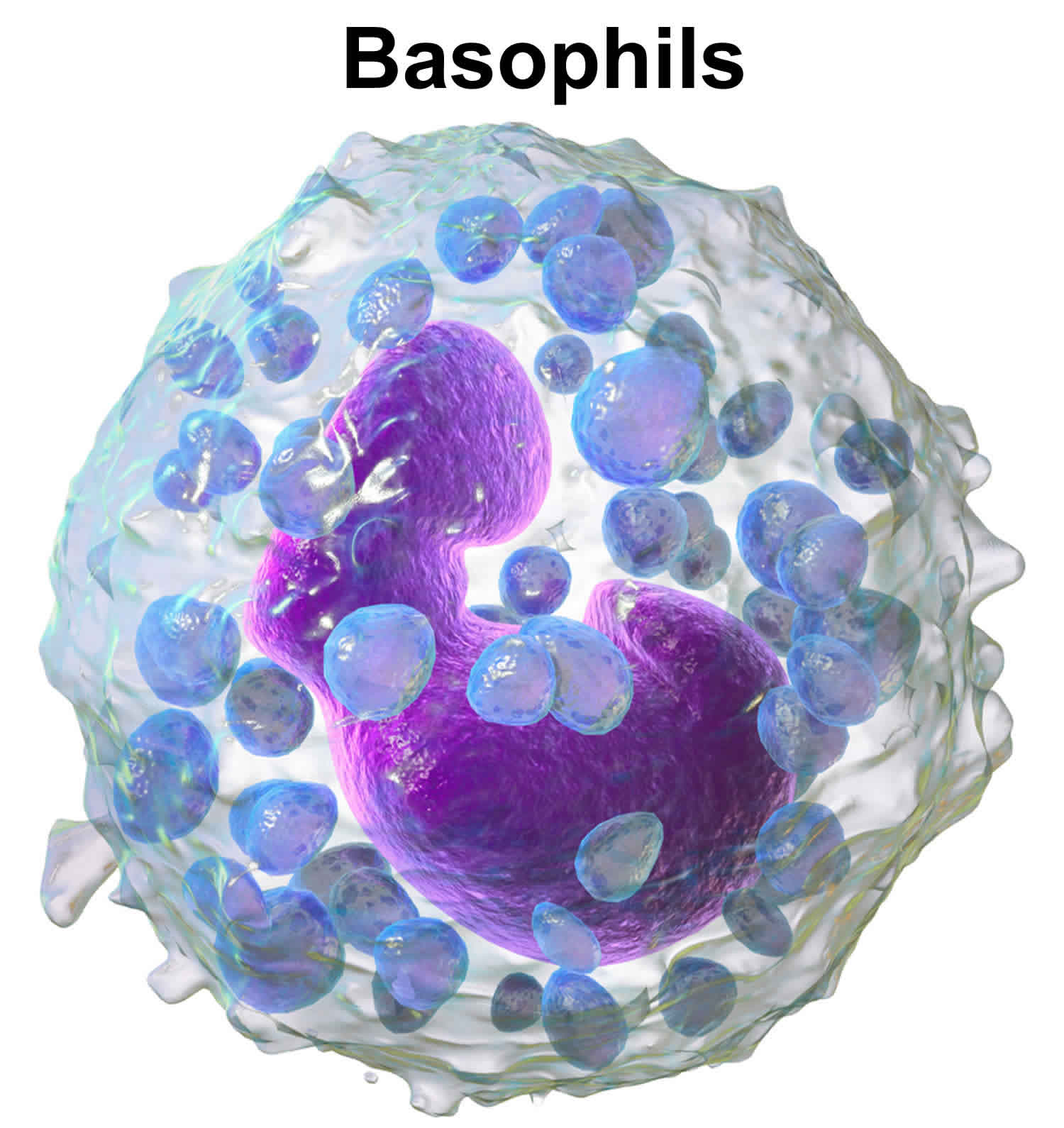
Basophils are effector cells of the innate immune system that are associated with allergic inflammation and infections with helminth (worm) parasites 1. Basophils are the least abundant white blood cells (leucocytes) primarily found in the blood circulation. Basophils comprise only a small percentage (∼ 0·5%) of circulating blood cells under steady-state conditions, but rapidly expand in the bone marrow in response to inflammatory signals and are mobilized to the blood, spleen, lung and liver. Basophils are generated from the granulocyte–monocyte progenitors in the bone marrow and populate the periphery as fully mature cells 2. The lifespan of basophils is short; recently estimated to be in the range of 1–2 days 3. Thus, it is thought that basophil differentiation from precursor populations is continually occurring in order to maintain basophil populations in the periphery. Basophils are rarely found in normal tissue; however, they accumulate in sites of inflammation 4, where they facilitate secretion of histamine, leukotrienes, and cytokines 5. Several studies have demonstrated that basophils are pivotal for T helper 2 (Th2) immune responses and rapid generation of interleukin (IL)-4 and IL-13 in both humans 6. In addition, basophil-derived IL-4 (interlukin-4) has been shown to differentiate naïve CD4 T cells into T helper 2 (Th2) effector cells, which play key roles in eliciting and maintaining allergic responses 7. Basophils and mast cells may be derived from common precursor cells, which share similar phenotypic and functional characteristics. For example, these cells express FcεRI on their surfaces, and both cells are central effectors in Th2 immune reactions 8. Despite sharing a number of phenotypic and functional properties, basophils and mast cells exhibit different activities. Although they play roles in allergic reactions, mast cells are known as the major effector cells in the immediate hypersensitivity reaction, whereas basophils are recruited to inflammatory sites, where they act to enhance the inflammatory process and thus may be associated with the severity of allergic diseases 9.
Although present in very small numbers at baseline levels, basophil populations can significantly expand in response to certain factors, such as IL-3 (interkukin-3) and granulocyte macrophage colony-stimulating factor 10. Interkukin-3 (IL-3) promotes the differentiation of basophils from bone marrow cells (Figure 3), induces the generation of basophils in mice, and IL-3 signals are necessary for infection induced basophilia following Nippostrongylus brasiliensis and Strongyloides venezuelensis infections 10. In addition, IL-3 (interkukin-3) can augment many aspects of basophil function including the release of effector molecules after IgE-dependent stimulation 11. Collectively, these studies supported the hypothesis that basophil responses were critically dependent on IL-3 signaling.
For many years basophils have been a somewhat enigmatic immune cell type and questions regarding their role in protective immunity as well as the specific pathogens or insults that elicit basophil responses are not fully answered 12. In addition to their scarcity and short lifespan, studies of basophils have been hindered by the lack of information about how basophils can be distinguished both phenotypically and functionally from the developmentally related mast cells. Both mast cells and basophils express the high-affinity receptor for IgE and can release a similar spectrum of mediators upon IgE cross-linking 13.
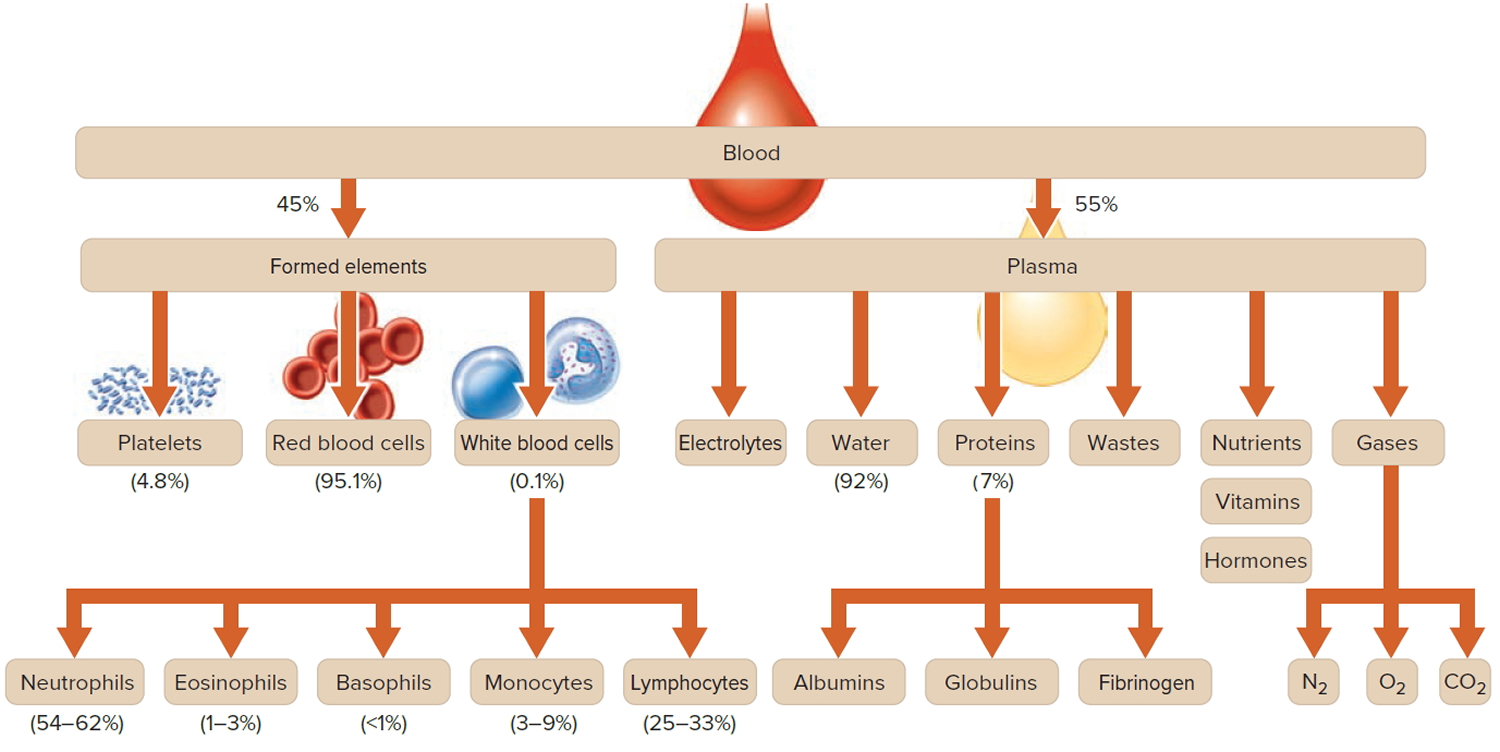
Figure 1. Blood composition
Note: Blood is a complex mixture of formed elements in a liquid extracellular matrix, called blood plasma. Note that water and proteins account for 99% of the blood plasma.
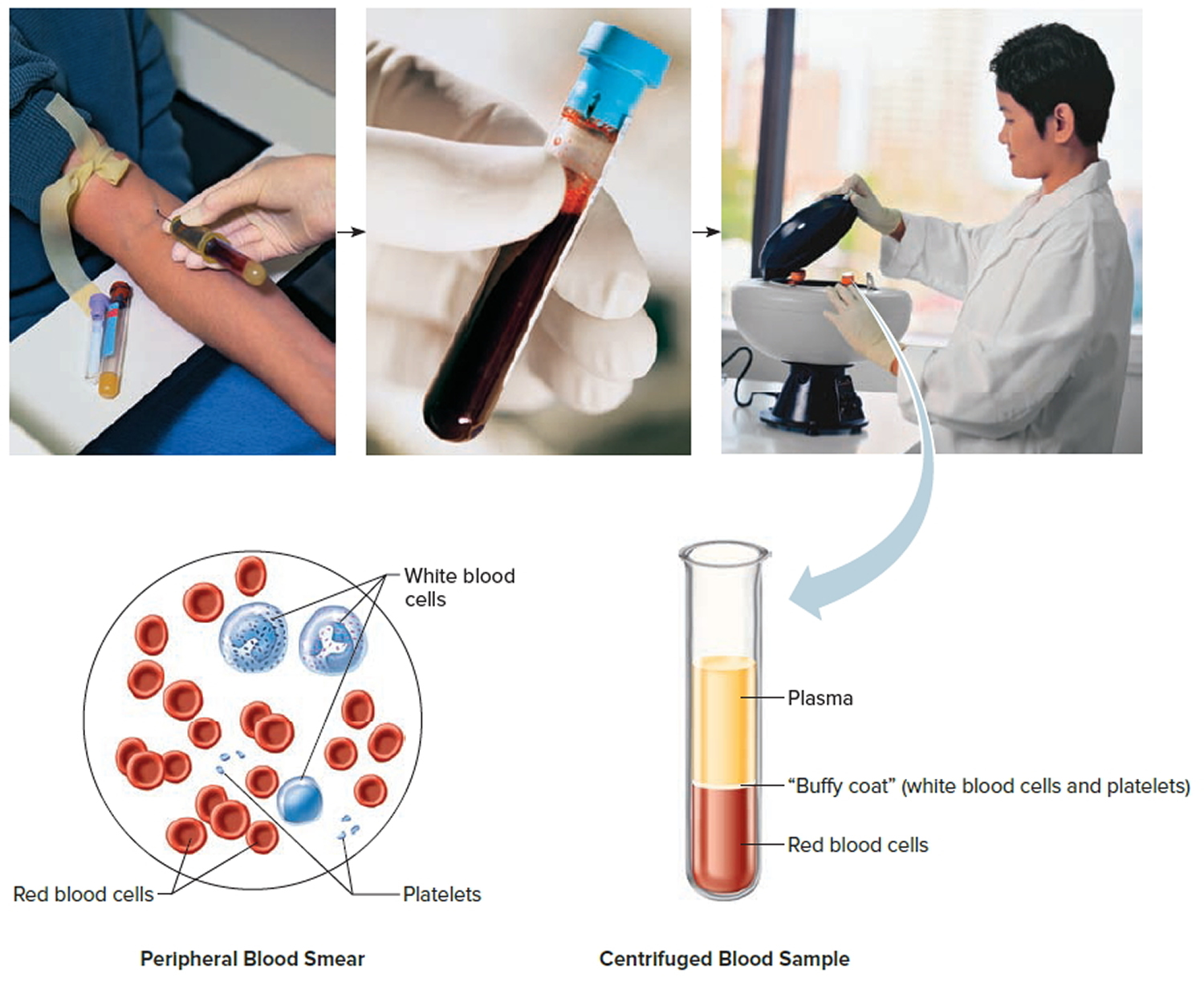
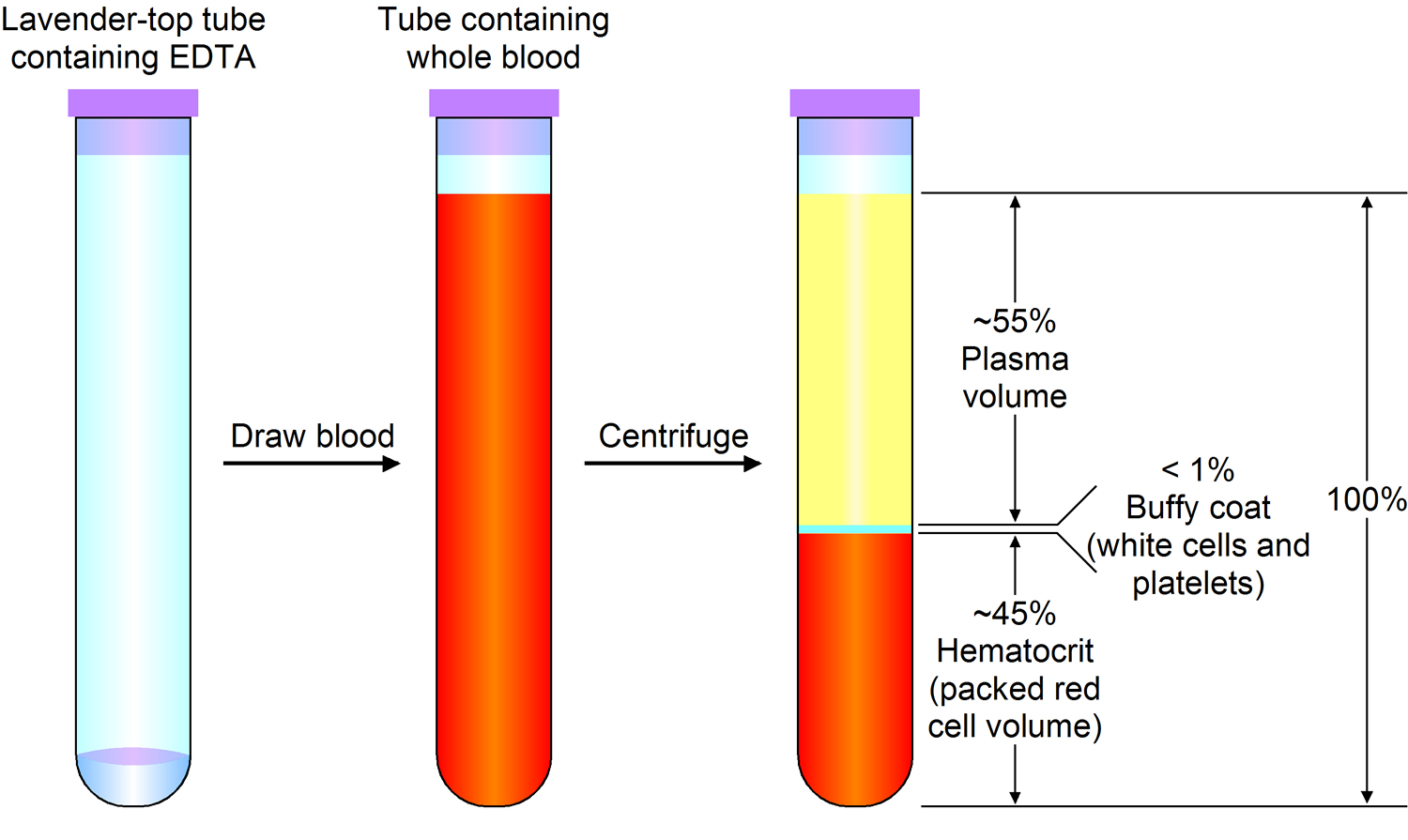
Note: Blood consists of a liquid portion called plasma and a solid portion (the formed elements) that includes red blood cells, white blood cells, and platelets. When blood components are separated by centrifugation, the white blood cells and platelets form a thin layer, called the “buffy coat,” between the plasma and the red blood cells, which accounts for about 1% of the total blood volume. Blood cells and platelets can be seen under a light microscope when a blood sample is smeared onto a glass slide.
Figure 2. Bone marrow anatomy
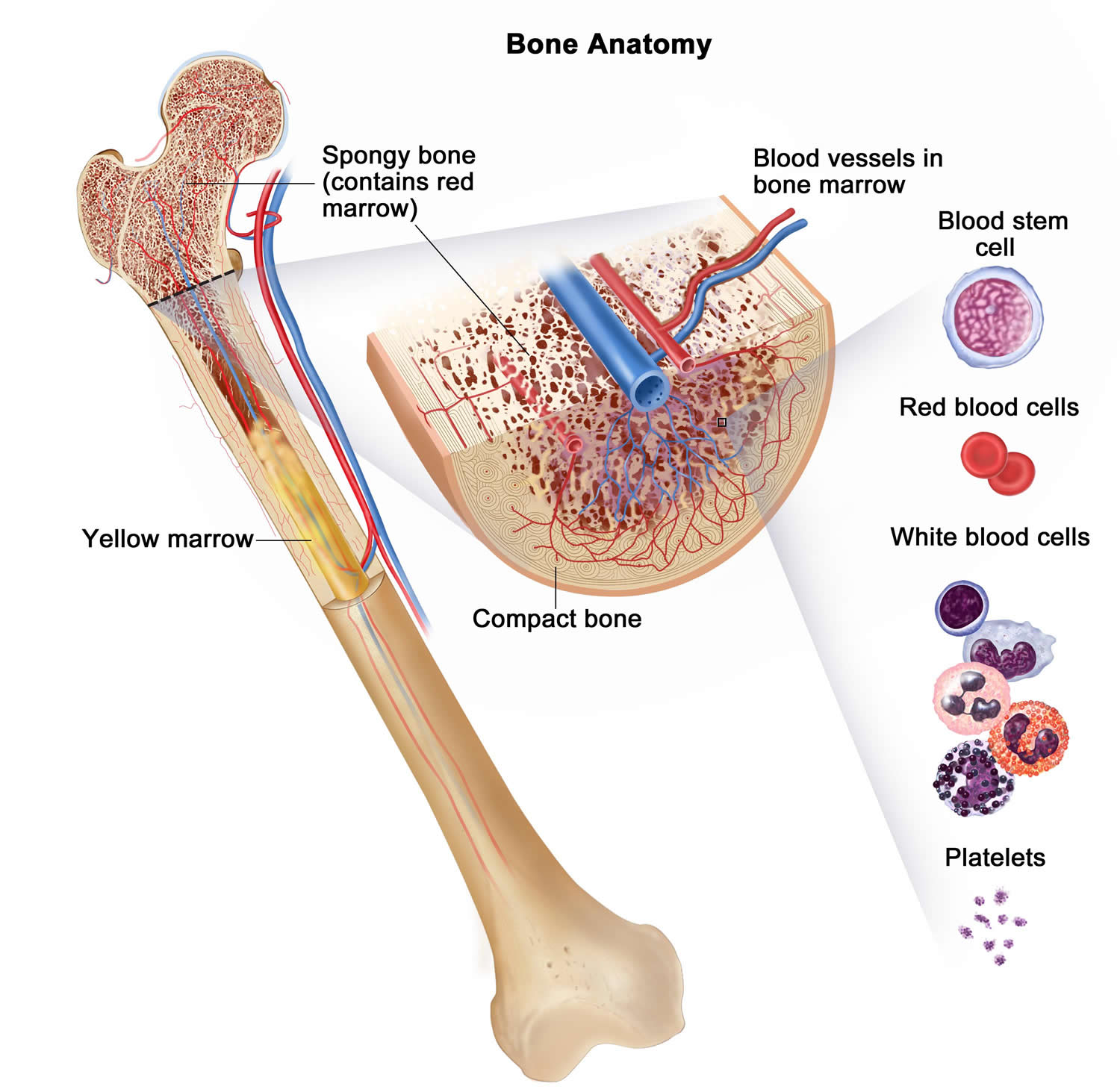
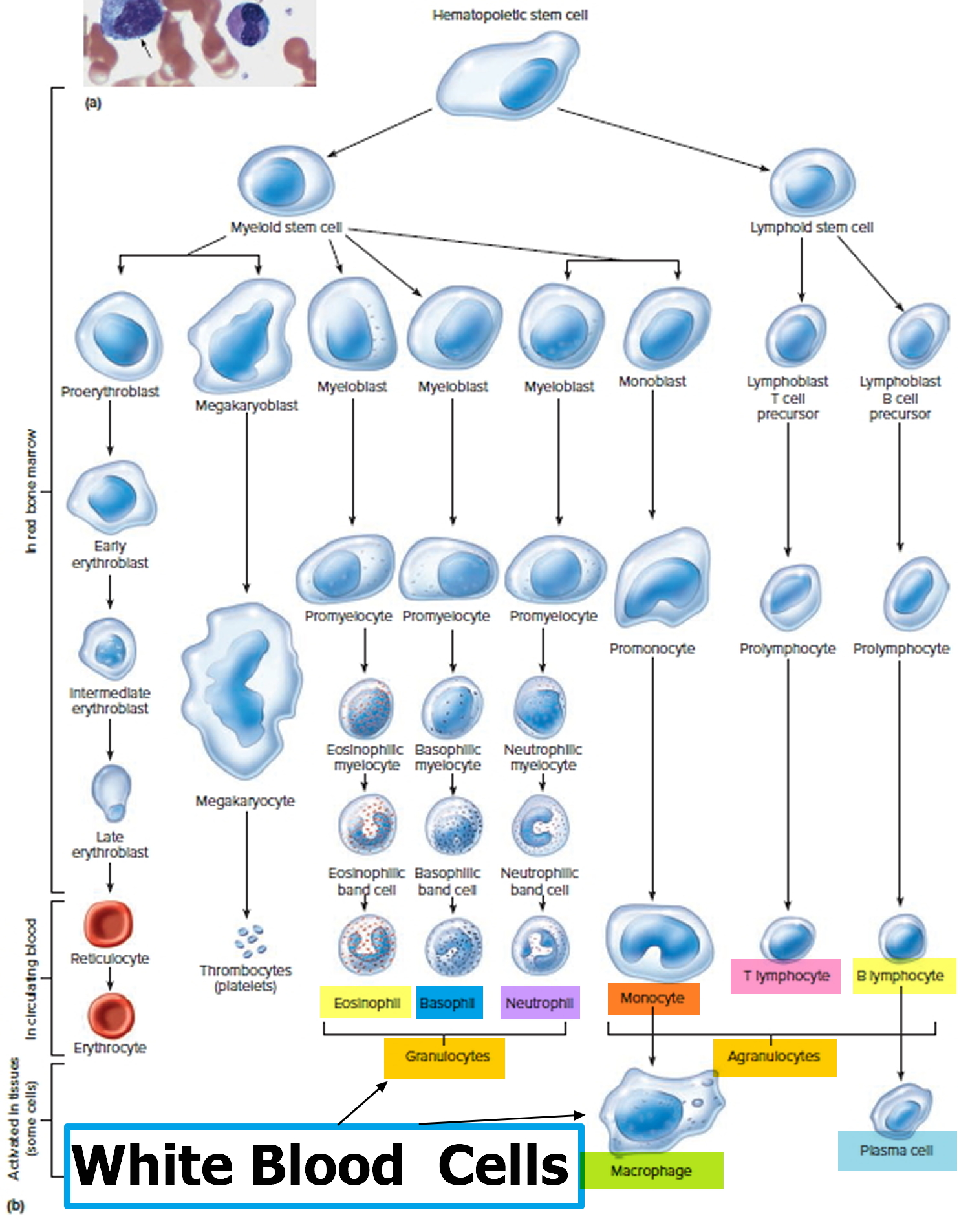
Figure 3. White blood cells development
A major breakthrough in the understanding of basophil effector functions came from studies using interleukin-4 (IL-4) -reporter animals, and led to the discovery that basophils are a primary source of IL-4 in vivo 14. It was therefore hypothesized that basophils direct the development of robust IL-4-producing CD4 T cells. It is well accepted that upon initial antigen encounter, naive CD4 T-cell differentiation into IL-4-producing T helper type 2 (Th2) effector cells occurs with the help of IL-4 present in the microenvironment 15. The initial source of IL-4 remained elusive for many years. However, unlike CD4 T cells, which must undergo rounds of differentiation to express significant IL-4 16, basophils immediately secrete IL-4 following activation, and this is readily achieved even in immature basophil precursor cells 17.
What do basophils do?
Basophils appear in many specific kinds of inflammatory reactions, particularly those that cause allergic symptoms. Basophils contain anticoagulant heparin, which prevents blood from clotting too quickly. They also contain the vasodilator histamine, which promotes blood flow to tissues. They can be found in unusually high numbers at sites of ectoparasite infection, e.g., ticks. Like eosinophils, basophils play a role in both parasitic infections and allergies 18. They are found in tissues where allergic reactions are occurring and probably contribute to the severity of these reactions. Basophils have protein receptors on their cell surface that bind IgE, an immunoglobulin involved in macroparasite defense and allergy. It is the bound IgE antibody that confers a selective response of these cells to environmental substances, for example, pollen proteins or helminth antigens. Recent studies in mice suggest that basophils may also regulate the behavior of T cells and mediate the magnitude of the secondary immune response 19.
Basophils in local acute allergic reactions
Local allergen challenge induces a prompt migration of basophils to the site of allergic inflammation.
Nose
Basophils have been identified in the nasal washes of patients with allergic rhinitis and are thought to be an important source of histamine in responses to allergen challenge 20. Braunstahl et al 21 demonstrated that segmental bronchoprovocation in non‐asthmatic allergic rhinitis patients affects mast cell and basophil numbers in nasal and bronchial mucosa. The number of basophils increased significantly after challenge, whereas the numbers of mast cells decreased, probably because of the limited immunohistochemical detection (by tryptase and chymase staining) of mast cells after degranulation. At the same time, this study 21 also demonstrated a decrease in the percentage of blood basophils, which might suggest an influx of basophils from the blood into the nasal and bronchial mucosa after the challenge. Interestingly, successful grass pollen immunotherapy is associated with inhibition of seasonal increases in basophils and eosinophils, but not mast cells or neutrophils, within the nasal epithelium of allergic rhinitis patients 22.
Skin
The skin might be an important route of allergen exposure, especially in the case of skin barrier disruption, and significant increases in the numbers of basophils were previously observed 6 hours after intradermal injection of allergen 23 or, in patch‐test skin sites, for house dust mite allergen 24. Furthermore, basophil infiltration into skin lesions seems to be more common than previously thought, indicating that they may play important roles in a variety of inflammatory skin diseases 25. Higher number of basophils were detected in inflammatory skin diseases where eosinophils are present 25 and those observations are consistent with a recent study which demonstrated a significant correlation between airway basophils and eosinophils in asthma patients 26.
Basophils in systemic allergic reactions
Human experimental models of acute allergic reactions
Currently, there are two models for studying anaphylaxis in humans: emergency department (ED) studies and controlled challenge models (mostly to food, but also to Hymenoptera sawflies, wasps, bees, and ants venom) 27. Smith et al 28 performed the first prospective human study during sting challenge‐induced systemic allergic reactions, which was followed by a series of similarly designed studies by van der Linden et al in the 1990s 29. However, for safety reasons, in controlled allergen challenge studies, patients with previous anaphylaxis are often excluded from challenge studies due to the potential for life‐threatening reactions 30. Furthermore, in the oral food challenge model, the reaction severity at challenge is also limited by the controlled nature of the challenge (allergen exposure is usually terminated at the onset of objective symptoms) and administration of pharmacologic interventions to treat the symptoms. Consequently, two studies which investigated the role of basophils in human anaphylaxis (after insect sting or food challenge), involved only a very limited number of patients who experienced severe reactions after challenge 31. However, studies in the challenge setting do have the advantage of allowing comparison with prereaction samples, optimal sampling and controlling potentially confounding factors (including acute treatment, where blood samples can often be taken prior to treatment) 32.
Allergen‐induced reactions often manifest themselves as an early asthmatic response, and bronchial allergen challenge may be another model for study of basophils during the acute allergic reaction 33. In addition, nasal allergen challenges could also be employed as an experimental set‐up to study the role of basophils in local allergic reactions 34.
The emergency department‐based anaphylaxis study was first described by Lin et al 35. Patients with anaphylaxis are studied prospectively at the time of presentation to the emergency department, with sample collection typically occurring 1 to 2 hours after onset of symptoms, and usually after initial treatment and stabilization. Patients with the most severe reactions including hypoxaemia or hypotension can be investigated, although this is typically after initial stabilization and treatment (usually with adrenaline). In the case of field‐treatment of anaphylaxis, patients are very often treated with systemic corticosteroids and antihistamines as well. Corticosteroids have broad immunological effects, albeit much delayed compared to other anti‐allergic therapies. With respect to basophils, corticosteroids inhibit their pro‐allergic functions 36 and this might be an important confounder.
The current evidence for basophil degranulation resulting in anaphylaxis in humans is very limited 37. However, there are several important indirect observations. Total tryptase (which is produced by mast cells, but not basophils) is within normal limits in up to 30% of patients with anaphylaxis 37. The proportion of patients with normal tryptase is even higher in the case of food‐induced anaphylaxis (even when blood samples are optimally timed) 38 or in the case of positive oral food challenge in which symptoms of anaphylaxis are observed 39. From these data, some authors speculate that, at least in some patients, the anaphylactic episode may primarily involve basophil and not mast cell degranulation 27. However, there are several other possible reasons for this discrepancy. For example, in the case of localized (e.g, in the gut) rather than generalized mast cell degranulation, tryptase may enter the circulation less efficiently. A further level of complexity is added by reports that some mast cells express less tryptase (i.e ,those present in the respiratory epithelium, alveolar wall and small intestinal mucosa) than others (eg in the skin, heart and perivascular tissue), and that in some subjects tryptase may be eliminated very rapidly 27.
Several studies have assessed the impact of the anti‐IgE monoclonal antibody omalizumab (which prevents IgE from binding to the high‐affinity IgE receptor) on the acute allergic response to nasal allergen or oral food challenge models, which has allowed an evaluation of the relative contribution of basophils and mast cells 40. Using titrated skin prick testing to assess mast cell responses, and histamine release assays after in vitro allergen stimulation to assess basophil responses, these studies demonstrated that a reduction in symptoms occurred when the basophil—rather than mast cell—response was reduced. These results indirectly suggest a potentially important role for basophils in acute allergic reactions. However, in vitro stimulation cannot directly show that basophils are involved in acute allergic response in different target organs.
Migration during anaphylaxis
The results of the experimental allergen challenge in the nose, airways and skin have demonstrated the influx of basophils to inflammatory sites several hours after allergen exposure 20. A recent study has suggested that basophils migrate from the circulation during anaphylaxis, both in emergency department and controlled allergen challenge models 32. In the emergency department study, which included predominantly venom‐allergic adult patients, there was a substantial reduction (80%) in circulating basophils during anaphylactic reactions, and these findings were replicated in peanut‐allergic individuals experiencing allergic reactions during double‐blind placebo‐controlled peanut challenge. In contrast to previous studies which monitored basophils at sites of allergen challenge 41, this study 32 assessed basophils in the peripheral blood, including absolute basophil count measured by flow cytometry using microbeads, with basophils identified as CD123+HLA‐DR‐ 80 or CRTh2+CD303‐CD123+ cells 42. Basophil migration was confirmed using whole blood gene expression analysis of genes which are specific for basophils, including the α‐subunit of the high‐affinity IgE receptor (FCER1A), carboxypeptidase A3 (CPA3) and histidine decarboxylase (HDC) 32. FcεRI is expressed on mast cells and basophils as tetramers (αβγ2) as well as on antigen‐presenting cells, although at substantially lower levels and only as trimmers (αγ2) 43. CPA3 is expressed in mast cells and basophils and may be expressed in populations of T cell progenitors and thymic T cells 44. HDC catalyses the formation of histamine from L‐histidine, and in hematopoietic cell lineages, the gene is expressed only in mast cells and basophils 45. Importantly, the expression of all three genes significantly decreased during anaphylaxis, and correlated with the absolute number of circulating basophils, indicating that the decrease in whole blood gene expression of FCER1A, CPA3 and HDC was due to reduced number of basophils in blood.
What is the clinical relevance of basophil migration?
The importance of basophil migration is currently unclear, and we do not know where or when basophil activation and degranulation occurs during anaphylaxis. The observations that IgE‐mediated basophil migration might occur without degranulation 46 and that migration out of the circulation may occur at the onset of symptoms 32, might be consistent with the hypothesis that basophils migrate to the site of allergen exposure where activation and degranulation could occur, thereby contributing to the clinical presentation (Figure 4). This is consistent with clinical observations of different severities and end‐organ patterns of anaphylaxis which suggest that local rather than generalized mast cell and/or basophil degranulation may predominate in some individuals 27.
Figure 4. Basophil migration in anaphylaxis
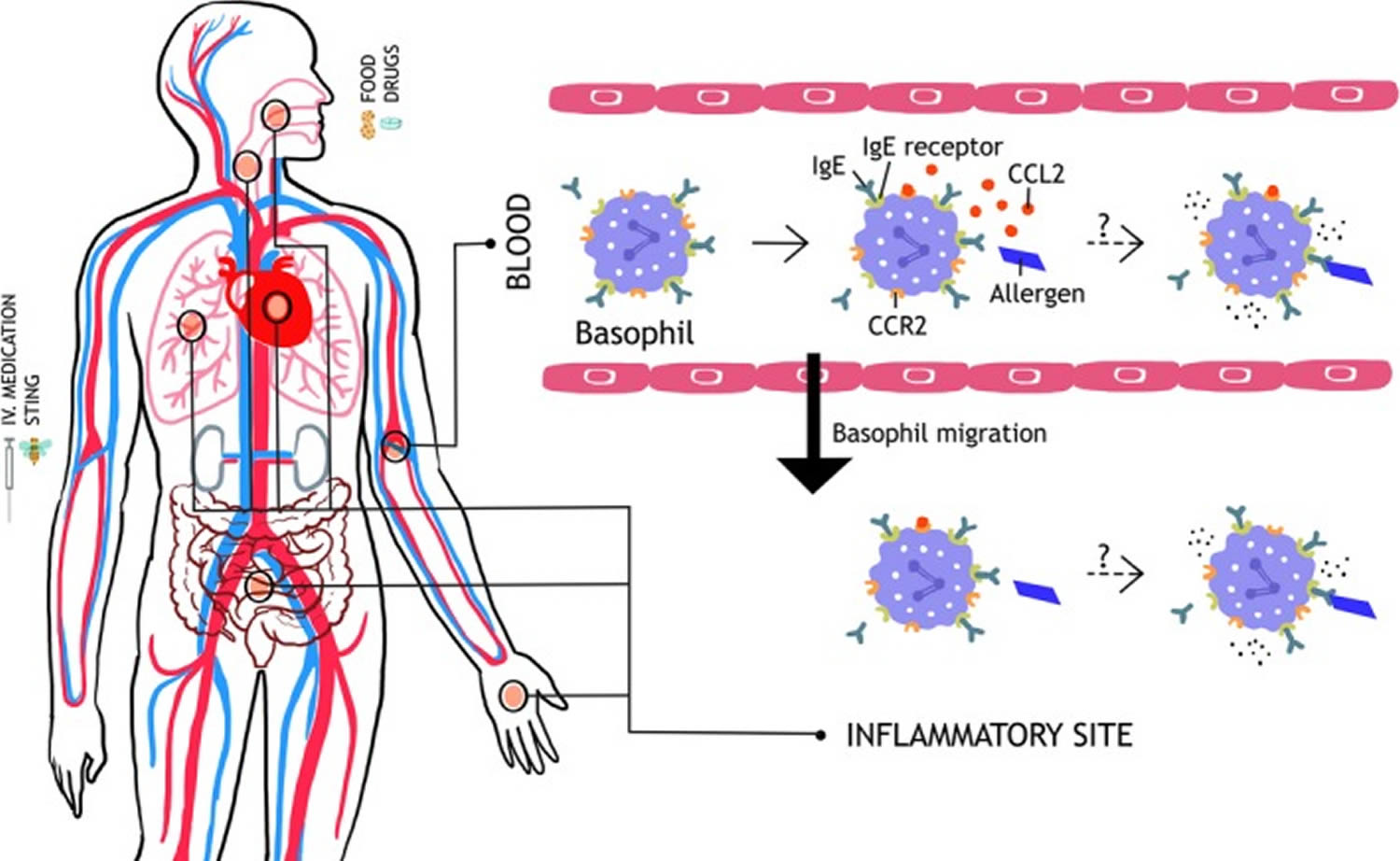
Footnotes: Hypothetical role of basophil migration in anaphylaxis. Upon allergen challenge, basophil‐directed chemokine CCL2 (possibly secreted from mast cells) induces a rapid migration of basophils out of the circulation. This may reactogenic, with migration to target organs resulting in activation and degranulation. Alternatively, the migration may be a protective response, removing basophils from the circulation so that they are unable to degranulate in response to circulating allergen.
Abbreviations: CCL2 = chemokine (C‐C motif) ligand 2; CCR2 = C‐C chemokine receptor type 2; IgE = Immunoglobulin E; IgE receptor, high‐affinity IgE Fc-epsilon RI receptor (FcεRI)
Basophils normal range
Basophils
Males:
- 0-14 days: 0.02-0.11 x 10(9)/L
- 15 days-7 weeks: 0.01-0.07 x 10(9)/L
- 8 weeks-35 months: 0.01-0.06 x 10(9)/L
- 3-17 years: 0.00-0.10 x 10(9)/L
- Adults: 0.01-0.08 x 10(9)/L
Females:
- 0-14 days: 0.02-0.07 x 10(9)/L
- 15 days-4 weeks: 0.01-0.06 x 10(9)/L
- 5 weeks-7 weeks: 0.01-0.05 x 10(9)/L
- 8 weeks-5 months: 0.01-0.07 x 10(9)/L
- 6 months-35 months: 0.01-0.06 x 10(9)/L
- 3-17 years: 0.00-0.10 x 10(9)/L
- Adults: 0.01-0.08 x 10(9)/L
Absolute basophils
Conventional Units
- Percent (mean) 0.3%
- Absolute count 0-200 (per microliter)
SI Units
- Mean number fraction 0.030
- Absolute count 0-0.20 X 109 per liter
Note: The reference ranges provided here represent a theoretical guideline that should not be used to interpret your test results. Some variation is likely between these numbers and the reference range reported by the lab that ran your test. Please consult your doctor for more information on reference ranges.
Reference ranges listed here are for adults older than 18 years old. They are not available for youths 0-18 years old due to wide variability.
Conventional Units are typically used for reporting results in U.S. labs. SI Units are used to report lab results outside of the U.S.
Many different conditions can result in increases or decreases in blood cell populations. Some of these conditions may require treatment, while others may resolve on their own.
Absolute basophil count
Basophils account for less than 3% of the circulating white blood cells (0 to 300 basophils per microliter of blood). The absolute basophil count (cells/L) can be determined by manual counting of peripheral blood smears or using cell-counting chambers as well as by automated hematology analyzers and fluorescence flow cytometry. Manual basophil counting of peripheral blood smears is currently regarded as the reference method, although the limitations of this method (distribution, observer, and statistical errors) are widely recognized. Automated hematology analyzers offer an advantage of larger numbers of counted cells and high throughput but are characterized by inconsistent analytical performance for basophil enumeration. Flow cytometric enumeration of circulating basophils using panels of monoclonal antibodies is being developed as novel candidate reference method for the absolute basophil count in peripheral blood. Basophil counting using fluorescence flow cytometry is characterized by high precision and statistical superiority.
Absolute basophils low
A decrease in the number of basophils (basopenia) can occur as a response to thyrotoxicosis, acute hypersensitivity reactions, and infections.
Absolute basophils high
An increase in the number of basophils (basophilia) can occur in people with hypothyroidism. In the myeloproliferative disorders (for example, polycythemia vera and myelofibrosis), a marked increase in the number of basophils can occur.
The symptoms usually depend on the disorder that is causing the change in the number of basophils, but increased numbers of basophils can cause itching and other allergic reactions.
The change in the number of basophils is usually detected by chance when a complete blood count is done for other reasons.
Basophils represent a part of malignant clone in primary myelofibrosis and are associated with unfavorable disease features and poor prognosis which is independent of currently established prognostic scoring system and monocytosis 47.
Treatment is directed at the disorder that is causing the change in the number of basophils.
- Basophil effector function and homeostasis during helminth infection. Blood 2009 113:2816-2825; doi: https://doi.org/10.1182/blood-2008-05-154773[↩]
- The 21st century renaissance of the basophil? Current insights into its role in allergic responses and innate immunity. Falcone FH, Zillikens D, Gibbs BF. Exp Dermatol. 2006 Nov; 15(11):855-64.[↩]
- Basophil effector function and homeostasis during helminth infection. Ohnmacht C, Voehringer D. Blood. 2009 Mar 19; 113(12):2816-25. http://www.bloodjournal.org/content/113/12/2816.long[↩]
- Min B, et al. Basophils produce IL-4 and accumulate in tissues after infection with a Th2-inducing parasite. J. Exp. Med. 2004;200:507–517. doi: 10.1084/jem.20040590.[↩]
- Stone KD, Prussin C, Metcalfe DD. IgE, mast cells, basophils, and eosinophils. J Allergy Clin. Immunol. 2010;125:S73–80. doi: 10.1016/j.jaci.2009.11.017.[↩]
- Yi M, Kim H-P, Jeong KY, Kim JY, Lee I-Y, Yong T-S. Effects of the Th2-dominant milieu on allergic responses in Der f 1-activated mouse basophils and mast cells. Scientific Reports. 2018;8:7706. doi:10.1038/s41598-018-25741-w. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5955989/[↩]
- Human basophils: mediator release and cytokine production. Schroeder JT, MacGlashan DW Jr, Lichtenstein LM. Adv Immunol. 2001; 77():93-122.[↩]
- IgE, mast cells, basophils, and eosinophils. Prussin C, Metcalfe DD. J Allergy Clin Immunol. 2003 Feb; 111(2 Suppl):S486-94.[↩]
- Roles of mast cells and basophils in innate and acquired immunity. Wedemeyer J, Tsai M, Galli SJ. Curr Opin Immunol. 2000 Dec; 12(6):624-31.[↩]
- IL-3 is required for increases in blood basophils in nematode infection in mice and can enhance IgE-dependent IL-4 production by basophils in vitro. Lantz CS, Min B, Tsai M, Chatterjea D, Dranoff G, Galli SJ. Lab Invest. 2008 Nov; 88(11):1134-42.[↩][↩]
- Purified human peripheral blood basophils release interleukin-13 and preformed interleukin-4 following immunological activation. Gibbs BF, Haas H, Falcone FH, Albrecht C, Vollrath IB, Noll T, Wolff HH, Amon U. Eur J Immunol. 1996 Oct; 26(10):2493-8.[↩]
- Min B, Prout M, Hu-Li J, et al. Basophils produce IL-4 and accumulate in tissues after infection with a Th2-inducing parasite. J Exp Med. 2004;200:507–17.[↩]
- Min B, Brown MA, LeGros G. Understanding the roles of basophils: breaking dawn. Immunology. 2012;135(3):192-197. doi:10.1111/j.1365-2567.2011.03530.x. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3311041/[↩]
- Type 2 immunity reflects orchestrated recruitment of cells committed to IL-4 production. Voehringer D, Shinkai K, Locksley RM. Immunity. 2004 Mar; 20(3):267-77.[↩]
- Differentiation of effector CD4 T cell populations (*). Zhu J, Yamane H, Paul WE. Annu Rev Immunol. 2010; 28():445-89.[↩]
- Helper T cell differentiation is controlled by the cell cycle. Bird JJ, Brown DR, Mullen AC, Moskowitz NH, Mahowald MA, Sider JR, Gajewski TF, Wang CR, Reiner SL. Immunity. 1998 Aug; 9(2):229-37.[↩]
- IL-3 promotes production of IL-4 by splenic non-B, non-T cells in response to Fc receptor cross-linkage. Le Gros G, Ben-Sasson SZ, Conrad DH, Clark-Lewis I, Finkelman FD, Plaut M, Paul WE. J Immunol. 1990 Oct 15; 145(8):2500-6.[↩]
- The role of basophils in helminth infection. Trends Parasitol. 2009 Dec;25(12):551-6. doi: 10.1016/j.pt.2009.09.004. Epub 2009 Sep 24. https://www.ncbi.nlm.nih.gov/pubmed/19782643[↩]
- Basophils as APC in Th2 response in allergic inflammation and parasite infection. Curr Opin Immunol. 2010 Dec;22(6):814-20. doi: 10.1016/j.coi.2010.10.018. Epub 2010 Nov 20. https://www.ncbi.nlm.nih.gov/pubmed/21095110[↩]
- Iliopoulos O, Baroody FM, Naclerio RM, Bochner BS, Kagey‐Sobotka A, Lichtenstein LM. Histamine‐containing cells obtained from the nose hours after antigen challenge have functional and phenotypic characteristics of basophils. J Immunol. 1992;148:2223‐2228.[↩][↩]
- Braunstahl GJ, Overbeek SE, Fokkens WJ, et al. Segmental bronchoprovocation in allergic rhinitis patients affects mast cell and basophil numbers in nasal and bronchial mucosa. Am J Respir Crit Care Med. 2001;164:858‐865[↩][↩]
- Wilson DR, Irani AM, Walker SM, et al. Grass pollen immunotherapy inhibits seasonal increases in basophils and eosinophils in the nasal epithelium. Clin Exp Allergy. 2001;31:1705‐1713.[↩]
- Irani AM, Huang C, Xia HZ, et al. Immunohistochemical detection of human basophils in late‐phase skin reactions. J Allergy Clin Immunol. 1998;101:354‐362[↩]
- Mitchell EB, Chapman M, Pope FM, Crow J, Jouhal S, Platts‐Mills TE. Basophils in allergen‐induced patch test sites in atopic dermatitis. Lancet. 1982;319:127‐130.[↩]
- Ito Y, Satoh T, Takayama K, Miyagishi C, Walls AF, Yokozeki H. Basophil recruitment and activation in inflammatory skin diseases. Allergy. 2011;66:1107‐1113.[↩][↩]
- Suzuki Y, Wakahara K, Nishio T, Ito S, Hasegawa Y. Airway basophils are increased and activated in eosinophilic asthma. Allergy. 2017;72:1532‐1539.[↩]
- Simons FER, Frew AJ, Ansotegui IJ, et al. Risk assessment in anaphylaxis: current and future approaches. J Allergy Clin Immunol. 2007;1(Suppl):S2‐S24.[↩][↩][↩][↩]
- Smith PL, Kagey‐Sobotka A, Bleecker ER, et al. Physiologic manifestations of human anaphylaxis. J Clin Invest. 1980;66:1072‐1080.[↩]
- van der Linden PW, Hack CE, Struyvenberg A, et al. Controlled insect‐sting challenge in 55 patients: correlation between activation of plasminogen and the development of anaphylactic shock. Blood. 1993;82:1740‐1748.[↩]
- Kowalski ML, Ansotegui I, Aberer W, et al. Risk and safety requirements for diagnostic and therapeutic procedures in allergology: World Allergy Organization Statement. World Allergy Organ J. 2016;9:33[↩]
- Commins SP, James HR, Stevens W, et al. Delayed clinical and ex vivo response to mammalian meat in patients with IgE to galactose‐alpha‐1,3‐galactose. J Allergy Clin Immunol. 2014;134:108‐115[↩]
- Korosec P, Turner PJ, Silar M, et al. Basophils, high‐affinity IgE receptors and CCL2 in human anaphylaxis. J Allergy Clin Immunol. 2017;140:750‐758.e15.[↩][↩][↩][↩][↩]
- Gauvreau GM, Lee JM, Watson RM, Irani AM, Schwartz LB, O’Byrne PM. Increased numbers of both airway basophils and mast cells in sputum after allergen inhalation challenge of atopic asthmatics. Am J Respir Crit Care Med. 2000;161:1473‐1478.[↩]
- Eckman JA, Sterba PM, Kelly D, et al. Effects of omalizumab on basophil and mast cell responses using an intranasal cat allergen challenge. J Allergy Clin Immunol. 2010;125:889‐895.e7.[↩]
- Lin RY, Schwartz LB, Curry A, et al. Histamine and tryptase levels in patients with acute allergic reactions: an emergency department‐based study. J Allergy Clin Immunol. 2000;106(1 Pt 1):65‐71.[↩]
- Saavedra‐Delgado AM, Mathews KP, Pan PM, Kay DR, Muilenberg ML. Dose‐response studies of the suppression of whole blood histamine and basophil counts by prednisone. J Allergy Clin Immunol. 1980;66:464‐471[↩]
- Korošec P, Gibbs BF, Rijavec M, Custovic A, Turner PJ. Important and specific role for basophils in acute allergic reactions. Clinical and Experimental Allergy. 2018;48(5):502-512. doi:10.1111/cea.13117. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5947573/[↩][↩]
- Brown SGA, Stone SF, Fatovich DM, et al. Anaphylaxis: clinical patterns, mediator release, and severity. J Allergy Clin Immunol. 2013;132:1141‐1149.e5.[↩]
- Sampson HA, Mendelson L, Rosen JP. Fatal and near‐fatal anaphylactic reactions to food in children and adolescents. N Engl J Med. 1992;327:380‐384.[↩]
- Savage JH, Courneya J‐P, Sterba PM, MacGlashan DW, Saini SS, Wood RA. Kinetics of mast cell, basophil, and oral food challenge responses in omalizumab‐treated adults with peanut allergy. J Allergy Clin Immunol. 2012;130:1123‐1129.[↩]
- Iliopoulos O, Baroody FM, Naclerio RM, Bochner BS, Kagey‐Sobotka A, Lichtenstein LM. Histamine‐containing cells obtained from the nose hours after antigen challenge have functional and phenotypic characteristics of basophils. J Immunol. 1992;148:2223‐2228[↩]
- Shamji MH, Bellido V, Scadding GW, et al. Effector cell signature in peripheral blood following nasal allergen challenge in grass pollen allergic individuals. Allergy Eur J Allergy Clin Immunol. 2015;70:171‐179.[↩]
- Stone KD, Prussin C, Metcalfe DD. IgE, mast cells, basophils, and eosinophils. J Allergy Clin Immunol. 2010;125(2 Suppl. 2):S73‐S80[↩]
- Lilla JN, Chen CG, Mukai K, et al. Reduced mast cell and basophil numbers and function in Cpa3‐Cre; Mcl‐1 fl/fl mice. Blood. 2011;118:6930‐6938[↩]
- Kuramasu A, Saito H, Suzuki S, Watanabe T, Ohtsu H. Mast cell‐/basophil‐specific transcriptional regulation of human L‐histidine decarboxylase gene by CpG methylation in the promoter region. J Biol Chem. 1998;273:31607‐31614[↩]
- Suzukawa M, Hirai K, Iikura M, et al. IgE‐ and Fc RI‐mediated migration of human basophils. Int Immunol. 2005;17:1249‐1255.[↩]
- High absolute basophil count is a powerful independent predictor of inferior overall survival in patients with primary myelofibrosis. Hematology. 2018 May;23(4):201-207. doi: 10.1080/10245332.2017.1376843. Epub 2017 Sep 14. https://www.ncbi.nlm.nih.gov/pubmed/28906207[↩]


{kind=link}