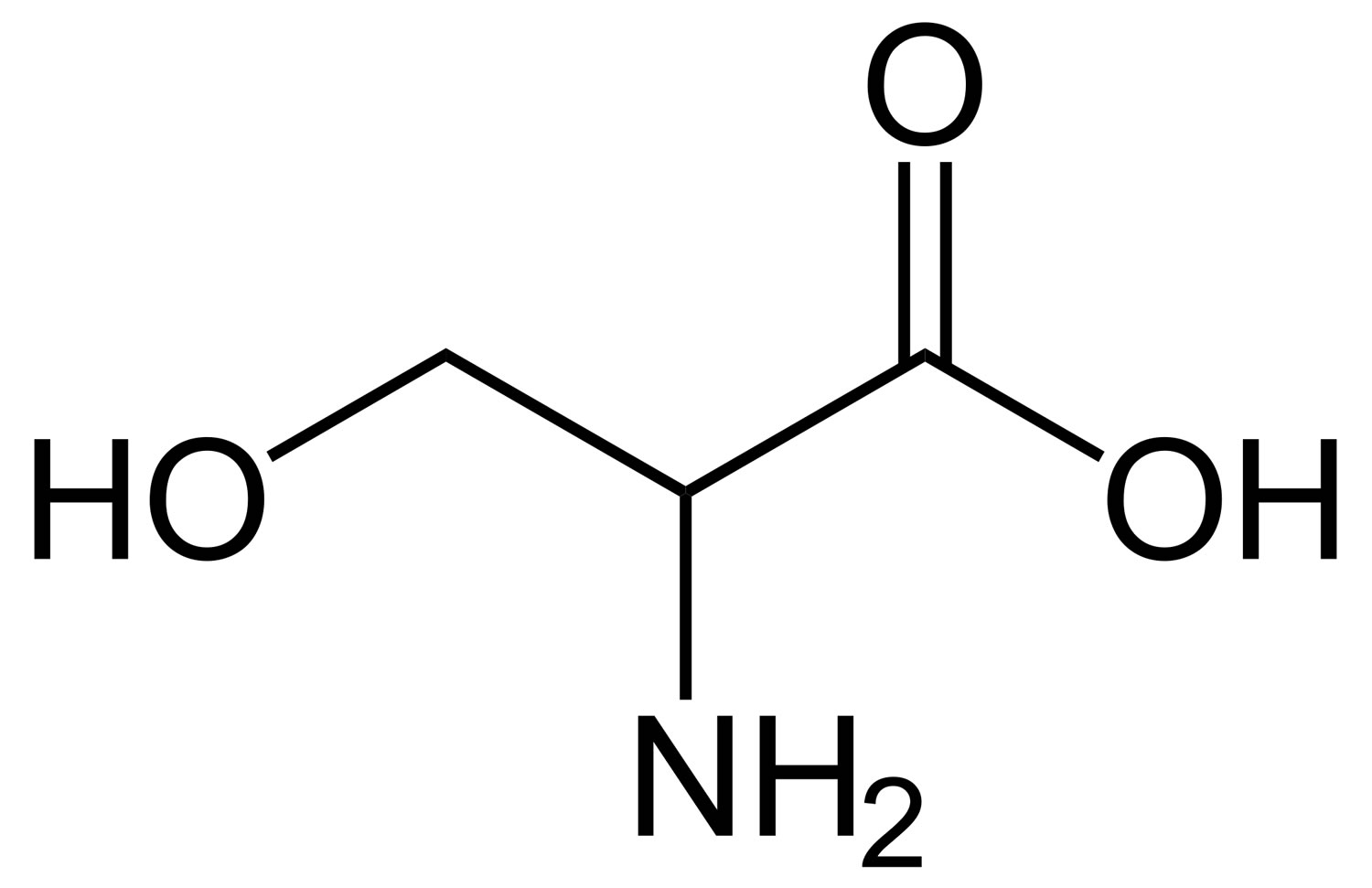
What is serine
Serine is a nonessential amino acid since it is synthesized in your body from other metabolites, including glycine. Serine can also be derived from your diet and the degradation of protein and/or phospholipids 1. Serine was first obtained from silk protein, a particularly rich source. Its name is derived from the Latin for silk, sericum. Only the L-stereoisomer (L-serine) appears naturally in proteins. L-serine is required to synthesize membrane lipids such as phosphatidylserine and sphingolipids 2. Serine is highly concentrated in all cell membranes. Unlike other amino acids, L-serine is used directly for the synthesis of phosphatidylserine and ceramide, the hydrophobic moiety of sphingolipids. The latter sphingolipids act as important mediators of the signaling cascades involved in various cellular functions, such as apoptosis, proliferation, and the stress response 3. The first step of sphingolipid synthesis is the condensation of L-serine and palmitoyl-CoA catalyzed by serine palmitoyltransferase, which is a rate-limiting enzyme in the sphingolipid synthetic pathway (Figure 2A) 4. Like all the amino acid building blocks of protein and peptides, serine can become essential under certain conditions, and is thus important in maintaining health and preventing disease. Low-average concentration of serine compared to other amino acids is found in muscle. L-Serine may be derived from four possible sources: dietary intake; biosynthesis from the glycolytic intermediate 3-phosphoglycerate; from glycine ; and by protein and phospholipid degradation 5. Little data is available on the relative contributions of each of these four sources of L-serine to serine homoeostasis. It is very likely that the predominant source of L-serine will be very different in different tissues and during different stages of human development.
In liver tissue, the serine biosynthetic pathway is regulated in response to dietary and hormonal changes. Of the three synthetic enzymes, the properties of 3-phosphoglycerate dehydrogenase (3- PGDH) and phosphoserine phosphatase are the best documented. Hormonal factors such as glucagon and corticosteroids also influence 3-phosphoglycerate dehydrogenase (3- PGDH) and phosphoserine phosphatase activities in interactions dependent upon the diet. L-serine plays a central role in cellular proliferation. L-Serine is the predominant source of one-carbon groups for the de novo synthesis of purine nucleotides and deoxythymidine monophosphate. It has long been recognized that, in cell cultures, L-serine is a conditional essential amino acid, because it cannot be synthesized in sufficient quantities to meet the cellular demands for its utilization. In recent years, L-serine and the products of its metabolism have been recognized not only to be essential for cell proliferation, but also to be necessary for specific functions in the central nervous system. The findings of altered levels of serine and glycine in patients with psychiatric disorders and the severe neurological abnormalities in patients with defects of L-serine synthesis underscore the importance of L-serine in brain development and function 5.
Figure 1. Serine biosynthesis
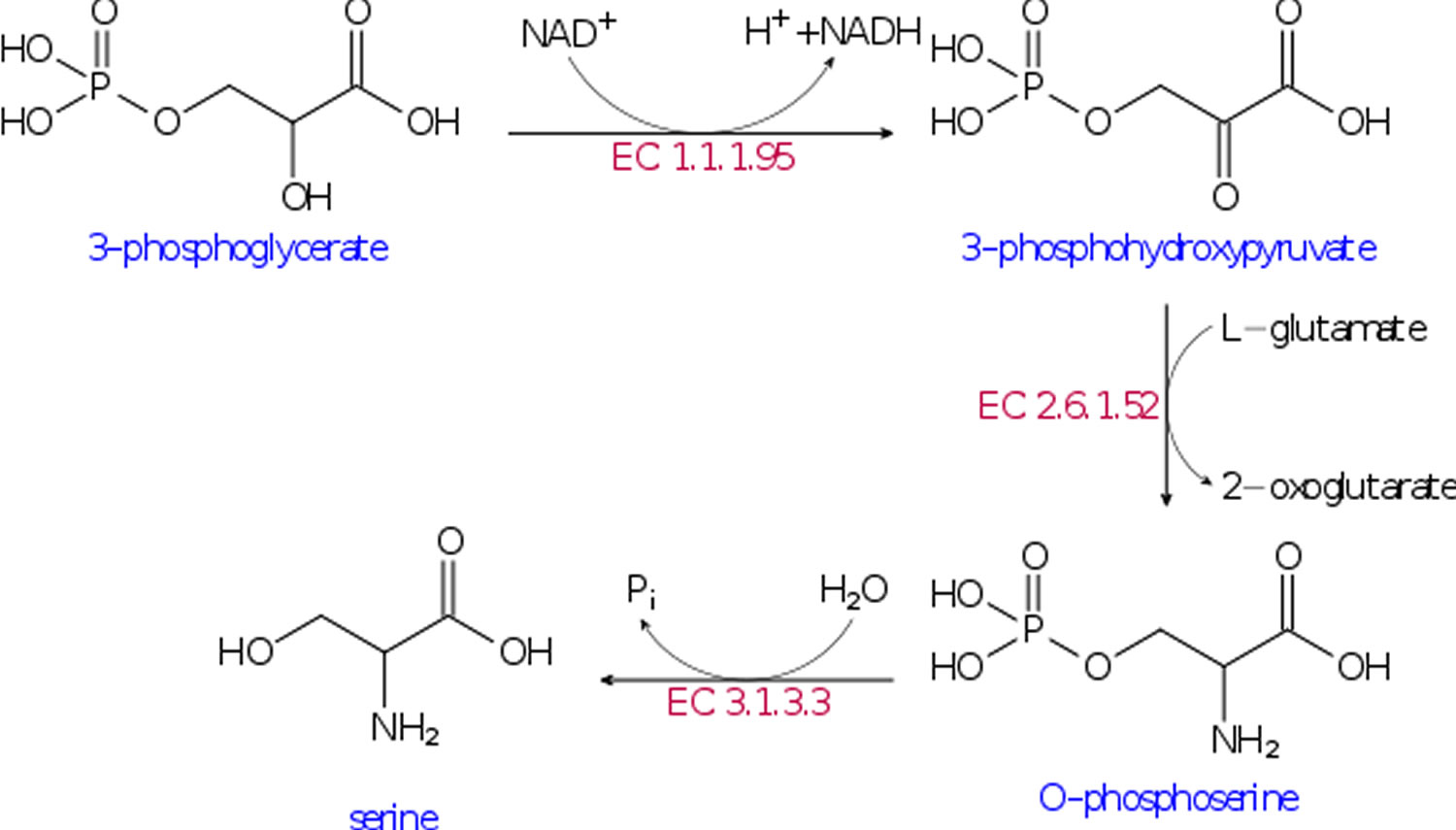
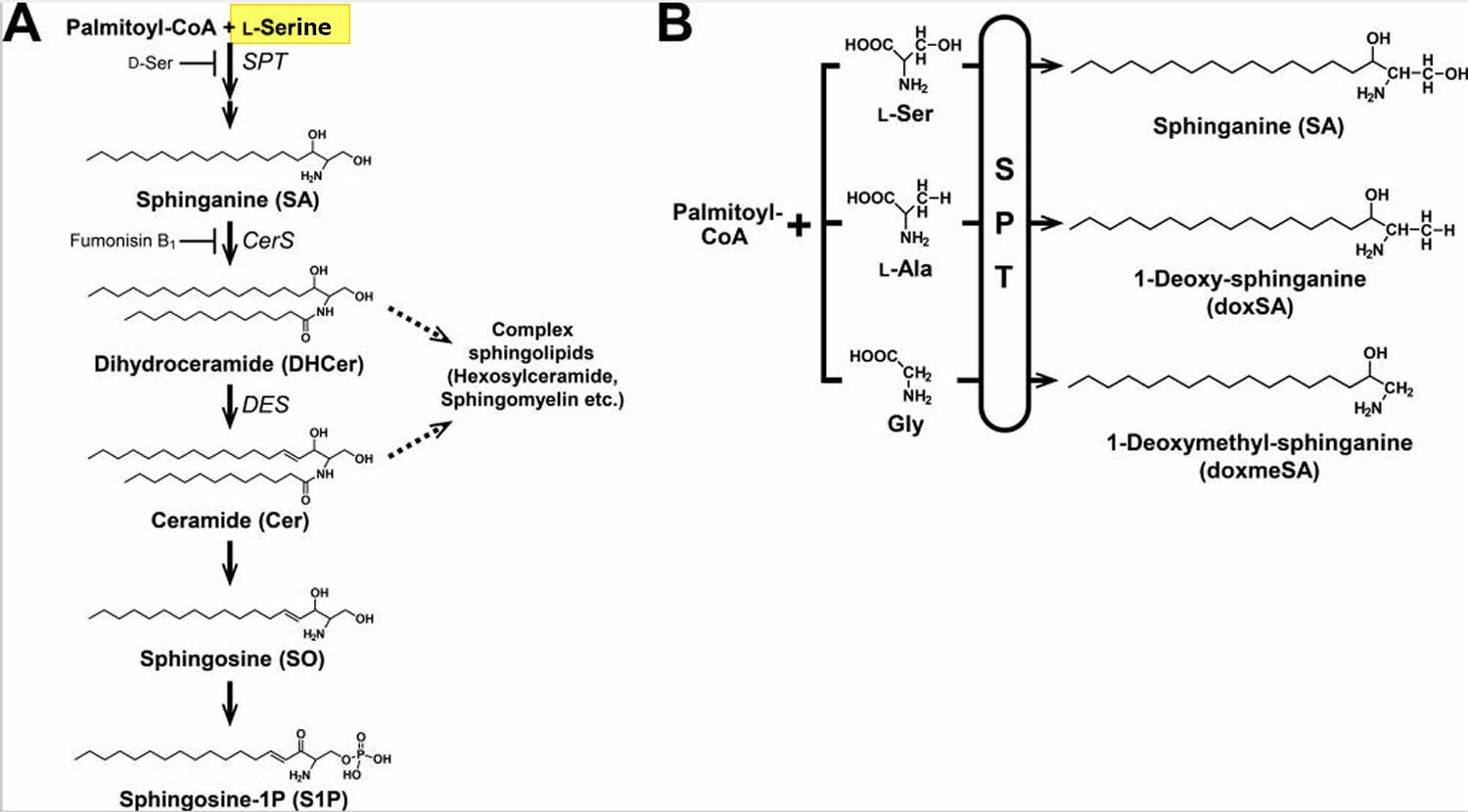
Figure 2. Sphingolipid biosynthetic pathway
Footnotes:
(A) L-Serine and palmitoyl-CoA are condensed by serine palmitoyltransferase (SPT) to form 3-keto-sphinganine, which is reduced to sphinganine (SA). Then, sphinganine (SA) is N-acylated to form dihydroceramide (DHCer) and desaturated to form ceramide (Cer). In addition, ceramide (Cer) is converted to sphingosine (SO) and subsequently phosphorylated to generate sphingosine 1-phosphate (S1P) by the degradative pathway.
(B) Sphingoid bases generated by serine palmitoyltransferase (SPT), which uses not only L-Serine but also L-Alanine or Glycine as a substrate and synthesizes atypical 1-deoxysphingolipids (doxSLs). Because of the lack of the C1 hydroxyl group, 1-deoxysphingolipids (doxSLs) can neither be converted to complex sphingolipids nor degraded by the degradative pathway.
These results indicate that 1-deoxysphingolipids (doxSLs) are generated under conditions of imbalance between L-Alanine and L-Serine in nonmalignant cells and tissues, suggesting a possible role of 1-deoxysphingolipids (doxSLs) in the pathobiology of L-Serine deficiency disorders and other diseases involving an increased ratio of L-Alanine to L-Serine in serum and/or tissues.
Thus, understanding of molecular interactions between 1-deoxysphingolipids (doxSLs) and cellular components, and their consequences, is expected to facilitate the development of therapeutic strategies against the cellular damage evoked by L-Serine deficiency and other metabolic disorders.
[Source 2]L-Serine is synthesized de novo from a glycolytic intermediate, 3-phosphoglycerate, through three catalytic steps known as the phosphorylated pathway 7. The first step in this pathway is catalyzed by 3-phosphoglycerate dehydrogenase (3- PGDH) 8. Phosphohydroxypyruvate is metabolized to phosphoserine by phosphohydroxypyruvate aminotransferase and, finally, phosphoserine is converted into L-serine by phosphoserine phosphatase. Humans with mutated 3-phosphoglycerate dehydrogenase (3- PGDH) have lower levels of free L-Serine in the plasma and in cerebrospinal fluid. These L-Serine-deficient patients exhibit severe neurological symptoms, including congenital microcephaly, psychomotor retardation, and intractable seizures 9. In addition to these data from humans with 3-phosphoglycerate dehydrogenase (3- PGDH) deficiency, animal study demonstrated that conventional d-3-phosphoglycerate dehydrogenase knock-out mice display severe consequences of embryonic development, such as brain malformation with overall growth retardation, and die after embryonic day 13.5 10. These human and mouse studies highlighted the absolute necessity of L-Serine, synthesized de novo via the phosphorylated pathway, for embryonic viability and nervous system development. In addition, recent functional genomic studies showed that de novo biosynthesis of L-Serine plays a crucial role in invasiveness, malignant transformation, and proliferation of certain types of cancer 11. Many cancer cells consume serine in preference to glycine 12. Cancer cells selectively consumed exogenous serine, which was converted to intracellular glycine and one-carbon units for building nucleotides 13. Restriction of exogenous glycine or depletion of the glycine cleavage system did not impede proliferation. In the absence of serine, uptake of exogenous glycine was unable to support nucleotide synthesis. Indeed, higher concentrations of glycine inhibited proliferation. Under these conditions, glycine was converted to serine, a reaction that would deplete the one-carbon pool. Providing one-carbon units by adding formate rescued nucleotide synthesis and growth of glycine-fed cells. L-Serine also acts on pyruvate kinase M2 as an allosteric effector and supports aerobic glycolysis, which is a metabolic hallmark of cancer cells 11. These reports provide unexpected evidence that enhanced L-Serine availability in the body is involved in intrinsic metabolic reprogramming of cancer cells 2 and that nucleotide synthesis and cancer cell proliferation are supported by serine—rather than glycine—consumption 13.
Malignant development is accompanied by genetic changes in cancer cells that drive abnormal proliferation, growth, survival, and invasion. Each of these phenotypes is supported by changes in cellular metabolism, and several metabolic enzymes have been identified as oncogenes or tumor suppressors 14. Although alterations in glucose and glutamine metabolism are central to metabolic transformation 15, recent studies have focused on the role of the nonessential amino acids serine and glycine in supporting tumor growth 16. In addition to their role in protein synthesis, serine and glycine contribute to anabolic pathways important for the generation of glutathione, nucleotides, phospholipids, and other metabolites 16 (see Figure 3). The requirement for intracellular serine and glycine for the support of cell growth and division is therefore clear. However, how cancer cells obtain these nutrients (uptake versus biosynthesis) and how they metabolize them remains to be fully elucidated. Mechanistic insight into this question will significantly enhance our ability to target serine/glycine metabolism for therapeutic gain.
Figure 3. Intracellular serine and glycine metabolism
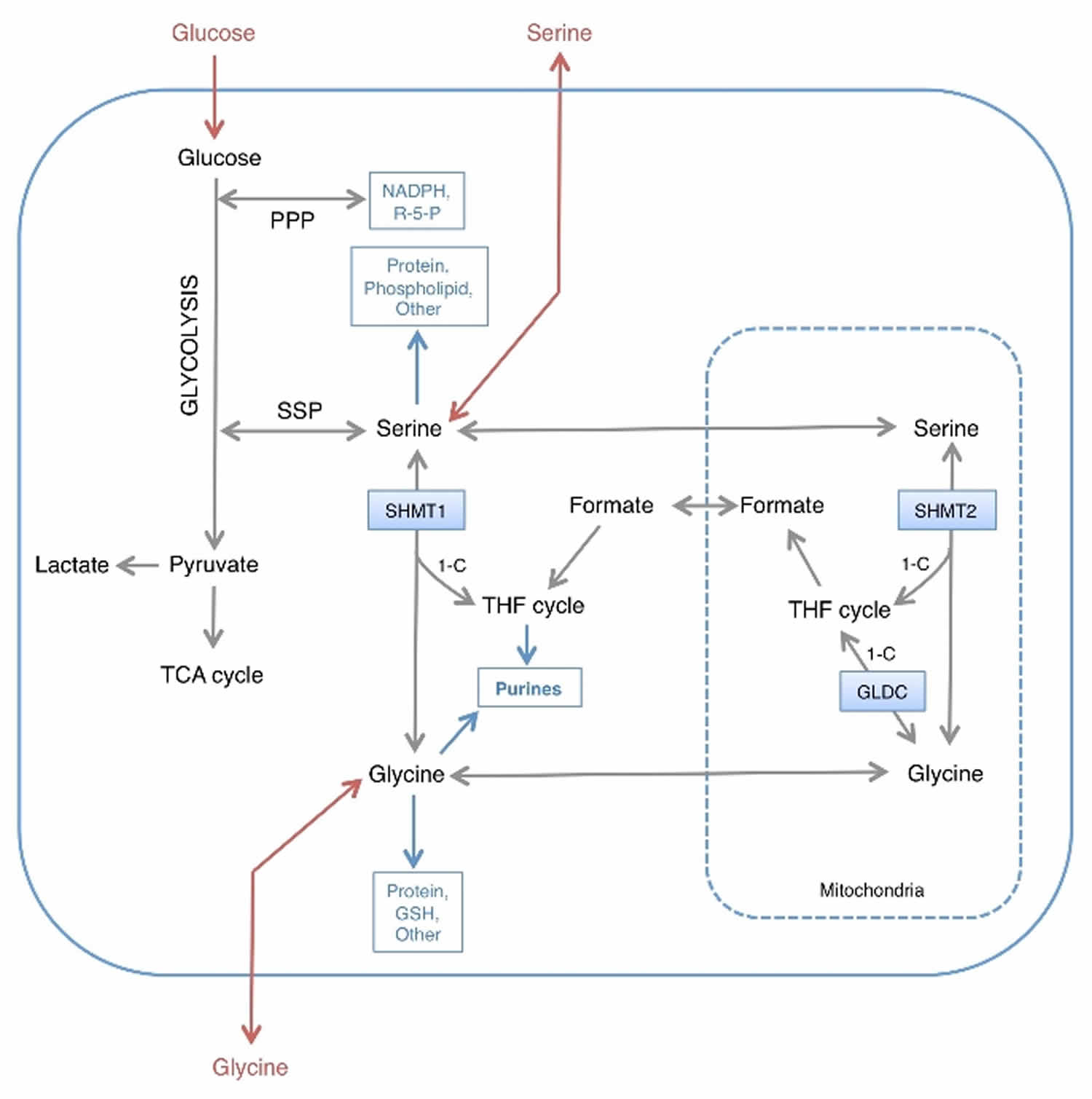
Footnotes: 1‐C; one carbon unit transferred to and from the tetrahydrofolate (THF) cycle.
Abbreviations: R‐5‐P = ribose 5‐phosphate; PPP = Pentose Phosphate Pathway; SSP = Serine synthesis pathway.
[Source 13]Figure 4. Nucleotide synthesis and cancer cell proliferation are supported by serine rather than glycine
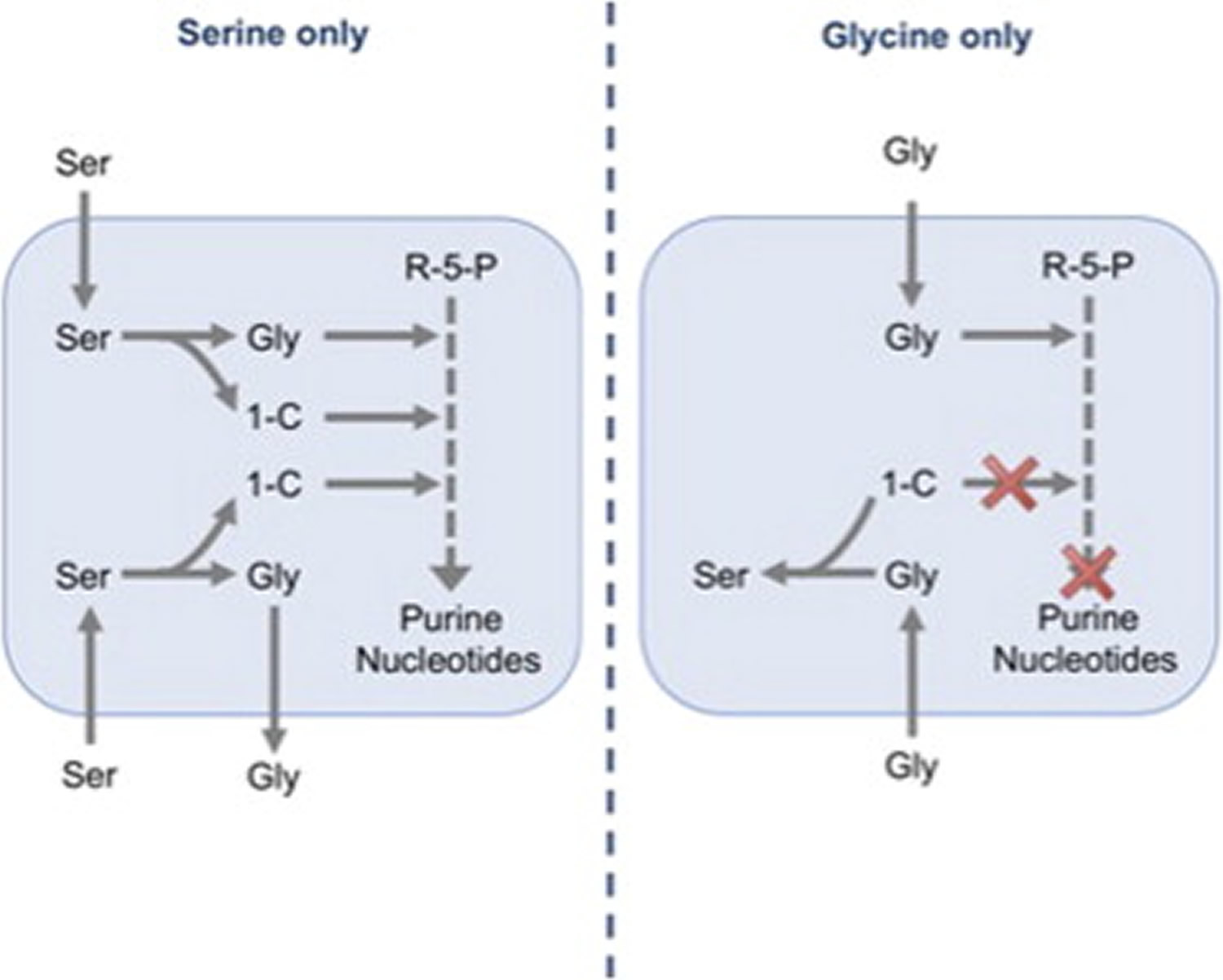
Amplification of 3-phosphoglycerate dehydrogenase (3- PGDH), the first enzyme of the de novo serine synthesis pathway, has been found in breast cancers and melanomas 17. Many tumor cells, however, remain highly dependent on uptake of exogenous serine 18, suggesting that in these cells, de novo serine synthesis alone cannot support the requirements for proliferation. A recent study showed that glycine uptake is correlated with rapid proliferation 19; this contrasts with other studies showing that excess dietary glycine has an inhibitory effect on tumorigenesis in multiple in vivo models 20. Another study suggests that cancer cells fail to consume glycine when serine is plentiful 18.
The present lack of clarity is due, at least in part, to the complexity of serine and glycine metabolism, which can be carried out by mitochondrial and cytoplasmic pathways 21, both of which are upregulated in cancer 19. Serine can be converted to glycine by serine hydroxymethyl transferase (cytoplasmic, SHMT1; mitochondrial, SHMT2), a reaction that yields one-carbon units, which enter the tetrahydrofolate (THF) cycle and are critical for nucleotide synthesis. Glycine can also be cleaved by the mitochondrial glycine cleavage system to yield one-carbon units that are transferred to the THF cycle 21 (see Figure 3). Amplification of GLDC (a component of the glycine cleavage system) in cancers 22 suggests that this pathway is an important source of one-carbon units. In addition to cleavage, glycine can also be converted into serine by SHMT1 and SHMT2. Taken together, these reactions allow serine and glycine to be metabolized into the same set of metabolic precursors, suggesting that serine and glycine may be used interchangeably and may be equally effective in supporting proliferation (Figure 3).
In this study 13 the authors demonstrated that exogenous glycine cannot substitute for serine for the support of cancer cell proliferation. Tracking the intracellular fate of exogenous serine and glycine showed that in the absence of exogenous serine, glycine does not enter the one-carbon cycle, but is converted into serine, a process that consumes rather than produces one-carbon units and prevents nucleotide synthesis (see Figure 4). Consistent with this, the authors show that the inability of cells to grow in glycine could be rescued by addition of formate, which directly supplements the one-carbon pool for nucleotide synthesis.
Serine deficiency
Serine deficiency disorders are caused by a defect in one of the three synthesizing enzymes of the L-serine biosynthesis pathway 23. Serine deficiency disorders give rise to a neurological phenotype with psychomotor retardation, microcephaly and seizures in newborns and children or progressive polyneuropathy in adult patients. There are three defects that cause serine deficiency of which 3-phosphoglycerate dehydrogenase (3-PGDH) deficiency, the defect affecting the first step in the pathway, has been reported most frequently. The other two disorders in L-serine biosynthesis phosphoserine aminotransferase deficiency and phosphoserine phosphatase deficiency have been reported only in a limited number of patients 23. The biochemical hallmarks of all three disorders are low concentrations of serine in cerebrospinal fluid and plasma. Prompt recognition of affected patients is important, since serine deficiency disorders are treatable causes of neurometabolic disorders. The use of age-related reference values for serine in CSF and plasma can be of great help in establishing a correct diagnosis of serine deficiency, in particular in newborns and young children.
Foods high in serine
Table 1. Serine foods (ordered from highest to low)
| Description | Serine (g) Value Per 100 grams |
| Egg, white, dried, stabilized, glucose reduced | 6.16 |
| Egg, white, dried, powder, stabilized, glucose reduced | 6.08 |
| Egg, white, dried, flakes, stabilized, glucose reduced | 5.67 |
| Egg, white, dried | 5.59 |
| Soy protein isolate | 4.59 |
| Soy protein isolate, potassium type | 4.59 |
| Egg, whole, dried | 3.77 |
| Egg, whole, dried, stabilized, glucose reduced | 3.66 |
| Soy protein concentrate, produced by alcohol extraction | 3.37 |
| Soy protein concentrate, produced by acid wash | 3.37 |
| Seaweed, spirulina, dried | 3 |
| Beverages, Protein powder soy based | 2.96 |
| Egg, yolk, dried | 2.81 |
| Seeds, sesame flour, low-fat | 2.73 |
| Soy flour, defatted | 2.73 |
| Soy meal, defatted, raw | 2.6 |
| Gelatins, dry powder, unsweetened | 2.6 |
| Snacks, pork skins, plain | 2.6 |
| Peanut flour, defatted | 2.57 |
| Fish, cod, Atlantic, dried and salted | 2.56 |
| Seeds, cottonseed flour, low fat (glandless) | 2.49 |
| Snacks, pork skins, barbecue-flavor | 2.48 |
| Seeds, cottonseed meal, partially defatted (glandless) | 2.46 |
| Whale, beluga, meat, dried (Alaska Native) | 2.42 |
| Cheese, parmesan, shredded | 2.4 |
| Soybeans, mature seeds, raw | 2.36 |
| Soybeans, mature seeds, dry roasted | 2.29 |
| Seeds, sunflower seed flour, partially defatted | 2.27 |
| Tofu, dried-frozen (koyadofu) | 2.26 |
| Tofu, dried-frozen (koyadofu), prepared with calcium sulfate | 2.26 |
| Meat extender | 2.23 |
| Mollusks, whelk, unspecified, cooked, moist heat | 2.22 |
| Seeds, sesame flour, partially defatted | 2.2 |
| Cheese, parmesan, hard | 2.07 |
| Seeds, cottonseed flour, partially defatted (glandless) | 2.05 |
| Soybeans, mature seeds, roasted, salted | 2.04 |
| Soybeans, mature seeds, roasted, no salt added | 2.04 |
| Soy flour, full-fat, roasted | 2.02 |
| Soy flour, full-fat, raw | 2 |
| Leavening agents, yeast, baker’s, active dry | 1.98 |
| Milk, dry, nonfat, regular, without added vitamin A and vitamin D | 1.97 |
| Milk, dry, nonfat, regular, with added vitamin A and vitamin D | 1.97 |
| Milk, dry, nonfat, calcium reduced | 1.93 |
| Milk, dry, nonfat, instant, with added vitamin A and vitamin D | 1.91 |
| Milk, dry, nonfat, instant, without added vitamin A and vitamin D | 1.91 |
| Fish, caviar, black and red, granular | 1.9 |
| Gelatin desserts, dry mix, reduced calorie, with aspartame, added phosphorus, potassium, sodium, vitamin C | 1.88 |
| Gelatin desserts, dry mix, reduced calorie, with aspartame, no added sodium | 1.88 |
| Lupins, mature seeds, raw | 1.87 |
| Milk, buttermilk, dried | 1.87 |
| Cheese, romano | 1.84 |
| Seeds, safflower seed meal, partially defatted | 1.79 |
| Cheese, roquefort | 1.77 |
| Cheese, gruyere | 1.72 |
| Seeds, hemp seed, hulled | 1.71 |
| Cheese, parmesan, grated | 1.69 |
| Seeds, sesame flour, high-fat | 1.68 |
| Seeds, pumpkin and squash seed kernels, dried | 1.67 |
| Peanut flour, low fat | 1.67 |
| Seeds, pumpkin and squash seed kernels, roasted, without salt | 1.65 |
| Seeds, pumpkin and squash seed kernels, roasted, with salt added | 1.65 |
| Cheese, swiss | 1.64 |
| Nuts, butternuts, dried | 1.64 |
| Pork, cured, bacon, cooked, microwaved | 1.63 |
| Seeds, cottonseed kernels, roasted (glandless) | 1.63 |
| Nuts, mixed nuts, oil roasted, without peanuts, without salt added | 1.62 |
| Nuts, mixed nuts, oil roasted, without peanuts, with salt added | 1.62 |
| Cheese, parmesan, dry grated, reduced fat | 1.61 |
| Cheese, edam | 1.55 |
| Cheese, gouda | 1.54 |
| Beef, round, top round roast, boneless, separable lean only, trimmed to 0″ fat, select, cooked, roasted | 1.52 |
| Beef, round, eye of round roast, boneless, separable lean only, trimmed to 0″ fat, select, cooked, roasted | 1.51 |
| Seeds, watermelon seed kernels, dried | 1.51 |
| Snacks, soy chips or crisps, salted | 1.5 |
| Beef, loin, top sirloin filet, boneless, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 1.5 |
- Genome-scale metabolic modelling of hepatocytes reveals serine deficiency in patients with non-alcoholic fatty liver disease. Nature Communications volume 5, Article number: 3083,2014. https://www.nature.com/articles/ncomms4083.pdf[↩]
- Esaki K, Sayano T, Sonoda C, et al. l-Serine Deficiency Elicits Intracellular Accumulation of Cytotoxic Deoxysphingolipids and Lipid Body Formation. The Journal of Biological Chemistry. 2015;290(23):14595-14609. doi:10.1074/jbc.M114.603860. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4505526/[↩][↩][↩]
- Hannun Y. A., Obeid L. M. (2008) Principles of bioactive lipid signalling: lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 9, 139–150[↩]
- Lowther J., Naismith J. H., Dunn T. M., Campopiano D. J. (2012) Structural, mechanistic and regulatory studies of serine palmitoyltransferase. Biochem. Soc. Trans. 40, 547–554[↩]
- De Koning TJ, Snell K, Duran M, Berger R, Poll-The B-T, Surtees R. L-serine in disease and development. Biochemical Journal. 2003;371(Pt 3):653-661. doi:10.1042/BJ20021785. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1223326/pdf/12534373.pdf[↩][↩]
- Serine. https://en.wikipedia.org/wiki/Serine[↩]
- Ichihara A., Greenberg D. M. (1955) Pathway of serine formation from carbohydrate in rat liver. Proc. Natl. Acad. Sci. U.S.A. 41, 605–609[↩]
- Snell K. (1984) Enzymes of serine metabolism in normal, developing and neoplastic rat tissues. Adv. Enzyme Regul. 22, 325–400[↩]
- de Koning T. J., Klomp L. W., van Oppen A. C., Beemer F. A., Dorland L., van den Berg I., Berger R. (2004) Prenatal and early postnatal treatment in 3-phosphoglycerate-dehydrogenase deficiency. Lancet 364, 2221–2222[↩]
- Kawakami Y., Yoshida K., Yang J. H., Suzuki T., Azuma N., Sakai K., Hashikawa T., Watanabe M., Yasuda K., Kuhara S., Hirabayashi Y., Furuya S. (2009) Impaired neurogenesis in embryonic spinal cord of Phgdh knockout mice, a serine deficiency disorder model. Neurosci. Res. 63, 184–193[↩]
- Chaneton B., Hillmann P., Zheng L., Martin A. C., Maddocks O. D., Chokkathukalam A., Coyle J. E., Jankevics A., Holding F. P., Vousden K. H., Frezza C., O’Reilly M., Gottlieb E. (2012) Serine is a natural ligand and allosteric activator of pyruvate kinase M2. Nature 491, 458–462[↩][↩]
- Serine, but Not Glycine, Supports One-Carbon Metabolism and Proliferation of Cancer Cells. Cell Reports Volume 7, ISSUE 4, P1248-1258, May 22, 2014. https://doi.org/10.1016/j.celrep.2014.04.045[↩]
- Serine, but Not Glycine, Supports One-Carbon Metabolism and Proliferation of Cancer Cells. Cell Reports Volume 7, Issue 4, P1248-1258, May 22, 2014. https://doi.org/10.1016/j.celrep.2014.04.045[↩][↩][↩][↩][↩]
- Shaw, R.J.Cantley, L.C. Decoding key nodes in the metabolism of cancer cells: sugar & spice and all things nice. F1000 Biol. Rep. 2012; 4: 2[↩]
- Jones, N.P.Schulze, A. Targeting cancer metabolism—aiming at a tumour’s sweet-spot. Drug Discov. Today. 2012; 17: 232-241[↩]
- Locasale, J.W. Serine, glycine and one-carbon units: cancer metabolism in full circle.Nat. Rev. Cancer. 2013; 13: 572-583[↩][↩]
- Phosphoglycerate dehydrogenase diverts glycolytic flux and contributes to oncogenesis.Nat. Genet. 2011; 43: 869-874[↩]
- Serine starvation induces stress and p53-dependent metabolic remodelling in cancer cells.Nature. 2013; 493: 542-546[↩][↩]
- Metabolite profiling identifies a key role for glycine in rapid cancer cell proliferation.Science. 2012; 336: 1040-1044[↩][↩]
- Dietary glycine inhibits the growth of B16 melanoma tumors in mice.Carcinogenesis. 1999; 20: 793-798[↩]
- Compartmentalization of Mammalian folate-mediated one-carbon metabolism.Annu. Rev. Nutr. 2010; 30: 57-81[↩][↩]
- Glycine decarboxylase activity drives non-small cell lung cancer tumor-initiating cells and tumorigenesis.Cell. 2012; 148: 259-272[↩]
- An update on serine deficiency disorders. J Inherit Metab Dis. 2013 Jul;36(4):613-9. DOI:10.1007/s10545-013-9592-4[↩][↩]
- United States Department of Agriculture Agricultural Research Service. USDA Food Composition Databases. https://ndb.nal.usda.gov/ndb/search/list[↩]
{kind=link}