Contents
What is leucine
Leucine (C6H13O2N) is an essential branched-chain amino acid (BCAA) that is used in the biosynthesis of proteins, many metabolic functions and important for hemoglobin formation. Leucine is one of nine essential amino acids in humans meaning your body cannot synthesize it, leucine must be obtained from food and leucine is also used as a food additive (E641) as a flavor enhancer. Human dietary sources of leucine are foods that contain protein, such as meats, dairy products, soy products, beans and legumes and leucine deficiency is rare. Leucine contributes to regulation of blood-sugar levels; growth and repair of muscle and bone tissue; growth hormone production; and wound healing. Leucine is available to skeletal muscle where it functions as a nutrient signal, is used for protein synthesis, and serves as a metabolic fuel and/or a nitrogen donor for the synthesis of glutamine and alanine 1. Leucine also prevents breakdown of muscle proteins after trauma or severe stress and may be beneficial for individuals with phenylketonuria. Additionally, similarly to other amino acids, the carbon skeleton of leucine can be used to generate ATP 2. Therefore, leucine supplementation has been studied in a variety of conditions such as aging, muscle lesions, protein/energy deprivation, obesity, and diabetes mellitus. Because leucine availability influences signaling pathways involved in the regulation of metabolism and because the incidence of metabolic diseases has reached alarming levels worldwide, investigating nutritional supplements that are potentially beneficial for the treatment and prevention of obesity and diabetes mellitus has become of paramount importance. However, base on the available evidence, oral leucine supplementation produces no or very mild effects on food intake 2. Furthermore, despite the well-known stimulatory effects of leucine on protein synthesis, leucine supplementation in aging rats or humans 3, 4 apparently did not lead to an increase in lean and protein body mass. Interestingly, several studies found no effects on adiposity 5 or even a predisposition to accumulate more body fat 6 in animals that began to receive leucine supplementation when they were already obese. A likely explanation for these findings is that mTOR complex 1 (mTORC1) activation may promote fat storage in adipocytes by suppressing lipolysis and stimulating de novo lipogenesis 7. Additionally, mice with disrupted mTORC1 complex exhibit less adipose tissue, suggesting that fat deposition may depend on mTORC1 activity 8. Branched-chain amino acid (BCAA) supplementation in pregnant rats consuming a protein-restricted diet restored the fat mass of their offspring to levels similar to those of non-restricted animals 9. Studies that initiated leucine supplementation in already-obese animals found no beneficial effects of leucine or even found worsening of the degree of adiposity 2. Therefore, based on the presented data, leucine supplementation is not likely to be helpful as a dietary supplement for treating obesity 2.
The initiation of mRNA translation is the major mechanism by which leucine stimulates protein synthesis. Classical studies have shown that the regulation of mRNA translation by leucine is dependent on the mammalian target of rapamycin (mTOR) because rapamycin, a specific mTOR inhibitor, is able to blunt the effects of leucine 10. Mammalian target of rapamycin (mTOR) is a serine/threonine kinase that is involved in the regulation of multiple cellular processes, including protein synthesis and cell growth, proliferation, and survival. Mammalian target of rapamycin (mTOR) controls protein synthesis through mTOR complex 1 (mTORC1), which comprises mTOR itself and other proteins.
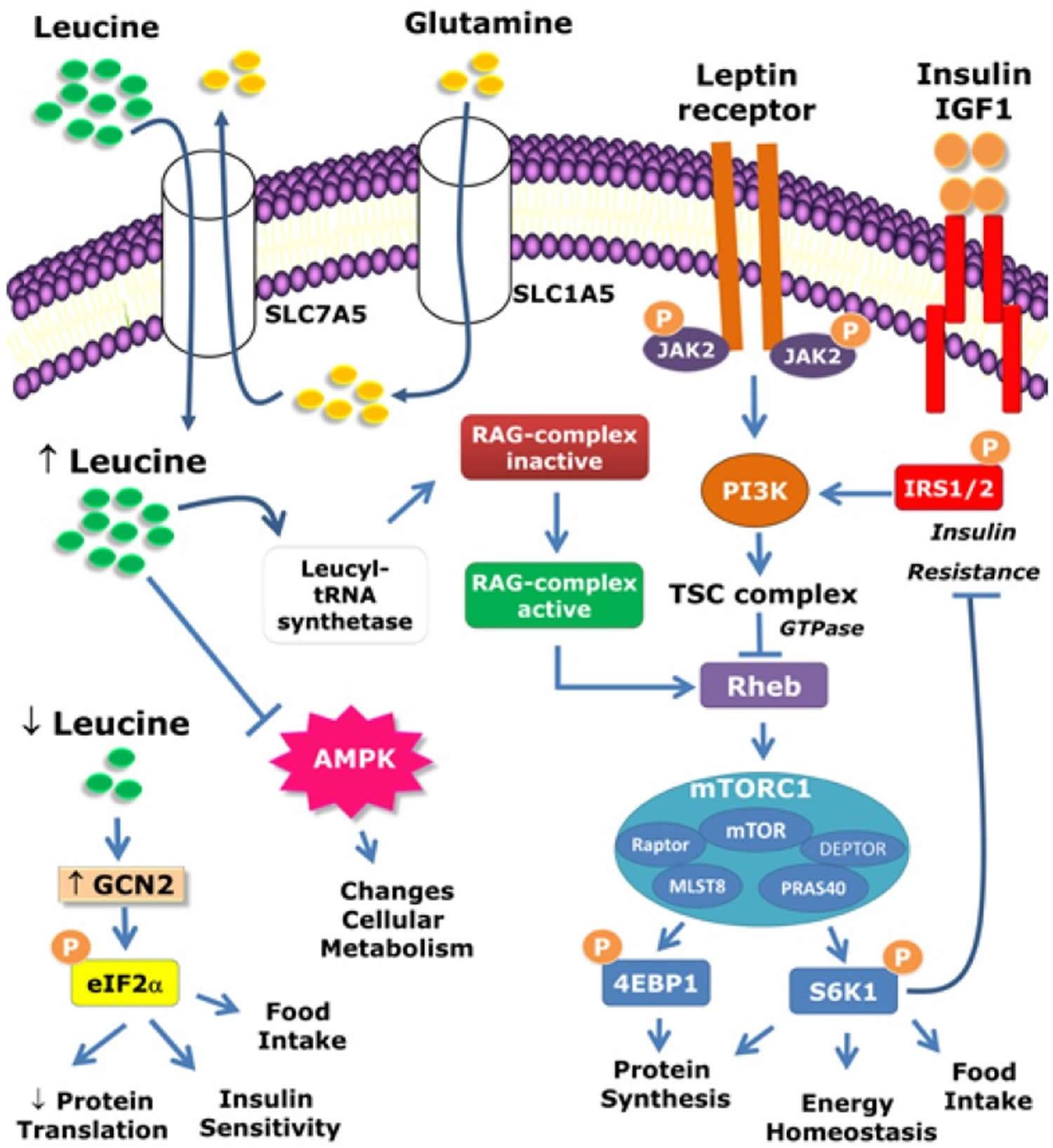
Figure 1. Intracellular mechanisms activated by leucine
Footnotes:
The mammalian target of rapamycin complex 1 (mTORC1) comprises mTOR, Raptor, mLST8, PRAS40, and DEPTOR. mTORC1 is activated by amino acids (especially leucine) as well as by hormones such as leptin, insulin, and IGF-1. mTORC1 can be activated by different pathways. Hormonal activation primarily occurs through the TSC complex. However, amino acid-dependent mTORC1 activation occurs through the Rag complex. The leucyl-tRNA synthetase is responsible for sensing leucine cellular levels and activating the Rag complex. The cellular uptake of L-glutamine and its subsequent rapid efflux in the presence of leucine represent the rate-limiting step of mTOR activation. The protein p70-S6 kinase 1 (S6K1) and eukaryotic initiation factor 4E (eIF4E) binding protein 1 (4E-BP1) are key downstream targets of mTORC1. S6K1 also phosphorylates components of the insulin signaling pathway, which may lead to insulin resistance in situations of nutrient abundance such as in obesity. The anorexigenic effects of leptin require both the phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3K) and mTOR/S6K1 signaling pathways. Because mTOR is a downstream target of PI3K signaling, the acute anorexigenic effects of leptin may depend on the PI3K/mTOR/S6K1 pathway.
[Source 2]How much leucine per day?
42 mg/kg body weight/day 11
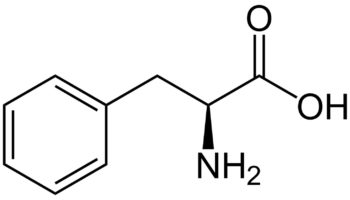
What is L-leucine
L-leucine, also known as 2-amino-4-methylvaleric acid, belongs to leucine and derivatives class of compounds. Those are compounds containing leucine or a derivative thereof resulting from reaction of leucine at the amino group or the carboxy group, or from the replacement of any hydrogen of glycine by a heteroatom. L-leucine is soluble (in water) and a moderately acidic compound. L-leucine can be found primarily in most biofluids, including blood, cerebrospinal fluid (CSF), feces, and sweat, as well as throughout most human tissues. Within the cell, L-leucine is primarily located in the mitochondria. It can also be found in the extracellular space. L-leucine exists in all living species, ranging from bacteria to humans.
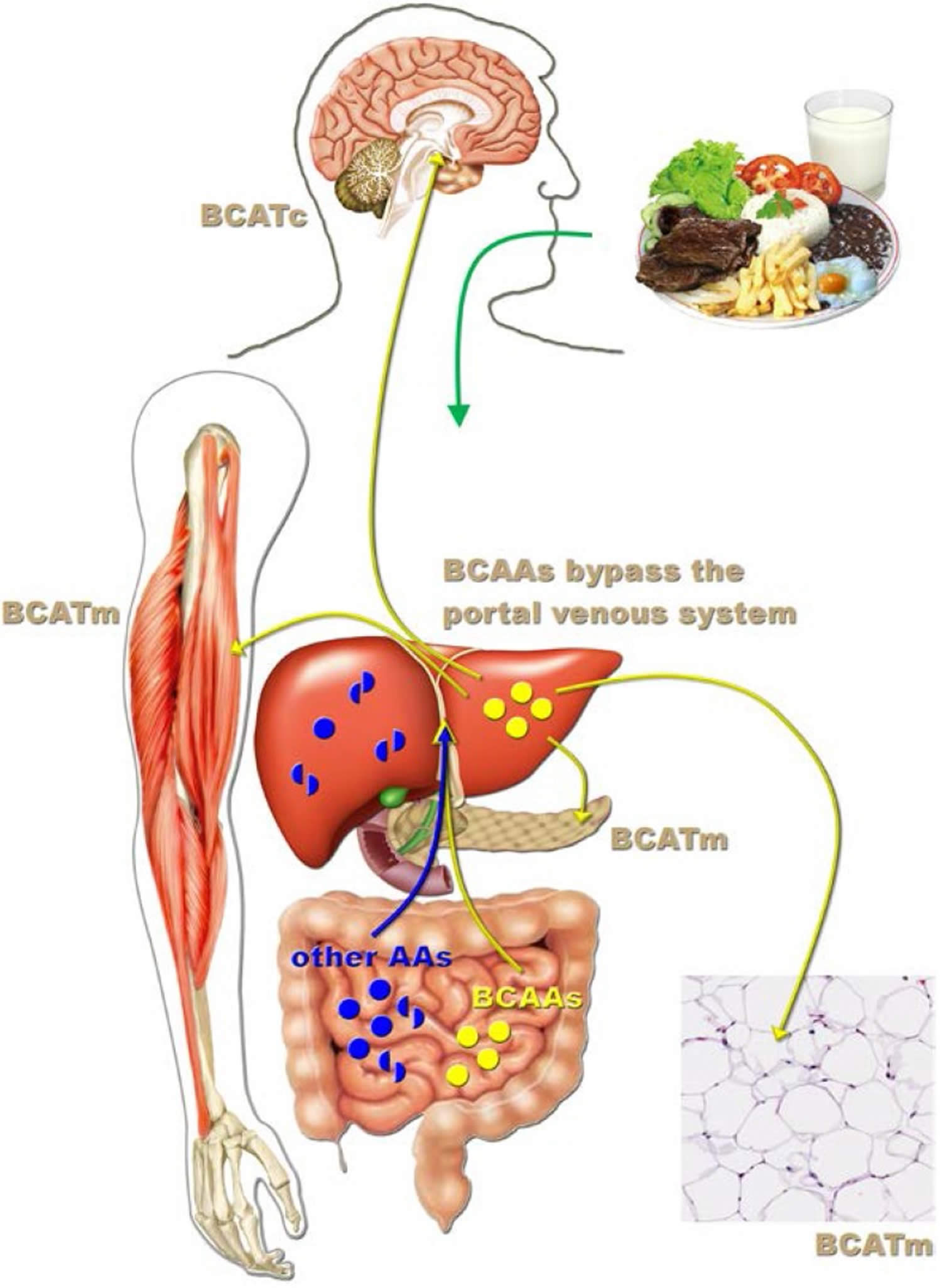
Figure 2. Leucine-responsive tissues
Footnotes:
After protein-rich meals, circulating branched-chain amino acid (BCAA) levels significantly increase, whereas other amino acids are highly metabolized by the gut or liver before reaching the systemic circulation. Branched-chain amino acid transaminase (BCAT) catalyzes the first and reversible transamination step of leucine degradation. This enzyme is not expressed in the liver, which allows the BCAAs to bypass the portal venous system following their intestinal absorption. In the brain, leucine is metabolized by the cytosolic form of BCAT (BCATc), whereas in other tissues (e.g., white adipose tissue, skeletal muscle, and pancreas), the mitochondrial form of BCAT (BCATm) prevails.
[Source 2]Does leucine work for weight loss?
Several studies found that central leucine infusion (infusion of leucine into the brain) reduces food intake in rodents 12. However, the capacity of leucine to modulate food intake is controversial 2. Many supplementation approaches have been used to study the effects of leucine on food intake, including leucine supplementation in the drinking water, in the diet and through gavage, as well as by subcutaneous (sc), intraperitoneal (ip), and central injections 2. To clarify whether leucine supplementation is able to influence feeding behavior, the studies evaluating the effects of central leucine infusion observed decreased food intake 2. These findings demonstrated that leucine could inhibit food intake by directly affecting the CNS (central nervous system) (see Figure 3). This result is interesting because it is well known that the brain is able to sense changing nutrient levels to regulate the energy balance 13. However, of 30 studies investigating the effects of leucine supplementation in the diet, two studies found increased food intake in leucine-treated animals 14, 15, and only four studies reported decreased food intake in the leucine-supplemented groups 16, 17, 18, 12. From these studies, two observed increased taste aversion to the leucine-rich diet, which may explain the reduction in food intake 16, 12. To avoid possible aversive behavior in relation to the diet, several studies supplemented leucine in the drinking water. Of 13 studies, two observed that leucine supplementation decreased food intake in mice in specific conditions 5, 19. For example, leucine supplementation in the drinking water decreased the food intake of a polygenic model predisposed to type 2 diabetes (RCS10 mice) but did not affect the food intake of a monogenic model predisposed to obesity and severe insulin resistance (yellow agouti mice) 5. In another study, leucine supplementation in the drinking water decreased the food intake in males but not in females consuming a regular rodent diet. No effect was observed in mice consuming an high fat diet 19. Additionally, one study reported increased food intake in animals supplemented with leucine in the drinking water 20. No changes in food intake were observed in mice that received leucine supplementation through gavage, intraperitoneal or subcutaneous injections. Thus, the central anorexigenic effect of leucine is not well repeated when leucine supplementation is provided through an oral form (Figure 3).
The divergent results caused by oral or central leucine supplementation may be explained by the capacity of leucine to cross the blood-brain barrier (BBB) and reach the CNS. Although a previous study demonstrated that a 4% leucine-enriched meal could increase the leucine concentration in the cerebrospinal fluid to 44% 13, whether this change is robust enough to reduce food intake or to persist in the long term remains unknown. In this same study, a high-protein diet did not significantly increase the leucine concentration in the cerebrospinal fluid (CSF). Additionally, the changes in leucine levels were approximately seven-fold higher in the plasma than in the cerebrospinal fluid 13. Therefore, additional studies are warranted to establish the minimum increase in central leucine levels to significantly affect food intake. The blood-brain barrier (BBB) and glial cells maintain amino acid concentrations in the CNS parenchyma at well-controlled levels. This is important because several ubiquitous neurotransmitters are amino acids (i.e., glutamate and glycine) or molecules derived from amino acids (i.e., GABA, dopamine, noradrenaline, serotonin, and histamine). Earlier studies have suggested that leucine availability affects the synthesis of amino acid neurotransmitters such as glutamate 21. Abrupt changes in amino acid levels in the brain can cause cell death and serious neuronal dysfunction. For example, excessive activation of the N-methyl-D-aspartate (NMDA) receptor, which is a glutamate receptor, causes neuronal death 22. Similarly, treatment with monosodium glutamate (MSG) at a young age, when the blood-brain barrier (BBB) is not completely formed, causes lesions in several brain areas 23. Therefore, the ingestion of specific amino acids does not necessarily lead to pronounced changes in their levels in the brain. Likewise, the direct infusion of amino acids into the brain does not necessarily reproduce the physiological effects caused by oral supplementation and therefore may produce supraphysiological effects. Even brain nuclei that are located near circumventricular organs (CVOs) are not free from the blood-brain barrier (BBB) influence. For example, the arcuate nucleus of the hypothalamus (ARH) is close to the median eminence (ME), and the nucleus of the solitary tract (NTS) is close to the area postrema (AP) (Figure 3). Blood vessels in these areas allow nutrients and hormones to more easily gain access to the CNS. Nonetheless, the BBB and glial cells in these areas remain capable of controlling abrupt changes in the concentration of molecules. Therefore, the available evidence suggests that oral leucine supplementation produces no or very mild effects on food intake (Figure 3). Because previous studies have suggested a role of leucine supplementation in the treatment of obesity 24, the practical implication is that if leucine regulates the energy balance and favors a reduced adiposity, this effect is likely not mediated by changes in food intake.
Furthermore, several studies found leucine supplementation had no effects on adiposity 5 or even a predisposition to accumulate more body fat 6 in animals that began to receive leucine supplementation when they were already obese. A likely explanation for these findings is that mTOR complex 1 (mTORC1) activation may promote fat storage in adipocytes by suppressing lipolysis and stimulating de novo lipogenesis 7. Additionally, mice with disrupted mTORC1 complex exhibit less adipose tissue, suggesting that fat deposition may depend on mTORC1 activity 8. Branched-chain amino acid (BCAA) supplementation in pregnant rats consuming a protein-restricted diet restored the fat mass of their offspring to levels similar to those of non-restricted animals 9.
Therefore, based on the presented data, leucine supplementation is not likely to be helpful as a dietary supplement for treating obesity 2.
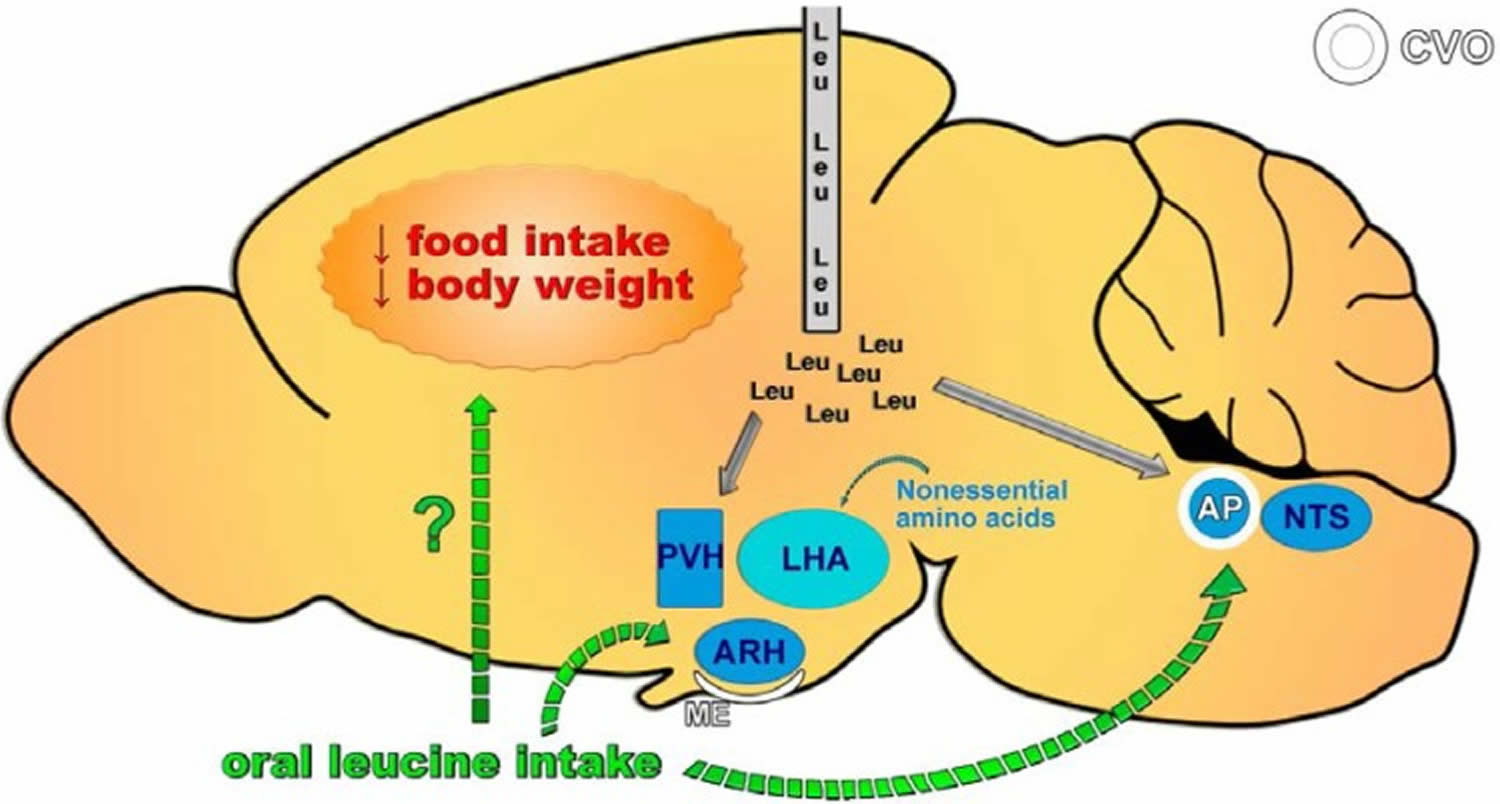
Figure 3. Brain effects of leucine on feeding
Footnotes:
Neuronal circuitries required for the central effects of leucine on feeding. Central leucine administration (intracerebroventricular or parenchymal) acutely decreases food intake and body weight. This response is due to the activation of hypothalamic nuclei involved in regulating energy balance, including the paraventricular nucleus of the hypothalamus (PVH) and the arcuate nucleus of the hypothalamus (ARH), as well as extra-hypothalamic sites such as the nucleus of the solitary tract (NTS). Conversely, oral leucine administration does not induce neuronal activation in the PVH, ARH, or NTS but does cause c-Fos expression in the area postrema (AP). Consequently, no robust evidence indicates that oral leucine intake affects food intake. CVO = circumventricular organ; ME = median eminence.
[Source 2]Leucine supplement side effects
Leucine toxicity, as seen in decompensated maple syrup urine disease, causes delirium and neurologic compromise, and can be life-threatening.
A high intake of leucine may cause or exacerbate symptoms of pellagra in people with low niacin status because it interferes with the conversion of L-tryptophan to niacin 25.
Leucine at a dose exceeding 500 mg/kg/d was observed with hyperammonemia 26. On the basis of plasma and urinary variables, the tolerable upper intake level (UL) for leucine in healthy adult men can be suggested at 500 mg/kg body weight per day or 35 g/day as a cautious estimate under acute dietary conditions 26.
Leucine rich foods
Table 1. Foods high in leucine
| Description | Leucine (g) Value Per 100 gram |
| Egg, white, dried, stabilized, glucose reduced | 7.36 |
| Egg, white, dried, powder, stabilized, glucose reduced | 7.17 |
| Egg, white, dried | 6.84 |
| Soy protein isolate | 6.78 |
| Soy protein isolate, potassium type | 6.78 |
| Egg, white, dried, flakes, stabilized, glucose reduced | 6.7 |
| Whale, beluga, meat, dried (Alaska Native) | 5.78 |
| Fish, cod, Atlantic, dried and salted | 5.11 |
| Seaweed, spirulina, dried | 4.95 |
| Soy protein concentrate, produced by alcohol extraction | 4.92 |
| Soy protein concentrate, produced by acid wash | 4.92 |
| Beverages, Protein powder soy based | 4.6 |
| Egg, whole, dried, stabilized, glucose reduced | 4.23 |
| Egg, whole, dried | 4.15 |
| Cheese, parmesan, shredded | 4.01 |
| Seeds, sesame flour, low-fat | 3.84 |
| Soy flour, defatted | 3.83 |
| Mollusks, whelk, unspecified, cooked, moist heat | 3.81 |
| Soy meal, defatted, raw | 3.66 |
| Tofu, dried-frozen (koyadofu) | 3.64 |
| Tofu, dried-frozen (koyadofu), prepared with calcium sulfate | 3.64 |
| Milk, dry, nonfat, regular, without added vitamin A and vitamin D | 3.54 |
| Milk, dry, nonfat, regular, with added vitamin A and vitamin D | 3.54 |
| Seeds, sunflower seed flour, partially defatted | 3.5 |
| Milk, dry, nonfat, calcium reduced | 3.48 |
| Cheese, parmesan, hard | 3.45 |
| Milk, dry, nonfat, instant, with added vitamin A and vitamin D | 3.44 |
| Milk, dry, nonfat, instant, without added vitamin A and vitamin D | 3.44 |
| Seeds, cottonseed flour, low fat (glandless) | 3.4 |
| Peanut flour, defatted | 3.38 |
| Milk, buttermilk, dried | 3.36 |
| Seeds, cottonseed meal, partially defatted (glandless) | 3.35 |
| Snacks, pork skins, plain | 3.32 |
| Soybeans, mature seeds, raw | 3.31 |
| Meat extender | 3.26 |
| Soybeans, mature seeds, dry roasted | 3.22 |
| Snacks, pork skins, barbecue-flavor | 3.22 |
| Beef, round, top round roast, boneless, separable lean only, trimmed to 0″ fat, select, cooked, roasted | 3.22 |
| Pork, cured, bacon, cooked, microwaved | 3.21 |
| Beef, round, eye of round roast, boneless, separable lean only, trimmed to 0″ fat, select, cooked, roasted | 3.19 |
| Beef, loin, top sirloin filet, boneless, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 3.17 |
| Beef, plate steak, boneless, inside skirt, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 3.15 |
| Cheese, gruyere | 3.1 |
| Seeds, sesame flour, partially defatted | 3.09 |
| Beef, round, top round steak, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, grilled | 3.08 |
| Cheese, romano | 3.07 |
| Beef, top loin filet, boneless, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 3.07 |
| Beef, loin, top loin steak, boneless, lip-on, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 3.05 |
| Beef, loin, top loin steak, boneless, lip off, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 3.04 |
| Beef, round, eye of round steak, boneless, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 3.04 |
| Beef, loin, top sirloin petite roast, boneless, separable lean only, trimmed to 0″ fat, select, cooked, roasted | 3.02 |
| Beef, rib, back ribs, bone-in, separable lean only, trimmed to 0″ fat, select, cooked, braised | 3.01 |
| Cheese, parmesan, dry grated, reduced fat | 2.98 |
| Beef, round, top round steak, boneless, separable lean only, trimmed to 0″ fat, choice, cooked, grilled | 2.98 |
| Beef, rib eye steak, boneless, lip-on, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 2.98 |
| Beef, rib eye steak, boneless, lip off, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 2.98 |
| Cheese, swiss | 2.96 |
| Beef, rib eye roast, boneless, lip-on, separable lean only, trimmed to 1/8″ fat, select, cooked, roasted | 2.95 |
| Beef, plate steak, boneless, outside skirt, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 2.94 |
| Beef, top loin petite roast, boneless, separable lean only, trimmed to 1/8″ fat, select, cooked, roasted | 2.93 |
| Veal, leg (top round), separable lean only, cooked, braised | 2.92 |
| Leavening agents, yeast, baker’s, active dry | 2.92 |
| Beef, round, top round roast, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, roasted | 2.92 |
| Beef, ribeye petite roast, boneless, separable lean only, trimmed to 0″ fat, select, cooked, roasted | 2.92 |
| Beef, loin, top sirloin cap steak, boneless, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 2.92 |
| Egg, yolk, dried | 2.91 |
| Game meat, bison, chuck, shoulder clod, separable lean only, cooked, braised | 2.9 |
| Beef, round, eye of round roast, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, roasted | 2.89 |
| Beef, loin, tenderloin steak, boneless, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 2.89 |
| Pork, cured, bacon, cooked, broiled, pan-fried or roasted, reduced sodium | 2.88 |
| Veal, leg (top round), separable lean and fat, cooked, braised | 2.88 |
| Beef, loin, top sirloin filet, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, grilled | 2.88 |
| Beef, rib eye steak, bone-in, lip-on, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 2.88 |
| Beef, round, top round steak, boneless, separable lean and fat, trimmed to 0″ fat, select, cooked, grilled | 2.87 |
| Soybeans, mature seeds, roasted, salted | 2.87 |
| Soybeans, mature seeds, roasted, no salt added | 2.87 |
| Beef, round, top round, separable lean only, trimmed to 0″ fat, choice, cooked, braised | 2.85 |
| Beef, round, top round, separable lean only, trimmed to 0″ fat, select, cooked, braised | 2.85 |
| Beef, ribeye filet, boneless, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 2.85 |
| Veal, shoulder, arm, separable lean only, cooked, braised | 2.84 |
| Beef, round, top round, separable lean and fat, trimmed to 0″ fat, all grades, cooked, braised | 2.83 |
| Soy flour, full-fat, roasted | 2.83 |
| Beef, loin, tenderloin roast, separable lean only, boneless, trimmed to 0″ fat, select, cooked, roasted | 2.83 |
| Beef, plate steak, boneless, inside skirt, separable lean only, trimmed to 0″ fat, all grades, cooked, grilled | 2.82 |
| Beef, round, top round, separable lean and fat, trimmed to 0″ fat, choice, cooked, braised | 2.81 |
| Beef, round, top round, separable lean and fat, trimmed to 0″ fat, select, cooked, braised | 2.81 |
| Pork, cured, bacon, pre-sliced, cooked, pan-fried | 2.81 |
| Beef, round, top round steak, boneless, separable lean and fat, trimmed to 0″ fat, all grades, cooked, grilled | 2.81 |
| Beef, short loin, t-bone steak, bone-in, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 2.81 |
| Soy flour, full-fat, raw | 2.81 |
| Beef, round, top round steak, boneless, separable lean and fat, trimmed to 0″ fat, choice, cooked, grilled | 2.81 |
| Beef, round, eye of round steak, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, grilled | 2.81 |
| Beef, round, top round steak, boneless, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 2.8 |
| Seeds, cottonseed flour, partially defatted (glandless) | 2.8 |
| Beef, short loin, porterhouse steak, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 2.8 |
| CRACKER BARREL, grilled sirloin steak | 2.79 |
| Spices, parsley, dried | 2.79 |
| Pork, cured, bacon, cooked, baked | 2.78 |
| Veal, cubed for stew (leg and shoulder), separable lean only, cooked, braised | 2.78 |
| Beef, round, bottom round, steak, separable lean only, trimmed to 0″ fat, select, cooked, braised | 2.78 |
| Beef, rib eye roast, bone-in, lip-on, separable lean only, trimmed to 1/8″ fat, select, cooked, roasted | 2.78 |
| Beef, loin, top loin steak, boneless, lip off, separable lean only, trimmed to 0″ fat, all grades, cooked, grilled | 2.78 |
| Beef, chuck, mock tender steak, boneless, separable lean only, trimmed to 0″ fat, choice, cooked, braised | 2.78 |
| Lamb, shoulder, arm, separable lean only, trimmed to 1/4″ fat, choice, cooked, braised | 2.76 |
| Mutton, cooked, roasted (Navajo) | 2.76 |
| Beef, chuck, arm pot roast, separable lean only, trimmed to 1/8″ fat, choice, cooked, braised | 2.76 |
| Beef, top loin filet, boneless, separable lean only, trimmed to 1/8″ fat, all grades, cooked, grilled | 2.76 |
| Beef, chuck, mock tender steak, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, braised | 2.76 |
| Beef, round, top round roast, boneless, separable lean only, trimmed to 0″ fat, choice, cooked, roasted | 2.76 |
| Beef, chuck, arm pot roast, separable lean only, trimmed to 1/8″ fat, all grades, cooked, braised | 2.76 |
| Beef, chuck, arm pot roast, separable lean only, trimmed to 1/8″ fat, select, cooked, braised | 2.75 |
| Game meat, beaver, cooked, roasted | 2.75 |
| Cheese, parmesan, grated | 2.75 |
| Beef, round, top round roast, boneless, separable lean and fat, trimmed to 0″ fat, select, cooked, roasted | 2.74 |
| Lupins, mature seeds, raw | 2.74 |
| Veal, rib, separable lean only, cooked, braised | 2.74 |
| Beef, round, bottom round, steak, separable lean only, trimmed to 1/8″ fat, select, cooked, braised | 2.74 |
| Beef, loin, top sirloin petite roast, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, roasted | 2.74 |
| Beef, round, bottom round, steak, separable lean and fat, trimmed to 0″ fat, select, cooked, braised | 2.74 |
| Beef, round, top round, separable lean and fat, trimmed to 1/8″ fat, select, cooked, braised | 2.73 |
| Beef, loin, top loin steak, boneless, lip-on, separable lean only, trimmed to 1/8″ fat, all grades, cooked, grilled | 2.73 |
| Beef, round, bottom round, steak, separable lean only, trimmed to 1/8″ fat, all grades, cooked, braised | 2.73 |
| Beef, round, eye of round roast, boneless, separable lean only, trimmed to 0″ fat, choice, cooked, roasted | 2.73 |
| Game meat, deer, shoulder clod, separable lean only, cooked, braised | 2.73 |
| Beef, chuck, mock tender steak, boneless, separable lean only, trimmed to 0″ fat, select, cooked, braised | 2.73 |
| Beef, loin, top sirloin filet, boneless, separable lean only, trimmed to 0″ fat, choice, cooked, grilled | 2.73 |
| Beef, round, eye of round roast, boneless, separable lean and fat, trimmed to 0″ fat, select, cooked, roasted | 2.72 |
| Beef, round, bottom round, steak, separable lean only, trimmed to 1/8″ fat, choice, cooked, braised | 2.72 |
| Beef, round, top round, separable lean and fat, trimmed to 1/8″ fat, all grades, cooked, braised | 2.71 |
| Beef, composite of trimmed retail cuts, separable lean only, trimmed to 0″ fat, select, cooked | 2.71 |
| Beef, ribeye filet, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, grilled | 2.71 |
| Beef, round, bottom round, steak, separable lean only, trimmed to 0″ fat, all grades, cooked, braised | 2.71 |
| Veal, sirloin, separable lean only, cooked, braised | 2.7 |
| Beef, round, top round, separable lean only, trimmed to 1/8″ fat, choice, cooked, pan-fried | 2.7 |
| Beef, brisket, flat half, separable lean only, trimmed to 0″ fat, select, cooked, braised | 2.7 |
| Beef, round, top round, separable lean and fat, trimmed to 1/8″ fat, choice, cooked, braised | 2.69 |
| Beef, chuck for stew, separable lean and fat, choice, cooked, braised | 2.69 |
| Beef, chuck for stew, separable lean and fat, all grades, cooked, braised | 2.68 |
| Beef, loin, tenderloin steak, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, grilled | 2.68 |
| Beef, round, eye of round steak, boneless, separable lean only, trimmed to 0″ fat, choice, cooked, grilled | 2.68 |
| Veal, shoulder, whole (arm and blade), separable lean only, cooked, braised | 2.68 |
| Veal, shoulder, arm, separable lean and fat, cooked, braised | 2.68 |
| Beef, chuck for stew, separable lean and fat, select, cooked, braised | 2.67 |
| Beef, brisket, flat half, separable lean and fat, trimmed to 0″ fat, select, cooked, braised | 2.67 |
| Veal, loin, separable lean only, cooked, braised | 2.67 |
| Beef, variety meats and by-products, liver, cooked, braised | 2.67 |
| Beef, round, bottom round, steak, separable lean and fat, trimmed to 0″ fat, all grades, cooked, braised | 2.67 |
| Beef, rib, back ribs, bone-in, separable lean only, trimmed to 0″ fat, all grades, cooked, braised | 2.67 |
| Beef, top loin petite roast, boneless, separable lean only, trimmed to 1/8″ fat, all grades, cooked, roasted | 2.67 |
| Cheese, fontina | 2.66 |
- Branched-chain amino acid catabolism: unique segregation of pathway enzymes in organ systems and peripheral nerves. Sweatt AJ, Wood M, Suryawan A, Wallin R, Willingham MC, Hutson SM. Am J Physiol Endocrinol Metab. 2004 Jan; 286(1):E64-76.[↩]
- Pedroso JAB, Zampieri TT, Donato J. Reviewing the Effects of l-Leucine Supplementation in the Regulation of Food Intake, Energy Balance, and Glucose Homeostasis. Nutrients. 2015;7(5):3914-3937. doi:10.3390/nu7053914. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4446786/[↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩]
- Verhoeven S., Vanschoonbeek K., Verdijk L.B., Koopman R., Wodzig W.K., Dendale P., van Loon L.J. Long-term leucine supplementation does not increase muscle mass or strength in healthy elderly men. Am. J. Clin. Nutr. 2009;89:1468–1475. doi: 10.3945/ajcn.2008.26668[↩]
- Leenders M., Verdijk L.B., van der Hoeven L., van Kranenburg J., Hartgens F., Wodzig W.K., Saris W.H., van Loon L.J. Prolonged leucine supplementation does not augment muscle mass or affect glycemic control in elderly type 2 diabetic men. J. Nutr. 2011;141:1070–1076. doi: 10.3945/jn.111.138495[↩]
- Guo K., Yu Y.H., Hou J., Zhang Y. Chronic leucine supplementation improves glycemic control in etiologically distinct mouse models of obesity and diabetes mellitus. Nutr. Metab. (Lond.) 2010;7:57. doi: 10.1186/1743-7075-7-57[↩][↩][↩][↩]
- Zampieri T.T., Torres-Leal F.L., Campana A.B., Lima F.B., Donato J., Jr. l-leucine supplementation worsens the adiposity of already obese rats by promoting a hypothalamic pattern of gene expression that favors fat accumulation. Nutrients. 2014;6:1364–1373. doi: 10.3390/nu6041364[↩][↩]
- Chakrabarti P., English T., Shi J., Smas C.M., Kandror K.V. Mammalian target of rapamycin complex 1 suppresses lipolysis, stimulates lipogenesis, and promotes fat storage. Diabetes. 2010;59:775–781. doi: 10.2337/db09-1602[↩][↩]
- Polak P., Cybulski N., Feige J.N., Auwerx J., Ruegg M.A., Hall M.N. Adipose-specific knockout of raptor results in lean mice with enhanced mitochondrial respiration. Cell Metab. 2008;8:399–410. doi: 10.1016/j.cmet.2008.09.003[↩][↩]
- Teodoro G.F., Vianna D., Torres-Leal F.L., Pantaleao L.C., Matos-Neto E.M., Donato J., Jr., Tirapegui J. Leucine is essential for attenuating fetal growth restriction caused by a protein-restricted diet in rats. J. Nutr. 2012;142:924–930. doi: 10.3945/jn.111.146266[↩][↩]
- Anthony J.C., Yoshizawa F., Anthony T.G., Vary T.C., Jefferson L.S., Kimball S.R. Leucine stimulates translation initiation in skeletal muscle of postabsorptive rats via a rapamycin-sensitive pathway. J. Nutr. 2000;130:2413–2419.[↩]
- Institute of Medicine (2002). “Protein and Amino Acids”. Dietary Reference Intakes for Energy, Carbohydrates, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids. Washington, DC: The National Academies Press. pp. 589–768. https://www.nap.edu/read/10490/chapter/12[↩]
- Ropelle E.R., Pauli J.R., Fernandes M.F., Rocco S.A., Marin R.M., Morari J., Souza K.K., Dias M.M., Gomes-Marcondes M.C., Gontijo J.A., et al. A central role for neuronal amp-activated protein kinase (ampk) and mammalian target of rapamycin (mtor) in high-protein diet-induced weight loss. Diabetes. 2008;57:594–605. doi: 10.2337/db07-0573[↩][↩][↩]
- Blouet C., Jo Y.H., Li X., Schwartz G.J. Mediobasal hypothalamic leucine sensing regulates food intake through activation of a hypothalamus-brainstem circuit. J. Neurosci. 2009;29:8302–8311. doi: 10.1523/JNEUROSCI.1668-09.2009[↩][↩][↩]
- Zeanandin G., Balage M., Schneider S.M., Dupont J., Hebuterne X., Mothe-Satney I., Dardevet D. Differential effect of long-term leucine supplementation on skeletal muscle and adipose tissue in old rats: An insulin signaling pathway approach. Age. 2012;34:371–387. doi: 10.1007/s11357-011-9246-0[↩]
- Li X., Wang X., Liu R., Ma Y., Guo H., Hao L., Yao P., Liu L., Sun X., He K., et al. Chronic leucine supplementation increases body weight and insulin sensitivity in rats on high-fat diet likely by promoting insulin signaling in insulin-target tissues. Mol. Nutr. Food Res. 2013;57:1067–1079. doi: 10.1002/mnfr.201200311[↩]
- Koch C.E., Goddeke S., Kruger M., Tups A. Effect of central and peripheral leucine on energy metabolism in the djungarian hamster (phodopus sungorus) J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2013;183:261–268. doi: 10.1007/s00360-012-0699-y[↩][↩]
- Freudenberg A., Petzke K.J., Klaus S. Dietary l: -leucine and l: -alanine supplementation have similar acute effects in the prevention of high-fat diet-induced obesity. Amino Acids. 2012;44:519–528. doi: 10.1007/s00726-012-1363-2[↩]
- Freudenberg A., Petzke K.J., Klaus S. Comparison of high-protein diets and leucine supplementation in the prevention of metabolic syndrome and related disorders in mice. J. Nutr. Biochem. 2012;23:1524–1530. doi: 10.1016/j.jnutbio.2011.10.005[↩]
- Drgonova J., Jacobsson J.A., Han J.C., Yanovski J.A., Fredriksson R., Marcus C., Schioth H.B., Uhl G.R. Involvement of the neutral amino acid transporter slc6a15 and leucine in obesity-related phenotypes. PLoS ONE. 2013;8:e68245. doi: 10.1371/journal.pone.0068245[↩][↩]
- Zhang Y., Guo K., LeBlanc R.E., Loh D., Schwartz G.J., Yu Y.H. Increasing dietary leucine intake reduces diet-induced obesity and improves glucose and cholesterol metabolism in mice via multimechanisms. Diabetes. 2007;56:1647–1654. doi: 10.2337/db07-0123[↩]
- Garcia-Espinosa M.A., Wallin R., Hutson S.M., Sweatt A.J. Widespread neuronal expression of branched-chain aminotransferase in the cns: Implications for leucine/glutamate metabolism and for signaling by amino acids. J. Neurochem. 2007;100:1458–1468[↩]
- Sisk C.L., Nunez A.A., Thebert M.M. Differential effects of electrolytic and chemical hypothalamic lesions on lh pulses in rats. Am. J. Physiol. 1988;255:E583–E590[↩]
- Pizzi W.J., Barnhart J.E., Fanslow D.J. Monosodium glutamate admlinistration to the newborn reduces reproductive ability in female and male mice. Science. 1977;196:452–454. doi: 10.1126/science.557837.[↩]
- Layman D.K., Walker D.A. Potential importance of leucine in treatment of obesity and the metabolic syndrome. J. Nutr. 2006;136:319S–323S.[↩]
- Badawy AA-B, Lake SL, Dougherty DM. Mechanisms of the Pellagragenic Effect of Leucine: Stimulation of Hepatic Tryptophan Oxidation by Administration of Branched-Chain Amino Acids to Healthy Human Volunteers and the Role of Plasma Free Tryptophan and Total Kynurenines. International Journal of Tryptophan Research : IJTR. 2014;7:23-32. doi:10.4137/IJTR.S18231. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4259507/[↩]
- Rajavel Elango, Karen Chapman, Mahroukh Rafii, Ronald O Ball, Paul B Pencharz; Determination of the tolerable upper intake level of leucine in acute dietary studies in young men, The American Journal of Clinical Nutrition, Volume 96, Issue 4, 1 October 2012, Pages 759–767, https://doi.org/10.3945/ajcn.111.024471[↩][↩]
- United States Department of Agriculture Agricultural Research Service. USDA Food Composition Databases. https://ndb.nal.usda.gov/ndb/search/list[↩]
{kind=link}