Contents
Mansonellosis
Mansonellosis is a parasitic filarial roundworm infection caused by the Mansonella parasites (Mansonella ozzardi, and Mansonella perstans [Dipetalonema perstans], and Mansonella streptocerca [Dipetalonema streptocerca]) in the family Onchocercidae that is transmitted by tiny blood-sucking midges Ceratopogonidae called Culicoides milnei and Culicoides grahamii mostly found in many parts of sub-Saharan Africa (West, East, and Central Africa) and parts of South and Central America (Argentina, Brazil, Bolivia, Colombia, Guyana, Suriname and Venezuela, and several Caribbean islands), but black flies (Simuliidae) are also known to play a role in the transmission of Mansonella ozzardi in parts of Latin America 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15. Of the three Mansonella species, Mansonella ozzardi is the major species in Latin America and the Caribbean, while Mansonella perstans and Mansonella streptocerca are reported in Africa 16. The greatest burden of Mansonella infection is observed in sub-Saharan Africa, where an estimated 114 million individuals are infected, while nearly 600 million people are at risk of contracting mansonellosis 17, 18. In Africa, the main Ceratopogonidae species suspected to be responsible for Mansonella transmission include Culicoides grahamii, Culicoides austeni, Culicoides inornatipennis, Culicoides milnei, Culicoides hortensis, Culicoides krameri, and Culicoides vitshumbieni, although the taxonomic identities of these species have been debated over time 19, 20. The classification of tropical Culicoides species remains incomplete, complicating the accurate identification of vectors in endemic areas. These blood sucking midges have a short, stout, and sharply pointed vertical proboscis, allowing them to take blood meals essential for female egg maturation 20. When a blood sucking midge fly bites a person with mansonellosis, it consumes microfilariae or microscopic immature worms, from the infected person’s skin. The Mansonella parasite develops in the midge or black fly and penetrate the midgut wall and migrate to the thoracic muscles becomes infectious third-stage larvae (L3) (see Mansonella Life Cycle below) 18, 21.
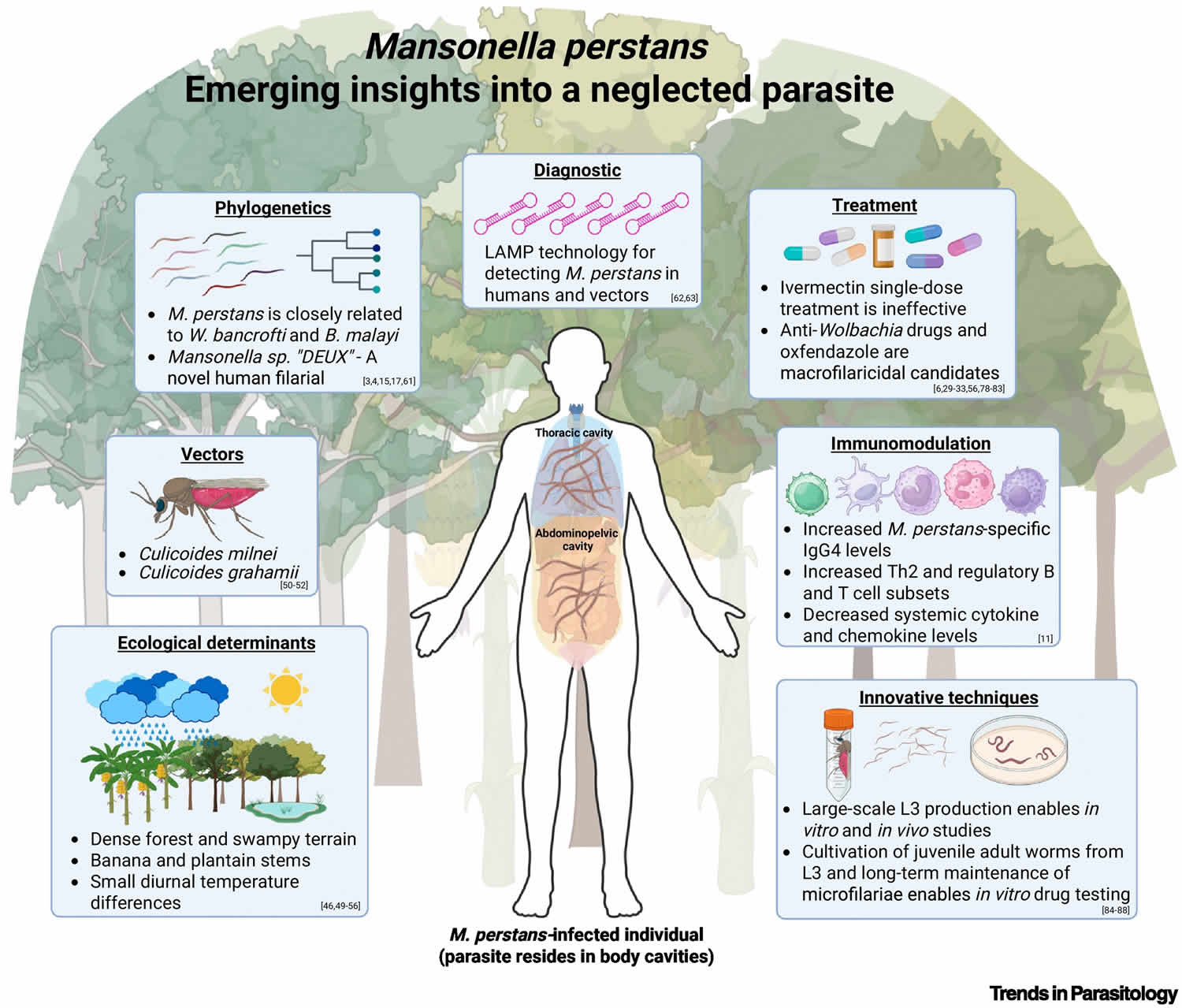
In 2015, a new potential human-infecting Mansonella species designated as Mansonella sp. “DEUX” has been discovered in febrile children in a hospital-based study in Gabon 22, 23, 22, 24. The Mansonella sp. “DEUX” is capable of infecting both humans and apes 22, 23. Further investigations showed that Mansonella sp. “DEUX” was the most frequent filarial species, as during a cross-sectional study in rural Fougamou, Gabon, 35% of individuals carried the Mansonella sp. “DEUX” parasite 24.
Historically, the term “mansonellosis” and its synonym “mansonelliasis” were used to describe Mansonella ozzardi infection in humans until the mid-1980s 25, 26, 27, 28, 29, 30. The modern use of the term mansonellosis began only after Orihel 31 and Eberhard 32 revised the genus Mansonella and added Mansonella perstans and Mansonella streptocerca. Prior to this, a number of systems of nomenclature were used to describe Mansonella parasite infections. For example, mansonellosis was usually restricted to infections caused by Mansonella ozzardi 33, 34 and the terms “perstans filariasis” and “streptocerciasis” were often used to describe infections with Mansonella perstans and Mansonella streptocerca, respectively 35, 36, 37, 38.
Futhermore, a key topic of debate over the past decade has been whether Mansonella perstans harbors endosymbiotic bacteria of the genus Wolbachia, which play essential roles in the development and reproduction of most filarial species, with the exception of Loa loa 39, 40, 41, 42. Two earlier studies reported no evidence of Wolbachia in Mansonella perstans 42, 43, possibly due to insufficient genetic information for designing effective PCR (polymerase chain reaction) primers. However, subsequent studies using improved molecular detection methods have identified the presence of Wolbachia in Mansonella perstans, consistent with positive outcomes from doxycycline treatment trials 44, 45, 46, 47, 48. Wolbachia were also detected in individuals mono infected with Mansonella sp “DEUX”, confirming carriage of endosymbionts also by this new species/genotype 24. As Wolbachia are usually transferred maternally and because of the known genetic variability of this endosymbiont among nematodes, Wolbachia genomes could give a further hint on the genetic relatedness of their host species 49.
Most people infected with the parasitic worms that cause mansonellosis never develop symptoms (asymptomatic) or can be associated with nonspecific clinical signs and symptoms such as fever, headache, joint pain (arthralgia), and eyelesions (Mansonella ozzardi); itch (pruritus), joint pain (arthralgia), abdominal pain, soft tissue swelling (angioedema), skin rash, and fatigue (Mansonella perstans and perhaps Mansonella ‘DEUX’); and itchy dermatitis and chronic lymph gland inflammation (Mansonella perstans) 50, 4.
Mansonellosis signs and symptoms typically include 18, 51, 52, 53:
- Occasional swelling
- Impaired vision, if worms enter your eye
- Itching
- Rashes
- Joint pain (arthralgia)
- Abdominal (belly) pain
- Enlargement of lymph glands (lymphadenopathy)
- Pericarditis (inflammation of the outer lining of the heart)
- Fever
- Feeling weak and tired (fatigue)
If you have traveled to an area where Mansonella parasites are spread and develop any of the symptoms, talk with your doctor about getting a blood test. Mansonella perstans and Mansonella ozzardi are usually diagnosed by the finding of microfilariae (immature worms) circulating in your blood whereas Mansonella streptocerca is usually diagnosed by finding microfilariae in skin snips 54. The rare human cases of Mansonella rodhaini were also diagnosed via skin snips. Concentration techniques (e.g. Knott’s concentration, polycarbonate membrane filtration) may increase sensitivity. Note that these Mansonella parasitic worms do not generally exhibit periodicity, though in some regions, Mansonella perstans-infected patients may exhibit slightly elevated microfilaremia in morning hours. Skin snips should be thick enough to include the outer part of the dermal papillae but not so thick as to produce bleeding (a ~2 mm wide section should be sufficient) 54. Skin snips should be placed immediately in normal saline or distilled water, just enough to cover the specimen 54. Microfilariae tend to emerge more rapidly in saline, however in either medium the microfilariae typically emerge in 30—60 minute and can be seen in wet mount preparations 54. For a definitive diagnosis, allow the wet mount to dry, fix in methanol, and stain with Giemsa or hematoxylin-and-eosin (H&E) 54. Serological assays developed for the detection of Mansonella species–specific antibodies or antigens are generally of limited utility due to cross-reactions with other filarial species and potentially other roundworms (nematodes) 54.
Due to the largely asymptomatic clinical presentation of Mansonella infection, most people do not require treatment for mansonellosis 53. However, some experts recommend that human infections with Mansonella species be treated, regardless of any apparent clinical manifestations 4. If you are a diagnosed with mansonellosis, your doctor will discuss treatment options, if needed, with you 53. There are currently no specific treatment guidelines or approved medications targeting Mansonella parasite 3. Only a limited number of studies have evaluated the efficacy of drugs against Mansonella perstans, and these investigations often involve small groups, making it difficult to draw robust conclusions. Furthermore, many of these studies have been conducted on travellers or expatriates returning to Europe or North America from endemic regions 6, raising concerns about the generalizability of findings due to potential immunological differences between individuals from endemic and non-endemic areas.
The available data on anthelminth and antibiotic treatments are limited and often contradictory 3. Single-dose regimens of commonly used anthelmintics such as albendazole, praziquantel, and ivermectin, used in mass drug administration (MDA) for onchocerciasis, lymphatic filariasis, or schistosomiasis, have shown minimal or no impact on microfilarial loads in infected individuals 18, 55, 56. In contrast, multiple-dose regimens using drugs such as thiabendazole, mebendazole, diethylcarbamazine (DEC), or a combination of mebendazole and diethylcarbamazine (DEC), have demonstrated greater efficacy 57. The mebendazole–diethylcarbamazine (DEC) combination achieved the highest clearance rate, with a reported 37% reduction in microfilariae 56. Among the most promising treatments is the antibiotic doxycycline, which targets Wolbachia endosymbionts. Doxycycline was shown to clear Mansonella perstans microfilariae from the peripheral blood for the observational period of up to 2 years 6, 44, 45, indicating a filarial sterilizing and/or macrofilaricidal effect. However, its requirement for a 6-week treatment course poses logistical challenges for use in large-scale control programmes.
New macrofilaricidal drugs allowing shorter treatment regimens, such as oxfendazole, are currently under investigation 3. Oxfendazole is a pan-nematode candidate which has been used for several decades as an anthelmintic in pets 3. Given oxfendazole’s proven efficacy in filarial animal models following a 5-day treatment, and its selective macrofilaricidal efficacy with no strong direct microfilaricidal efficacy 58, it is expected that oxfendazole does not cause microfilariae-induced adverse events in microfilaraemic individuals. An EDCTP3-funded Phase 2 clinical trial (PACTR202412611774752) is ongoing in Cameroon, Gabon, the Democratic Republic of the Congo, and Tanzania to evaluate oxfendazole’s efficacy against Mansonella perstans, Loa loa, Onchocerca volvulus, as well as trichuriasis.
Novel anti-Wolbachial macrofilaricidal candidates such as Corallopyronin A, which is expected to allow treatment regimens of 14 days or less 59, 60, and is currently under preparation for Phase 1 clinical trials in humans 61, are expected to be efficacious for all Wolbachia-containing filariae, that is, filariae causing onchocerciasis, lymphatic filariasis, and mansonellosis. Moreover, combinations of anti-Wolbachials with oxfendazole or albendazole are suggested to provide synergistic effects and may allow shorter treatment regimens and improve drug efficacy 62.
Figure 1. Mansonella perstans
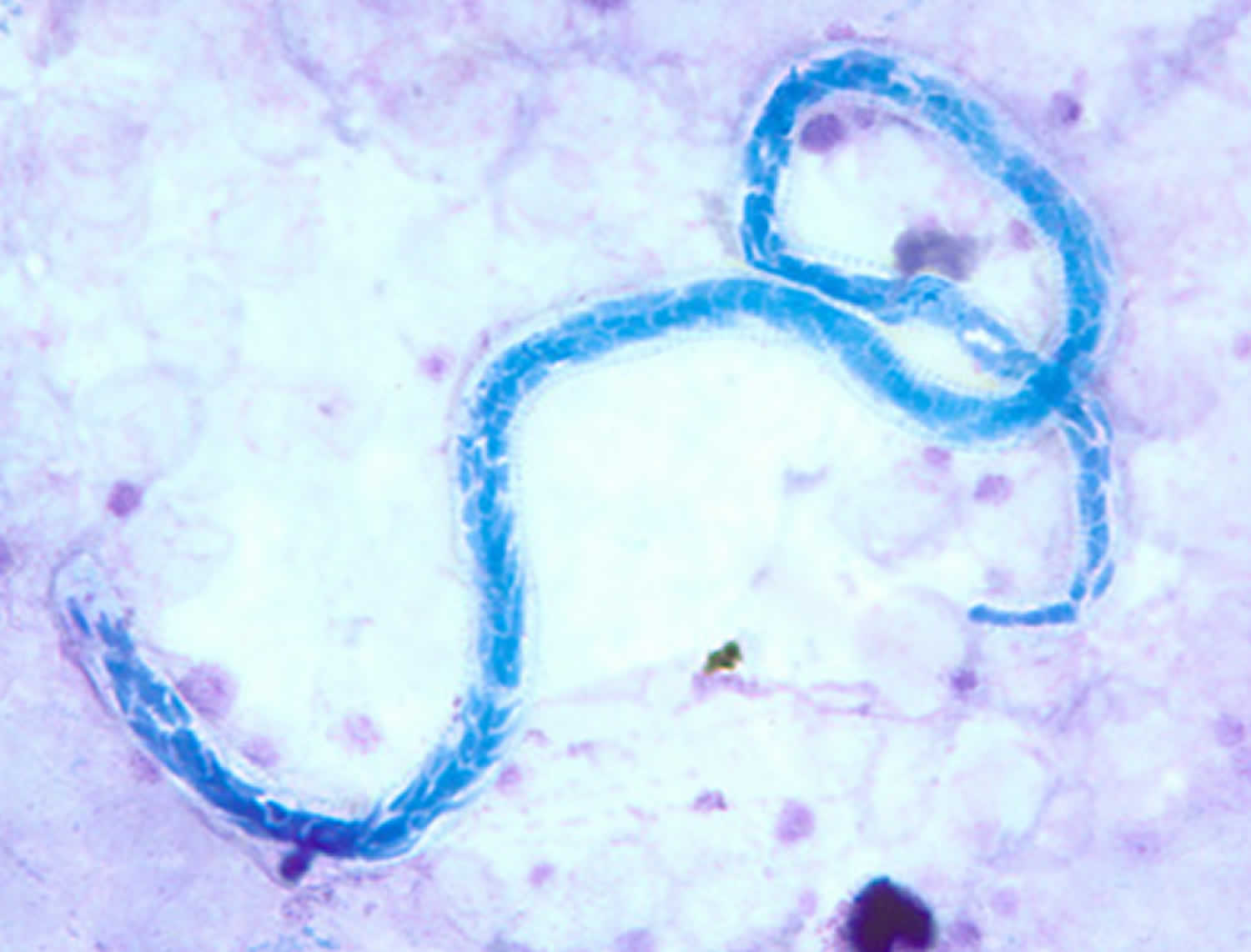
Footnotes: Mansonella perstans microfilaria in a thick blood smear stained with Giemsa, from a patient from Cameroon.
[Source 54 ]Figure 2. Mansonella ozzardi
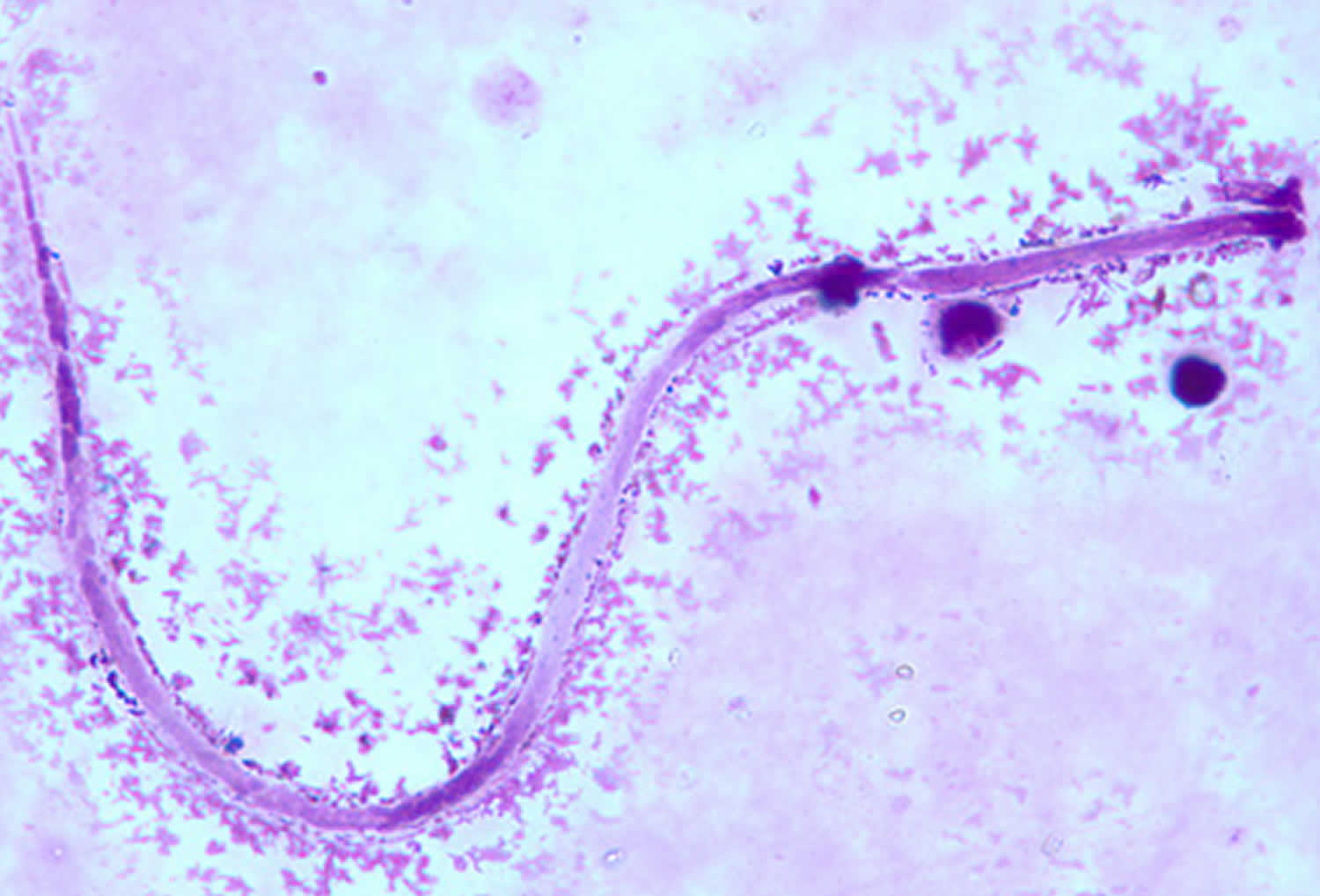
Footnotes: Mansonella ozzardi microfilaria in a thick blood smear, stained with Giemsa.
[Source 54 ]Figure 3. Mansonella streptocerca
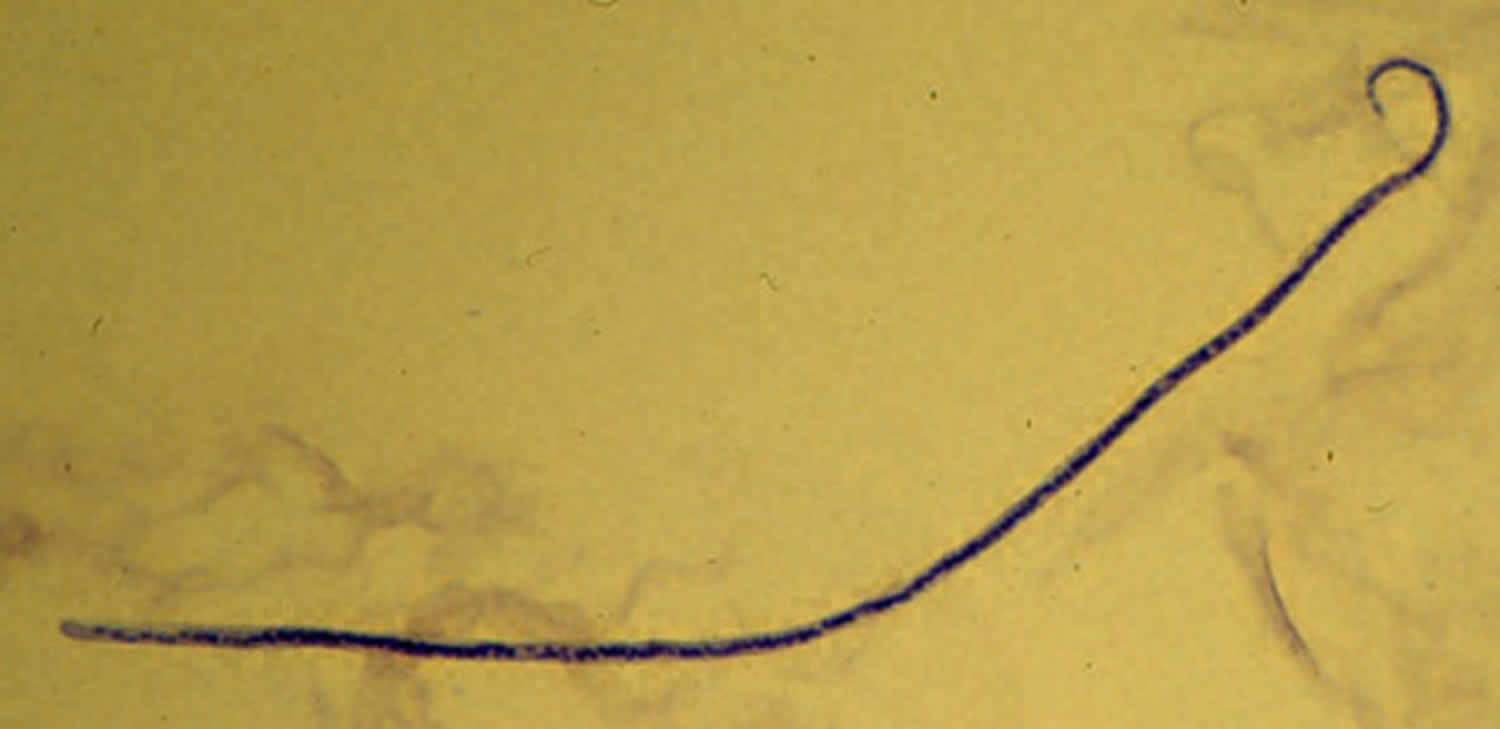
Footnotes: Mansonella streptocerca microfilaria fixed in 2% formalin and stained with hematoxylin.
[Source 54 ]Figure 4. Culicoides milnei

Footnotes: Blood sucking midges Culicoides milnei transmit Mansonella perstans 63
[Source 3 ]Figure 5. Mansonellosis pathophysiology
Footnotes: Mansonella perstans pathophysiology
[Source 3 ]What causes mansonellosis?
Mansonellosis is a parasitic filarial roundworm infection caused by the Mansonella parasites (Mansonella ozzardi, and Mansonella perstans [Dipetalonema perstans], and Mansonella streptocerca [Dipetalonema streptocerca]) in the family Onchocercidae that is transmitted by tiny blood-sucking midges Ceratopogonidae called Culicoides milnei and Culicoides grahamii mostly found in many parts of sub-Saharan Africa (West, East, and Central Africa) and parts of South and Central America (Argentina, Brazil, Bolivia, Colombia, Guyana, Suriname and Venezuela, and several Caribbean islands), but blackflies (Simuliidae) are also known to play a role in the transmission of Mansonella ozzardi in parts of Latin America 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15. In Africa, the main Ceratopogonidae species suspected to be responsible for Mansonella transmission include Culicoides grahamii, Culicoides austeni, Culicoides inornatipennis, Culicoides milnei, Culicoides hortensis, Culicoides krameri, and Culicoides vitshumbieni, although the taxonomic identities of these species have been debated over time 19, 20. The classification of tropical Culicoides species remains incomplete, complicating the accurate identification of vectors in endemic areas. These blood sucking midges have a short, stout, and sharply pointed vertical proboscis, allowing them to take blood meals essential for female egg maturation 20. When a blood sucking midge fly bites a person with mansonellosis, it consumes microfilariae or microscopic immature worms, from the infected person’s skin. The Mansonella parasite develops in the midge or black fly and penetrate the midgut wall and migrate to the thoracic muscles becomes infectious third-stage larvae (L3) (see Mansonella Life Cycle below) 18, 21.
In 2015, a new potential human-infecting Mansonella species designated as Mansonella sp. “DEUX” has been discovered in febrile children in a hospital-based study in Gabon 22, 23, 22, 24. The Mansonella sp. “DEUX” is capable of infecting both humans and apes 22, 23. Further investigations showed that Mansonella sp. “DEUX” was the most frequent filarial species, as during a cross-sectional study in rural Fougamou, Gabon, 35% of individuals carried the Mansonella sp. “DEUX” parasite 24.
Females of all known tiny blood-sucking midges Ceratopogonidae or Simulium black flies consume blood meals in order for their eggs to mature 64, 65. Microfilariae (immature worms) are ingested by the midge when a female midge takes a blood meal from a host infected with mansonellosis. The microfilariae penetrate the insect’s gut and go through several maturation stages in the thoracic flight muscles over 6–12 days before migrating to the head and proboscis, from where they can be transferred to a primate when the midge takes its next blood meal 11. During the blood meal, the third-stage (infective) filarial larvae (L3 larvae) actively break out of the mouthparts and penetrate into skin via the bite wound before migrating and maturing into adult worms. Males and female adult worms mate, and female worms begin to produce unsheathed microfilariae that circulate in the blood (Mansonella perstans and Mansonella ozzardi) or diffuse into the skin (Mansonella streptocerca and Mansonella ozzardi) of the primate host 6. Practical diagnostic guides often describe all three species (Mansonella perstans, Mansonella streptocerca and Mansonella ozzardi) as having aperiodic microfilariae that circulate in peripheral blood or skin throughout the day and night 6, although reports of cryptic periodicity for both Mansonella perstans and Mansonella ozzardi exist 11, 66, 67.
Mansonella perstans is considered the most common of the mansonellosis parasites and is endemic in a large portion of sub-Saharan Africa, as well as a northern part of the Amazon rainforest stretching from equatorial Brazil to the Caribbean coast of South America, with molecular analysis suggesting that the parasite arrived in South America relatively recently and most likely as a consequence of the slave trade 36, 68, 69. From the mid-1920s until the early 1970s, Mansonella perstans was frequently recorded as occurring in Papua New Guinea (PNG), but there is very little primary-source data to support this and no recent survey data that indicate its occurrence anywhere in the Eastern hemisphere outside continental Africa 70. Overall, more than 100 million people may be infected by Mansonella perstans, and it is estimated that 600 million people live at high risk of contracting an infection in Africa alone 36.
Mansonella streptocerca seems to have a distribution limited to continental Africa, where it occurs in the tropical rainforest areas of central and west Africa, as well as in Uganda 11, 71, 68. Perhaps because the skin-snip biopsies required for its epidemiological monitoring are more invasive and painful than the blood sampling required for Mansonella perstans surveys, or perhaps because blood sampling is simply more commonly done for other epidemiological studies, there have been fewer epidemiological surveys investigating Mansonella streptocerca epidemiology 6. As with Mansonella perstans and Mansonella ozzardi and many other filarial infections, prevalence rates of Mansonella streptocerca are highest in the oldest age-groups studied and higher among men than women in endemic areas 68, 72, 38. Mansonella streptocerca is transmitted by various species of Culicoides and some of the same vector species that transmit Mansonella perstans, although there are too few data available to evaluate properly whether its distribution is influenced by the same ecological drivers as Mansonella perstans 29, 73, 74. The few epidemiological studies that have tried to link patent Mansonella streptocerca infections with clinical symptoms have encountered similar rates of skin disease among those infected and those uninfected in their study areas 72, 75.
Mansonella ozzardi has a patchy geographic distribution across Latin America. It has been recorded from southern Mexico to northwestern Argentina, but has not been reported in Chile, Uruguay, or Paraguay 10, 11, 71, 76, 77. Mansonella ozzardi parasite occurs on several Caribbean Islands, and like Mansonella perstans has even been recorded in Papua New Guinea, although recent reports of its occurrence in the region are notable by their absence 11, 70. In the Caribbean region, Mansonella ozzardi parasite is transmitted by a diverse range of biting midges from the family Ceratopogonidae, whereas in Central and South America it is known only to be transmitted by biting midges from the genus Culicoides and blackflies from the genus Simulium 6. Because of these vectorial differences, it was once thought that Mansonella ozzardi from the Caribbean islands and those from continental South America might be different species 78, 79. Artificial-infection experiments and the recent publication of DNA sequences, however, strongly indicate that parasites from the two regions are morphologically, biologically, and genetically near identical, and thus that they are most parsimoniously regarded as just one species 80, 31, 81, 82, 69, 83.
Table 1. Mansonella perstans, Mansonella ozzardi, and Mansonella streptocerca main characteristics
| Mansonella perstans | Mansonella streptocerca | Mansonella ozzardi | ||
|---|---|---|---|---|
| Microfilaria tail |
|
|
| |
| Sheath in microfilariae |
|
|
| |
| Microfilariae: periodicity and localization |
|
|
| |
| Microfilaria: length × diameter | 200×4–5 mm | 180–240×3–5 mm | 163–203×3–5 mm | |
| Adults measurements | ♂ 3.5–4.5 cm, ♀ 5–8 cm | ♂ 1.3–1.8 cm, ♀ 2.7 cm | ♂ 2.4–2.8 cm, ♀ 3.2–8.1 cm | |
| Adults localization | Serous body cavities, mainly peritoneal, but may also appear subcutaneously | Subcutaneous tissues of the dermis | Thoracic cavity and mesenteries of the peritoneal cavity | |
| Vector | Africa | Culicoides spp. (biting midges) | Culicoides spp. (biting midges) |
Simuliidae
Ceratopogonidae
|
| Central America | ||||
| South America | Culicoides spp. (biting midges) | |||
| Caribbean |
| |||
| Geographical distribution | Central and west Africa, South America | Central and west Africa, western Uganda | Latin America and Caribbean islands | |
| Reported clinical symptoms |
|
|
| |
| Treatment |
|
|
| |
Mansonella Life Cycle
The three species of Mansonella parasite which cause mansonellosis in humans vary in biological, clinical, and epidemiological characteristics 6. The life cycles for all three Mansonella parasite species are similar, involving development in two hosts: an insect and a primate 6. Humans are the only known natural vertebrate host for Mansonella ozzardi, but other primates can serve as natural hosts for Mansonella perstans and Mansonella streptocerca 80, 81. All of the known insect vectors of mansonellosis are from the Diptera taxonomic grouping. The known vectors of Mansonella perstans and Mansonella streptocerca are from the biting-midge genus Culicoides. Mansonella ozzardi, however, is not exclusively transmitted by Culicoides 6. A range of non-Culicoides Ceratopogonidae biting midges and black-flies from the genus Simulium are known to be capable of transmitting Mansonella ozzardi 6.
Mansonella perstans life cycle
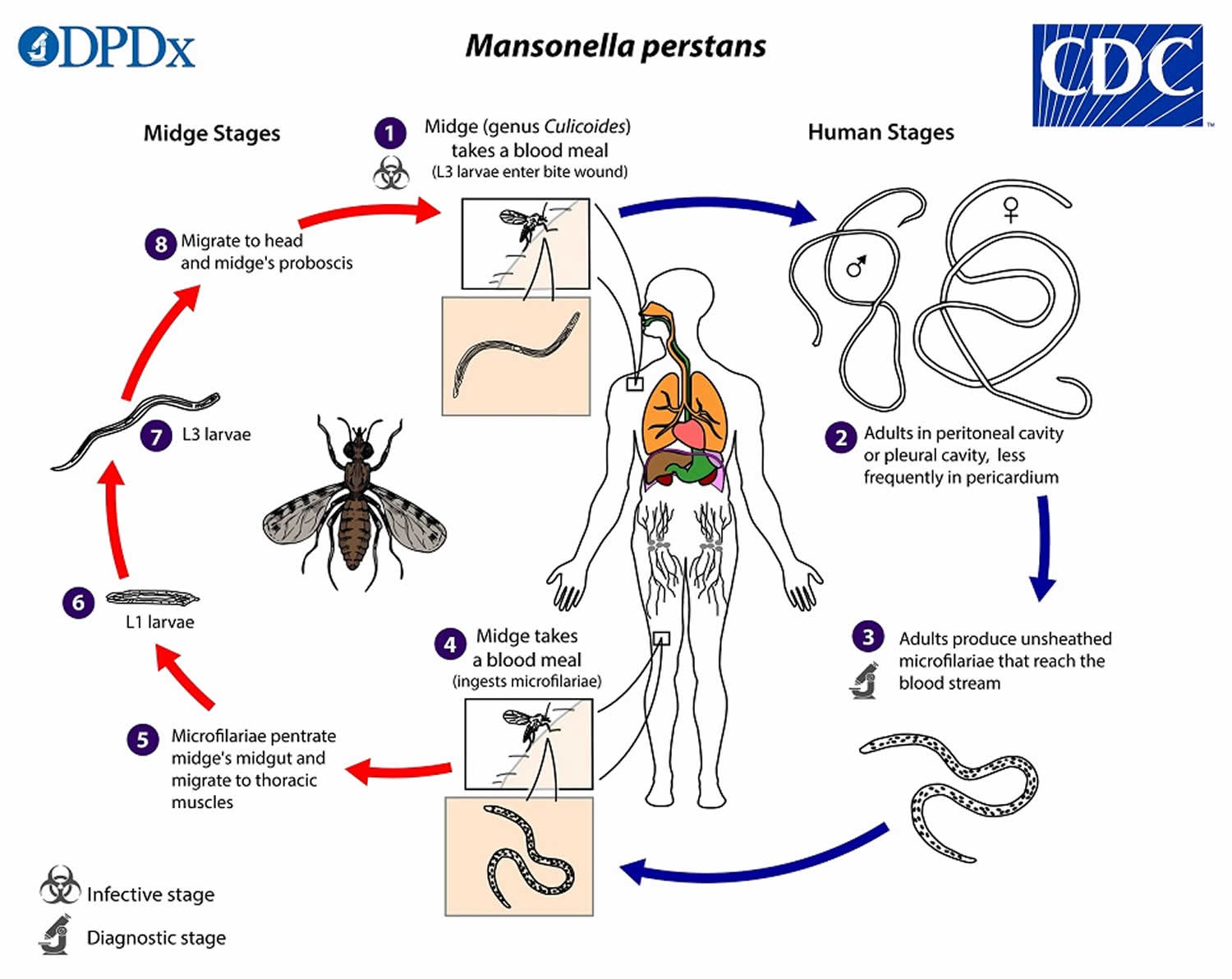
During a blood meal, an infected midge (Culicoides species) introduces Mansonella perstans third-stage (L3) larvae onto the skin of the human host, where they penetrate into the bite wound (number 1). The Mansonella perstans third-stage infective (L3) larvae develop into adults that reside in body cavities, most commonly the peritoneal cavity or pleural cavity, and less frequently in the pericardium (number 2). The size range for female Mansonella perstans worms is 70 to 80 mm in length and 120 µm in diameter, and the males measure approximately 45 mm by 60 µm. Mansonella perstans adult worms produce unsheathed, subperiodic microfilariae (immature filarial worms) (200 µm long, 4.5 µm wide) which enter peripheral circulation (number 3). Microfilariae circulate in the peripheral blood during both day and night 84, 85. However, one study detected a subtle pattern of periodicity, with slightly higher concentrations observed in the early morning, possibly correlating with the peak biting activity of the Culicoides midge 86. A Culicoides midge vector ingests microfilariae during a blood meal (number 4). After ingestion, the microfilariae migrate from the midge’s midgut through the hemocoel to the thoracic muscles of the arthropod (number 5). There the microfilariae develop into first-stage larvae (number 6) and subsequently into third-stage infective larvae (L3 larvae) (number 7). The third-stage infective larvae (L3 larvae) migrate to the midge’s proboscis (number 8) and can infect another human when the midge takes a blood meal (number 1) 87, 18.
Despite this general understanding of the Mansonella perstans life cycle, many aspects of its development within the human host remain unclear 3. For instance, the potential for transplacental transmission (the transmission of Mansonella parasites from a mother to her fetus across the placenta during pregnancy) has not been conclusively determined, with conflicting findings reported in the literature 88, 89. Furthermore, it remains to be clarified whether all infected individuals harbor detectable microfilariae in their peripheral blood, or if amicrofilaremic (a condition in which a person is infected with filarial parasites but has no detectable microfilariae in their blood or skin) cases exist, as is common in other filarial infections 3. This distinction is critical for mapping and surveillance, as current prevalence estimates rely on the detection of microfilariae in peripheral blood. Furthermore, a key topic of debate over the past decade has been whether Mansonella perstans harbors endosymbiotic bacteria of the genus Wolbachia, which play essential roles in the development and reproduction of most filarial species, with the exception of Loa loa 40, 41, 42. Two earlier studies reported no evidence of Wolbachia in Mansonella perstans 42, 43, possibly due to insufficient genetic information for designing effective PCR (polymerase chain reaction) primers. However, subsequent studies using improved molecular detection methods have identified the presence of Wolbachia in Mansonella perstans, consistent with positive outcomes from doxycycline treatment trials 44, 45, 46, 47, 48.
Figure 6. Mansonella perstans life cycle
Mansonella ozzardi life cycle
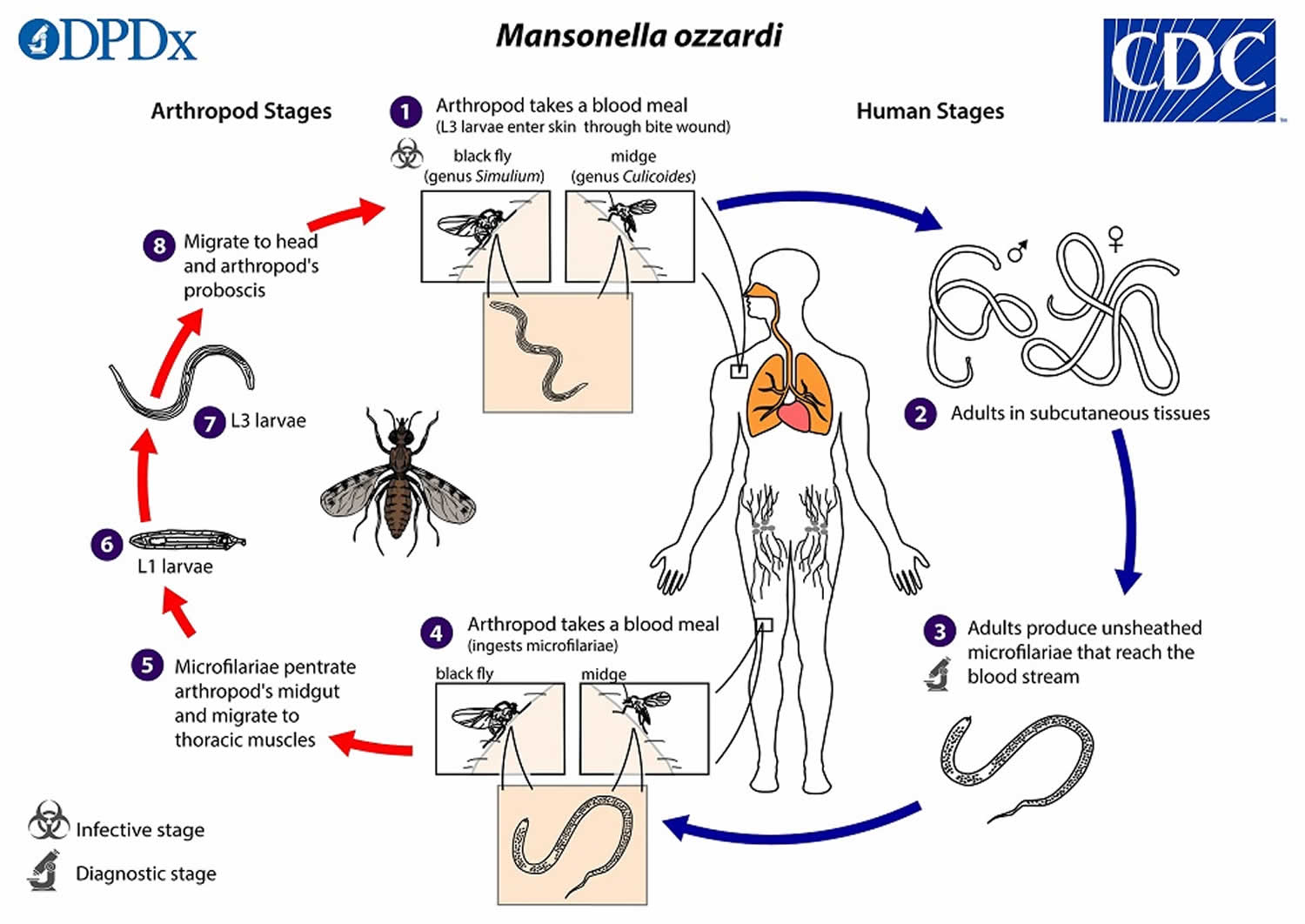
During a blood meal, an infected arthropod (midges, genus Culicoides, or blackflies, genus Simulium) introduces Mansonella ozzardi third-stage filarial larvae (L3 larvae) onto the skin of the human host, where they penetrate into the bite wound (number 1). The third-stage filarial larvae (L3 larvae) develop into adults that commonly reside in subcutaneous tissues (number 2). Mansonella ozzardi adult worms are small and slender, measuring about 49 mm long and 150 µm wide (females) or 26 mm long and 70 µm wide (males). Mansonella ozzardi adult worms produce unsheathed and non-periodic microfilariae (immature worms) that reach the blood stream (number 3). The arthropod (midges, genus Culicoides, or blackflies, genus Simulium) ingests microfilariae during a blood meal (number 4). After ingestion, the microfilariae migrate from the arthropod’s midgut through the hemocoel to the thoracic muscles (number 5). There the microfilariae develop into first-stage larvae (number 6) and subsequently into third-stage infective larvae (L3 larvae) (number 7). The third-stage infective larvae (L3 larvae) migrate to arthropod’s proboscis (number 8) and can infect another human when the arthropod takes a blood meal (number 1).
Figure 7. Mansonella ozzardi life cycle
Mansonella streptocerca life cycle
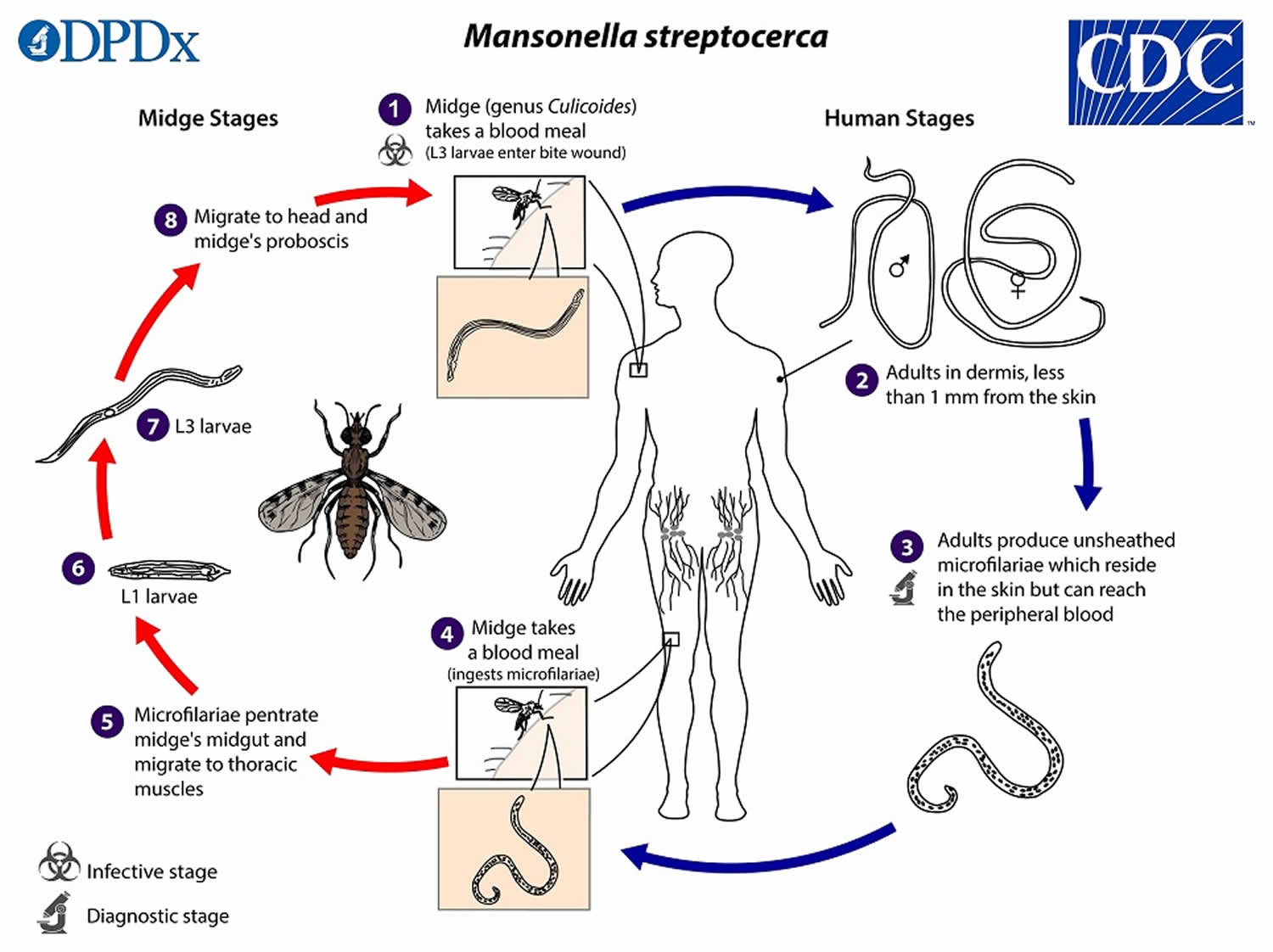
During a blood meal, an infected midge introduces Mansonella streptocerca infective third-stage filarial larvae (L3 larvae) onto the skin of the human host, where they penetrate into the bite wound (number 1). The infective third-stage filarial larvae (L3 larvae) develop into adults that reside in the dermis, most commonly less than 1 mm from the skin surface (number 2). The Mansonella streptocerca adult females measure approximately 27 mm in length, and are 50 µm wide at the level of the vulva (anteriorly) and ovaries (near the posterior end), and up to 85 µm at the mid-body. The Mansonella streptocerca adult males measure 50 µm in diameter. The Mansonella streptocerca adults produce unsheathed and non-periodic microfilariae (immature worms), measuring 180 to 240 µm by 3 to 5 µm, which reside in the skin but can also reach the peripheral blood (number 3). The midge vector ingests the microfilariae during a blood meal (number 4). After ingestion, the microfilariae migrate from the midge’s midgut through the hemocoel to the thoracic muscles (number 5). There the microfilariae develop into first-stage larvae (number 6) and subsequently into third-stage infective larvae (L3 larvae) (number 7). The third-stage infective larvae (l3 larvae) migrate to the midge’s proboscis (number 8) and can infect another human when the midge takes another blood meal (number 1).
Figure 8. Mansonella streptocerca life cycle
Mansonellosis Prevention
The best way to prevent mansonellosis is to avoid midge and black fly bites. If you’re in an area where mansonellosis is common use Environmental Protection Agency (EPA)-registered insect repellents containing DEET (diethyltoluamide or N,N-diethyl-meta-toluamide) or another EPA-registered active ingredient (https://www.epa.gov/insect-repellents/find-repellent-right-you). Wear loose-fitting, long-sleeved shirts and pants. Keep windows and doors closed or covered with screens to keep biting insects out. Repair broken screening on windows, doors, porches, and patios.
Mansonellosis signs and symptoms
Most people infected with the parasitic worms that cause mansonellosis never develop symptoms (asymptomatic) or can be associated with nonspecific clinical signs and symptoms such as fever, headache, joint pain (arthralgia), and eyelesions (Mansonella ozzardi); itch (pruritus), joint pain (arthralgia), abdominal pain, soft tissue swelling (angioedema), skin rash, and fatigue (Mansonella perstans and perhaps Mansonella ‘DEUX’); and itchy dermatitis and chronic lymph gland inflammation (Mansonella perstans) 50, 4.
Mansonellosis signs and symptoms typically include 18, 51, 52, 53:
- Occasional swelling
- Impaired vision, if worms enter your eye
- Itching
- Rashes
- Joint pain (arthralgia)
- Abdominal (belly) pain
- Enlargement of lymph nodes (lymphadenopathy)
- Pericarditis (inflammation of the outer lining of the heart)
- Fever
- Feeling weak and tired (fatigue)
Recent observations suggest that mansonellosis infections can influence the human immune system’s response and this can influence the development of secondary infections, like malaria 90. As mansonellosis infections are chronic and occur in many tropical areas where a great number of other infectious diseases also exist, it is possible that the way mansonellosis influences other pathogenic infections could actually account for a very substantial proportion of its pathogenicity and its total disease burden 11, 12, 71, 36, 68. At present, however, although there have been many accounts of mansonellosis parasites co-occurring with other parasites, there have been very few accounts about how mansonellosis influences the pathogenicity of other infectious diseases 91, 90, 92, 93, 94, 95.
Most symptoms ascribed to Mansonella perstans infections in modern scientific literature are based on symptoms that have been recorded in case study reports 6. As most of these reports are based on the treatment of tourists and expatriate Europeans and North Americans returning home from endemic areas, and not on people who have lived all their lives in endemic areas, it is not clear whether the symptoms reported from these studies can be used to compile a clinical picture that represents all or even most infections caused by mansonellosis 36, 68, 96, 97, 98. Based on these reports, Mansonella perstans can be considered to have little pathogenicity and almost always to be asymptomatic, but it can occasionally cause itching, joint pains, enlarged lymph glands, and vague abdominal symptoms 12, 36, 68, 96, 97, 98.
High-level eosinophilia are regularly observed in some but not all patients with Mansonella perstans infections, probably as a consequence of the body’s reaction against the adult worm, rather than against microfilariae 99. Mansonella perstans is also reported to be able to induce dermatological symptoms like the Calabar swellings of loiasis, fever, headaches, and pain in bursae and/or joint synovia or in serous cavities 36, 68, 96, 99. There has also been eye pathology ascribed to Mansonella perstans in which yellowish nodules develop in the bulbar conjunctiva; it has been reported that this pathology can on occasion provoke edema of the eyelids and proptosis 99, 100. This pathology, which has been described as a rare clinical manifestation of Mansonella perstans infections caused by adult Mansonella perstans migrating to the eye, is commonly referred to as “bulge” or “bung” eye disease 99, 100. Whether this pathology is caused by Mansonella perstans or a zoonotic parasite from the Mansonella genus is not presently clear 99, 100. It is not uncommon for zoonotic filarial infections to have ocular clinical manifestations of this type 100, 101.
In addition to showing that Mansonella ozzardi infections are associated with joint pain, leg chills, headaches, and corneal lesions, epidemiological studies have also added support to the notion that Mansonella ozzardi infections are mostly asymptomatic 102, 103, 104, 105, 106. Case reports from tourist-acquired infections have also suggested that Mansonella ozzardi infections can cause fatigue, respiratory problems, dermal swellings, and itching; however, evidence that such symptoms occur regularly in endemic settings is still lacking 33, 107.
There have been very few studies of Mansonella streptocerca infections. From what data are available, however, it is clear that infections with this parasite are very often (if not exclusively) asymptomatic 6. However, it has been proposed that infections cause various types of dermatological symptoms in the shoulders and thorax, where Mansonella streptocerca parasite is mostly encountered 11, 12, 35, 68, 72, 38. Hyperpigmented macules have been attributed to infections occurring in the Democratic Republic of the Congo, but have not been reported everywhere the parasite is found, including Uganda, and thus some authors have cautioned against their attribution to Mansonella streptocerca infections 35, 72, 38. Chronic papular dermatitis has been encountered in most Mansonella streptocerca-endemic areas and has been reported to be cleared from patients successfully treated for Mansonella streptocerca infections 72. However, in endemic areas where epidemiological studies have been performed, chronic papular dermatitis is no more common in those patients who have detectable Mansonella streptocerca in their skin snips than in those that do not, and thus at present there is no statistically robust evidence to support the parasites causing any symptoms at all 6.
Mansonella perstans infection signs and symptoms
In general, Mansonella perstans infections lack a distinct and specific signs and symptoms compared to other human filarial infections 3. This ambiguity contributes to the significant neglect and underrecognition of mansonellosis as a public health concern, despite its high prevalence. Nonetheless, clinical reports have described a range of symptoms associated with Mansonella perstans infection, including transient upper abdominal pain, allergic reactions, eosinophilia, subcutaneous swellings (resembling the ‘Calabar swellings’ seen in Loa loa), aches in bursae or joint synovia, discomfort in serous cavities and the liver region, neurological and psychological symptoms, profound fatigue, oedema, and hormonal disturbances 108, 18, 109, 110, 111, 112.
In 1962, Adolph et al. proposed a classification of clinical manifestations associated with Mansonella perstans infection into seven categories 109, 110:
- Acute swelling of the forearms, hands, and/or face, typically resolving within 3 to 4 days, often recurrent, and resembling Calabar swelling;
- Itch (pruritus), with or without accompanying rash or ulceration;
- Pain or discomfort in bursae and/or joint synovia;
- Pain in serous cavities;
- Discomfort or pain in the liver region (right upper quadrant of abdomen);
- Neurological and psychological symptoms; and
- Profound fatigue or exhaustion.
These signs and symptoms of Mansonella perstans infection are believed to result from worm migration and the resulting host immune responses. However, the majority of Mansonella perstans infected individuals remain asymptomatic, underscoring the immunomodulatory potential and evolutionary adaptation of Mansonella perstans.
Several studies have shown that Mansonella perstans infection is associated with eosinophilia and elevated total IgE levels 113, 112, 114, 115, which may explain symptoms such as itch and swelling. However, due to the historical lack of available Mansonella perstans worm material, these studies could not assess parasite-specific antibody responses. Additionally, many findings on the immunomodulatory capacity of Mansonella perstans have come from studies involving individuals with coinfections from other parasites 116, 117, 118, 119, 120, casting doubt on whether these immune alterations are specific to Mansonella perstans.
More recently, a study conducted on individuals with Mansonella perstans mono-infections, using worm-extract antigen, allowed for a detailed analysis of antigen-specific immune responses 121. In addition to increased total and antigen-specific IgE, the study found elevated levels of antigen-specific IgG4, decreased systemic concentrations of cytokines (IL-4, IL-6, IL-12p70) and chemokines (IL-8, RANTES), and increased production of IFN-γ, IL-10, and IL-13 upon antigen stimulation 121. Moreover, there were increased frequencies of Th2 cells, regulatory B cells, and regulatory T cells, showing that Mansonella perstans modulates systemic cytokine, chemokine, and immunoglobulin responses via specific regulatory immune cell populations 121. This targeted modulation of innate and adaptive immune responses likely aids the parasite in evading host immunity.
Immunomodulation may also affect the course of coinfections and reduce the efficacy of vaccines. Several studies have reported associations between Mansonella perstans infection and increased susceptibility or more severe disease outcomes in HIV, tuberculosis (TB), Buruli ulcer, and malaria 18, 122, 123, 124. These findings suggest that, although often asymptomatic, Mansonella perstans infections may still warrant treatment to minimise risks associated with coinfections.
Additionally, Mansonella perstans has been implicated in modulating vaccine efficacy. For instance, its presence impairs the protective response to Bacillus Calmette–Guérin (BCG) vaccination against TB. Furthermore, maternal Mansonella perstans infection during pregnancy can influence the infant’s immune response to vaccines, potentially through transplacental transfer of immunomodulatory factors 125.
Conversely, this immunomodulatory effect might also have beneficial side effects, particularly in the context of autoimmune, metabolic, or allergic diseases. Helminth-driven immune regulation has been shown to confer protection against conditions such as asthma and diabetes 126, 127. One study found that women with a history of asthma were less likely to be infected with Mansonella perstans, and among those who were infected, the likelihood of atopic sensitization was significantly lower 114. An ongoing study is investigating whether Mansonella perstans infection may influence metabolic disease and overall metabolic health 128. and preliminary data indicate that Mansonella perstans infection is associated with a decreased prevalence of diabetes and an improved liver enzyme profile in Cameroon.
Mansonellosis diagnosis
Mansonella perstans and Mansonella ozzardi are usually diagnosed by the finding of microfilariae (immature worms) circulating in your blood whereas Mansonella streptocerca is usually diagnosed by finding microfilariae in skin snips 54. The rare human cases of Mansonella rodhaini were also diagnosed via skin snips. Concentration techniques (e.g. Knott’s concentration, polycarbonate membrane filtration) may increase sensitivity. Note that these Mansonella parasitic worms do not generally exhibit periodicity, though in some regions, Mansonella perstans-infected patients may exhibit slightly elevated microfilaremia in morning hours. Skin snips should be thick enough to include the outer part of the dermal papillae but not so thick as to produce bleeding (a ~2 mm wide section should be sufficient) 54. Skin snips should be placed immediately in normal saline or distilled water, just enough to cover the specimen 54. Microfilariae tend to emerge more rapidly in saline, however in either medium the microfilariae typically emerge in 30—60 minute and can be seen in wet mount preparations 54. For a definitive diagnosis, allow the wet mount to dry, fix in methanol, and stain with Giemsa or hematoxylin-and-eosin (H&E) 54. Serological assays developed for the detection of Mansonella species–specific antibodies or antigens are generally of limited utility due to cross-reactions with other filarial species and potentially other roundworms (nematodes) 54.
Immunological tests using Dirofilaria immitis antigens have been explored for diagnosing Mansonella perstans, but their low sensitivity causes false negatives, while cross-reactivity with other filarial infections leads to false positives 18. Another diagnostic approach is antibody detection, usually targeting IgG4, which is elevated in microfilariae-positive individuals 6. However, no validated antigen or antibody tests currently exist for immunodiagnosis or surveillance, and antibody detection cannot differentiate past exposure from active infection.
Molecular techniques such as PCR (polymerase chain reaction) and sequencing offer a sensitive alternative to microscopy, capable of detecting all human filarial parasites. Species-specific PCR enhances sensitivity and enables identification of co-endemic species. A key advancement was the nested PCR assay targeting the internal transcribed spacer 1 (ITS1) region of ribosomal DNA, detecting all filarial life stages in humans, vectors, and animal reservoirs, with species differentiation based on amplicon size 129. Earlier, PCR-restriction fragment length polymorphism (RFLP) combined semi-nested PCR with enzyme digestion for species identification via fragment size 130. More recent methods target repetitive sequences such as Mansonella perstans repeat 1 (MpR1), allowing highly sensitive detection of Mansonella perstans DNA 131.
Another molecular approach, CO1 barcoding, uses a conserved region of the cytochrome c oxidase subunit I (CO1) gene to differentiate species. Its rapid evolution allows discrimination between closely related filariae. CO1 analysis shows that Mansonella perstans is closely related to Mansonella ozzardi, both within the ONC5 clade, and is more closely related to Wuchereria bancrofti and Brugia malayi than to Loa loa 132. These findings have been confirmed by whole-genome and mitochondrial genome sequencing, which place Mansonella perstans and Mansonella sp. ‘DEUX’ within the ONC5 clade 133, 23.
Despite their sensitivity, PCR (polymerase chain reaction) and sequencing require expensive equipment and reagents, limiting their use in resource-poor, endemic areas. As an alternative, loop-mediated isothermal amplification (LAMP) operates at a constant temperature without thermal cyclers and allows real-time detection via colorimetric or fluorescent signals. The LAMP (loop-mediated isothermal amplification) test is a form of testing that involves replicating or amplifying small fragments of genetic material at a constant temperature. The real-time loop-mediated amplification method that LAMP uses to amplify the genetic material in a sample makes it faster and more cost effective than a PCR (polymerase chain reaction) test. In the LAMP test, the fragments of DNA are amplified at one temperature only, which speeds up the reaction time. This method of amplifying DNA at one temperature is what sets the LAMP test apart from the more conventional PCR test, which uses temperature cycling for DNA replication. LAMP has been applied to detect Onchocerca volvulus, Brugia malayi and Wuchereria bancrofti 134, and more recently to distinguish Mansonella perstans and Mansonella ozzardi 134, 135. While promising for field use, further evaluation of LAMP’s performance and feasibility is still needed, especially for validation of the diagnosis of microfilariae-negative filarial infections.
Parasitological diagnosis
Traditional parasitological diagnosis, which is based on direct observation of a causative parasite, is still the most common way mansonellosis infections are diagnosed 6. It is performed by the detection and identification of sheathless Mansonella microfilariae in the skin or blood at any time of day or night 6. Detection of skin-dwelling microfilariae is usually done by examination of ~1.5 mg (~2 mm diameter) skin-snip (skin-biopsy) samples, taken with a Walser or Holth corneoscleral punch 6. The skin-snip test requires that a skin biopsy be taken from areas of presumed optimal microfilariae density, and this varies depending on geographic location. Blood samples used for mansonellosis diagnosis are often taken from peripheral blood samples by way of finger pricks, although venous blood samples can also be used for this purpose 6.
When microfilariae are present in high numbers in the patient’s blood, as is often the case in endemic areas, they may be easily found and identified in thick or thin blood smears stained with Giemsa or hematoxylin 6. Skin snips are usually incubated in water or saline, and the emergent microfilariae can be stained on microscope slides, but in fresh unstained wet preparations of skin snips, the way in which the microfilariae move can also help with their identification. The main diagnostic microfilariae characteristics can be found in Table 1 above. Morphologically, the most important features to identify microfilariae in blood smears stained with Giemsa are size and shape of the tail, presence or absence of a sheath, and the arrangement of terminal nuclei (in the tail) 6. All Mansonella microfilariae are unsheathed, and for diagnostic purposes are treated as aperiodic 6.
The microfilariae of Mansonella perstans are reported to be 200×4–5 μm, to have blunt rounded tails, and to have nuclei extending to the tips of their tails (Figure 1). Mansonella perstans is easily distinguishable from other blood-dwelling microfilariae, which have overlapping distributions (Loa loa or Wuchereria bancrofti), by their being smaller, their lack of an enveloping sheath, and their tail features (their terminal nuclei are bigger than the other microfilariae) 6.
Mansonella streptocerca microfilariae are smaller and thinner (180–240×3–5 μm) than Onchocerca volvulus, and have hook-shaped tails with nuclei that extend to the end (no nuclei are seen in the pointed tail region of Onchocerca volvulus microfilariae) (Figure 3). In wet mounts, live Mansonella streptocerca microfilariae are usually less motile than those of Onchocerca volvulus 12, 136. With these morphological descriptions, the two filariae should be easily distinguished. In the past, however, in some countries, such as Uganda, Mansonella streptocerca and Onchocerca volvulus have been confused, despite the clear morphological differences between them 137, 138, 72.
The microfilariae of Mansonella ozzardi are reported to be 163–203×3–5 μm, with long thin pointed tails and body nuclei that do not extend to the tips of their tails 6. As they can locate in both the blood and skin, from a public health perspective it is most important that the microfilariae can be differentiated from Wuchereria bancrofti and Onchocerca volvulus 6. The microfilariae of Mansonella ozzardi can be easily distinguished from those of Wuchereria bancrofti by virtue of being smaller and having no sheath (Figure 2) 6. Living microfilariae of Mansonella ozzardi can be obtained from skin snips and are not easily differentiated from Onchocerca volvulus by movement 6. Their morphology is similar to the pathogenic Onchocerca volvulus, and it is especially important to distinguish these two species where they are sympatric, eg, in Amazonia onchocerciasis focus. Although morphological characters can be used to discriminate these two species 139, difficulties have led to false reports of new onchocerciasis foci and continue to pose a problem for onchocerciasis epidemiological monitoring in the Amazonia focus, which is now the last Latin America onchocerciasis focus where transmission is considered to be ongoing 6.
Immunodiagnosis
Immunological methods involve detection of either antibody or antigen. When a filarial parasite immunodiagnostic assay is detecting antibodies, enhanced specificity is often achieved by assessment of IgG4 rather than total IgG, as IgG4 antibodies are significantly elevated in microfilaria-positive individuals 140. Although there is presently no effective antigen- or antibody-detecting immunological assay for diagnosis or epidemiological monitoring of mansonellosis infections, both kinds of tools have been developed for onchocerciasis and lymphatic filariasis disease-control programs, and have been tested for cross-reactivity with mansonellosis sera 6. The reliability (specificity and sensitivity) of these immunodiagnostic tools for lymphatic filariasis and onchocerciasis has not been fully characterized for all of the tests, and the best-understood and most reliable tests are not necessarily the most convenient, and thus whether they should be employed to assist with mansonellosis research and diagnostics is questionable 6. However, two recently published Mansonella perstans studies used generic immunological filariasis assays to support light-microscopy-based diagnoses 97.
Molecular diagnosis
Although DNA-based diagnostic tools can be used to detect and identify all of the known human filarial parasites, their most important utility to public health is their ability to differentiate skin infections of Mansonella streptocerca and Mansonella ozzardi from Onchocerca volvulus skin infections and Mansonella perstans parasitemias from Loa loa and Wuchereria bancrofti parasitemias 141, 142. DNA-based techniques for filarial parasite detection and identification have been proven to be both sensitive and specific, and have now begun to replace light-based microscopy in epidemiological surveys of ansonellosis 6. Molecular diagnosis may be used to detect microfilariae in both peripheral blood and skin biopsies, and adult worms in other tissues 6.
PCR-based amplification of species-specific target sequences allows increasing diagnostic sensitivity compared with microscopic methods and reliable differentiation of samples taken from individuals living in coendemic areas 6. In 2010, Tang et al 143 developed a popular nested PCR that could detect any form and life stage of filariae in the human host or vector. This assay uses universal filariae PCR primers to amplify a variable portion of filarial parasite ribosomal ITS1 DNA, and allows for the subsequent identification of species based on the size of the amplified fragment. This technique is used in conjunction with gel electrophoresis and/or Sanger sequencing, and allows for the characterization of previously unknown species (the Mansonella perstans variant Deux was found in this way) 6. Recently, this rDNA ITS1-based method was turned into a single-step diagnostic by adapting it for real-time PCR.113,131 This new assay allows for the identification of filarial parasite without the need for gel electrophoresis or Sanger sequencing, but does require more expensive reagents and infrastructure than standard PCR assays, and does not directly allow for the characterization of novel or unexpected filarial species 94, 144. Other methods such as PCR–restriction-fragment-length polymorphism (RFLP) that do not allow for the characterization of novel or unexpected filarial species, but require less infrastructure to support them and can differentiate a broad range of filarial species using universal primers with a combination of PCR and RFLP, have also been reported 145, 130. Despite requiring less complex infrastructural support than alternative methods, PCR-RFLP assays still require access to PCR reagents and a PCR machine, which can be a limiting factor for filarial parasite epidemiological studies.
Recently, the first DNA-detecting loop-mediated isothermal amplification (LAMP) filarial parasite assays were reported 146, 147. The LAMP (loop-mediated isothermal amplification) test is a form of testing that involves replicating or amplifying small fragments of genetic material at a constant temperature. The real-time loop-mediated amplification method that LAMP uses to amplify the genetic material in a sample makes it faster and more cost effective than a PCR (polymerase chain reaction) test. In the LAMP test, the fragments of DNA are amplified at one temperature only, which speeds up the reaction time. This method of amplifying DNA at one temperature is what sets the LAMP test apart from the more conventional PCR test, which uses temperature cycling for DNA replication. LAMP has been applied to detect Onchocerca volvulus, Brugia malayi and Wuchereria bancrofti 134, and more recently to distinguish Mansonella perstans and Mansonella ozzardi 134, 135. While promising for field use, further evaluation of LAMP’s performance and feasibility is still needed, especially for validation of the diagnosis of microfilariae-negative filarial infections.
Mansonellosis treatment
Due to the largely asymptomatic clinical presentation of Mansonella infection, most people do not require treatment for mansonellosis 53. However, some experts recommend that human infections with Mansonella species be treated, regardless of any apparent clinical manifestations 4. If you are a diagnosed with mansonellosis, your doctor will discuss treatment options, if needed, with you 53. There are currently no specific treatment guidelines or approved medications targeting Mansonella parasite 3. Only a limited number of studies have evaluated the efficacy of drugs against Mansonella perstans, and these investigations often involve small groups, making it difficult to draw robust conclusions. Furthermore, many of these studies have been conducted on travellers or expatriates returning to Europe or North America from endemic regions 6, raising concerns about the generalizability of findings due to potential immunological differences between individuals from endemic and non-endemic areas.
The available data on anthelminth and antibiotic treatments are limited and often contradictory 3. Single-dose regimens of commonly used anthelmintics such as albendazole, praziquantel, and ivermectin, used in mass drug administration (MDA) for onchocerciasis, lymphatic filariasis, or schistosomiasis, have shown minimal or no impact on microfilarial loads in infected individuals 18, 55, 56. In contrast, multiple-dose regimens using drugs such as thiabendazole, mebendazole, diethylcarbamazine (DEC), or a combination of mebendazole and diethylcarbamazine (DEC), have demonstrated greater efficacy 57. The mebendazole–diethylcarbamazine (DEC) combination achieved the highest clearance rate, with a reported 37% reduction in microfilariae 56. Among the most promising treatments is the antibiotic doxycycline, which targets Wolbachia endosymbionts. Doxycycline was shown to clear Mansonella perstans microfilariae from the peripheral blood for the observational period of up to 2 years 6, 44, 45, indicating a filarial sterilizing and/or macrofilaricidal effect. However, its requirement for a 6-week treatment course poses logistical challenges for use in large-scale control programmes.
New macrofilaricidal drugs allowing shorter treatment regimens, such as oxfendazole, are currently under investigation 3. Oxfendazole is a pan-nematode candidate which has been used for several decades as an anthelmintic in pets 3. Given oxfendazole’s proven efficacy in filarial animal models following a 5-day treatment, and its selective macrofilaricidal efficacy with no strong direct microfilaricidal efficacy 58, it is expected that oxfendazole does not cause microfilariae-induced adverse events in microfilaraemic individuals. An EDCTP3-funded Phase 2 clinical trial (PACTR202412611774752) is ongoing in Cameroon, Gabon, the Democratic Republic of the Congo, and Tanzania to evaluate oxfendazole’s efficacy against Mansonella perstans, Loa loa, Onchocerca volvulus, as well as trichuriasis.
Novel anti-Wolbachial macrofilaricidal candidates such as Corallopyronin A, which is expected to allow treatment regimens of 14 days or less 59, 60, and is currently under preparation for Phase 1 clinical trials in humans 61, are expected to be efficacious for all Wolbachia-containing filariae, that is, filariae causing onchocerciasis, lymphatic filariasis, and mansonellosis. Moreover, combinations of anti-Wolbachials with oxfendazole or albendazole are suggested to provide synergistic effects and may allow shorter treatment regimens and improve drug efficacy 62.
Mansonellosis prognosis
The prognosis for mansonellosis is generally good as most infections are asymptomatic and considered benign, though some cases can involve mild symptoms like itching, swelling, leg-chills, joint pains, headaches, corneal lesions and fevers, or even contribute to severe complications with co-infections 4, 148.
- Simonsen PE, Onapa AW, Asio SM. Mansonella perstans filariasis in Africa. Acta Trop. 2011 Sep;120 Suppl 1:S109-20. https://doi.org/10.1016/j.actatropica.2010.01.014[↩][↩]
- Ta-Tang, T. H., Luz, S. L. B., Crainey, J. L., & Rubio, J. M. (2021). An Overview of the Management of Mansonellosis. Research and Reports in Tropical Medicine, 12, 93–105. https://doi.org/10.2147/RRTM.S274684[↩][↩]
- Ritter M, Hsu HY, Lenz B, Kien CA, Gandjui NVT, Hübner MP, Hoerauf A, Wanji S. Mansonella perstans – the forgotten filaria. Trends Parasitol. 2025 Sep 3:S1471-4922(25)00225-9. https://www.cell.com/trends/parasitology/fulltext/S1471-4922(25)00225-9[↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩]
- Ferreira, M. U., Crainey, J. L., & Gobbi, F. G. (2023). The search for better treatment strategies for mansonellosis: an expert perspective. Expert Opinion on Pharmacotherapy, 24(15), 1685–1692. https://doi.org/10.1080/14656566.2023.2240235[↩][↩][↩][↩][↩][↩][↩]
- Bain O, Mutafchiev Y, Junker K, Guerrero R, Martin C, Lefoulon E, Uni S. Review of the genus Mansonella Faust, 1929 sensu lato (Nematoda: Onchocercidae), with descriptions of a new subgenus and a new subspecies. Zootaxa. 2015 Feb 11;3918(2):151-93. doi: 10.11646/zootaxa.3918.2.1[↩][↩]
- Ta-Tang TH, Crainey JL, Post RJ, Luz SL, Rubio JM. Mansonellosis: current perspectives. Res Rep Trop Med. 2018 Jan 18;9:9-24. doi: 10.2147/RRTM.S125750[↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩]
- Debrah, L.B., Arthur, J.F., Yeboah, A. et al. Diversity of Culicoides in the middle belt of Ghana with Implications on the transmission of Mansonella perstans; a molecular approach. Parasites Vectors 17, 123 (2024). https://doi.org/10.1186/s13071-024-06179-8[↩][↩]
- Ebai R, Kien CA, Fombad FF, Esofi F, Ouam E, Ntuh AN, Amambo GN, Gandjui VNT, Chunda VC, Nietcho F, Nchang LC, Magha C, Cho JF, Esum ME, Enyong PI, Pfarr K, Hoerauf A, Ritter M, Wanji S. Culicoides Species of the Rain Forest Belt of the Littoral Region of Cameroon: Their Incrimination in the Transmission of Mansonella perstans. Pathogens. 2024 Feb 6;13(2):146. doi: 10.3390/pathogens13020146[↩][↩]
- Downes BL, Jacobsen KH. A systematic review of the epidemiology of mansonelliasis. Afr J Infect Dis. 2010;4(1):7–14. doi: 10.4314/ajid.v4i1.55085[↩][↩]
- Lima NF, Aybar CA, Juri MJ, Ferreira MU. Mansonella ozzardi: a neglected New World filarial nematode. Pathog Glob Health. 2016;110(3):97–107. doi: 10.1080/20477724.2016.1190544[↩][↩][↩]
- Medeiros JF, Crainey JL, Pessoa FA, Luz SL. Mansonelliasis. In: Marcondes CB, editor. Arthropod Borne Diseases. Heidelberg: Springer; 2017. pp. 405–426.[↩][↩][↩][↩][↩][↩][↩][↩][↩]
- Nutman TB. Filarial infections. In: Cohen J, Powderly WG, Opal SM, editors. Infectious Diseases. Amsterdam: Elsevier; 2016. pp. 1046–1052.[↩][↩][↩][↩][↩][↩]
- Shelley AJ, Coscarón S. Simuliid blackflies (Diptera: Simuliidae) and ceratopogonid midges (Diptera: Ceratopogonidae) as vectors of Mansonella ozzardi (Nematoda: Onchocercidae) in northern Argentina. Mem Inst Oswaldo Cruz. 2001 May;96(4):451-8. doi: 10.1590/s0074-02762001000400003[↩][↩]
- Asio SM, Simonsen PE, Onapa AW. Mansonella perstans filariasis in Uganda: patterns of microfilaraemia and clinical manifestations in two endemic communities. Trans R Soc Trop Med Hyg. 2009 Mar;103(3):266-73. doi: 10.1016/j.trstmh.2008.08.007[↩][↩]
- Santa Maria Asio, Paul E. Simonsen, Ambrose W. Onapa, Mansonella perstans filariasis in Uganda: patterns of microfilaraemia and clinical manifestations in two endemic communities, Transactions of The Royal Society of Tropical Medicine and Hygiene, Volume 103, Issue 3, March 2009, Pages 266–273, https://doi.org/10.1016/j.trstmh.2008.08.007[↩][↩]
- Portela CS, Mendes de Araújo CP, Moura Sousa P, Gomes Simão CL, Silva de Oliveira JC, Crainey JL. Filarial disease in the Brazilian Amazon and emerging opportunities for treatment and control. Curr Res Parasitol Vector Borne Dis. 2023 Dec 23;5:100168. doi: 10.1016/j.crpvbd.2023.100168[↩]
- Kamtchum Tatuene J.G.F.R., Nkoa T., Tchateng Mbougua J.B., Nana Djeunga H.C., Bopda J. and Kamgno J., Epidemiology of Loa loa and Mansonella perstans filariasis in the Akonolinga health district, centre region, Cameroon. Health Sci Dis, 2014. 15(1): p. 1.[↩]
- Simonsen PE, Onapa AW, Asio SM. Mansonella perstans filariasis in Africa. Acta Trop. 2011 Sep;120 Suppl 1:S109-20. doi: 10.1016/j.actatropica.2010.01.014[↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩]
- HOPKINS CA. Notes on the biology of certain Culicoides studied in the British Cameroons, West Africa, together with observations on their possible rôle as vectors of Acanthocheilonema perstans. Ann Trop Med Parasitol. 1952 Sep;46(2):165-72. doi: 10.1080/00034983.1952.11685521[↩][↩]
- Linley JR, Hoch AL, Pinheiro FP. Biting midges (Diptera: Ceratopogonidae) and human health. J Med Entomol. 1983 Jul 21;20(4):347-64. doi: 10.1093/jmedent/20.4.347[↩][↩][↩][↩]
- Ebai R, Kien CA, Fombad FF, et al. Culicoides Species of the Rain Forest Belt of the Littoral Region of Cameroon: Their Incrimination in the Transmission of Mansonella perstans. Pathogens. 2024 Feb 6;13(2):146. doi: 10.3390/pathogens13020146[↩][↩]
- Mourembou G, Fenollar F, Lekana-Douki JB, Ndjoyi Mbiguino A, Maghendji Nzondo S, Matsiegui PB, Zoleko Manego R, Ehounoud CH, Bittar F, Raoult D, Mediannikov O. Mansonella, including a Potential New Species, as Common Parasites in Children in Gabon. PLoS Negl Trop Dis. 2015 Oct 20;9(10):e0004155. doi: 10.1371/journal.pntd.0004155[↩][↩][↩][↩][↩][↩]
- Rodi M, Gross C, Sandri TL, Berner L, Marcet-Houben M, Kocak E, Pogoda M, Casadei N, Köhler C, Kreidenweiss A, Agnandji ST, Gabaldón T, Ossowski S, Held J. Whole genome analysis of two sympatric human Mansonella: Mansonella perstans and Mansonella sp “DEUX”. Front Cell Infect Microbiol. 2023 Apr 14;13:1159814. doi: 10.3389/fcimb.2023.1159814[↩][↩][↩][↩][↩]
- Sandri TL, Kreidenweiss A, Cavallo S, Weber D, Juhas S, Rodi M, Woldearegai TG, Gmeiner M, Veletzky L, Ramharter M, Tazemda-Kuitsouc GB, Matsiegui PB, Mordmüller B, Held J. Molecular Epidemiology of Mansonella Species in Gabon. J Infect Dis. 2021 Feb 3;223(2):287-296. doi: 10.1093/infdis/jiaa670. Erratum in: J Infect Dis. 2022 Mar 2;225(5):929. doi: 10.1093/infdis/jiab439[↩][↩][↩][↩][↩]
- Raccurt CP. Mansonella ozzardi and its vectors in the New World: an update with emphasis on the current situation in Haiti. J Helminthol. 2017 Oct 25; doi: 10.1017/S0022149X17000955[↩]
- Nelson GS. Filarial infections as zoonoses. J Helminthol. 1965;39(2):229–250. doi: 10.1017/s0022149x00020630[↩]
- Neafie R, Meyers W, Connor DJ. Mansonelliasis. In: Binford C, Connor D, editors. Pathology of Tropical and Extraordinary Diseases. Washington: Armed Forces Institute; 1976. p. 390.[↩]
- Shelley AJ, Dias AP, Moraes MA. Simulium species of the amazonicum group as vectors of Mansonella ozzardi in the Brazilian Amazon. Trans R Soc Trop Med Hyg. 1980;74(6):784–788. doi: 10.1016/0035-9203(80)90200-x[↩]
- Linley JR, Hoch AL, Pinheiro FP. Biting midges (Diptera: Ceratopogonidae) and human health. J Med Entomol. 1983;20(4):347–364. doi: 10.1093/jmedent/20.4.347[↩][↩]
- Marinkelle CJ, German E. Mansonelliasis in the Comisaría del Vaupes of Colombia. Trop Geogr Med. 1970 Mar;22(1):101-11.[↩]
- Orihel TC, Eberhard ML. Mansonella ozzardi: a redescription with comments on its taxonomic relationships. Am J Trop Med Hyg. 1982;31(6):1142–1147. doi: 10.4269/ajtmh.1982.31.1142[↩][↩]
- Eberhard ML, Orihel TC. The genus Mansonella (syn. Tetrapetalonema): a new classification. Ann Parasitol Hum Comp. 1984;59(5):483–496. doi: 10.1051/parasite/1984595483[↩]
- Nutman TB, Nash TE, Ottesen EA. Ivermectin in the successful treatment of a patient with Mansonella ozzardi infection. J Infect Dis. 1987;156(4):662–665. doi: 10.1093/infdis/156.4.662[↩][↩]
- Kozek WJ, D’Alessandro A, Silva J, Navarette SN. Filariasis in Colombia: prevalence of mansonellosis in the teenage and adult population of the Colombian bank of the Amazon, Comisaria del Amazonas. Am J Trop Med Hyg. 1982;31(6):1131–1136. doi: 10.4269/ajtmh.1982.31.1131[↩]
- Duke BO. A case of streptocerciasis in a European. Ann Trop Med Parasitol. 1957;51(4):364–367. doi: 10.1080/00034983.1957.11685825[↩][↩][↩]
- Simonsen PE, Onapa AW, Asio SM. Mansonella perstans filariasis in Africa. Acta Trop. 2011;120(Suppl 1):S109–S120. doi: 10.1016/j.actatropica.2010.01.014[↩][↩][↩][↩][↩][↩][↩]
- Bassene H, Sambou M, Fenollar F, et al. High prevalence of Mansonella perstans filariasis in rural Senegal. Am J Trop Med Hyg. 2015;93(3):601–606. doi: 10.4269/ajtmh.15-0051[↩]
- Meyers WM, Connor DH, Harman LE, Fleshman K, Moris R, Neafie RC. Human streptocerciasis. A clinico-pathologic study of 40 Africans (Zairians) including identification of the adult filaria. Am J Trop Med Hyg. 1972 Sep;21(5):528-45.[↩][↩][↩][↩]
- Ferri E., Bain O., Barbuto M., Martin C., Lo N., Uni S., et al. (2011). New insights into the evolution of wolbachia infections in filarial nematodes inferred from a large range of screened species. PloS One 6 (6), e20843. doi: 10.1371/journal.pone.0020843[↩]
- Taylor MJ, Hoerauf A. Wolbachia bacteria of filarial nematodes. Parasitol Today. 1999 Nov;15(11):437-42. doi: 10.1016/s0169-4758(99)01533-1[↩][↩]
- McGarry HF, Pfarr K, Egerton G, Hoerauf A, Akue JP, Enyong P, Wanji S, Kläger SL, Bianco AE, Beeching NJ, Taylor MJ. Evidence against Wolbachia symbiosis in Loa loa. Filaria J. 2003 May 2;2(1):9. doi: 10.1186/1475-2883-2-9[↩][↩]
- Büttner DW, Wanji S, Bazzocchi C, Bain O, Fischer P. Obligatory symbiotic Wolbachia endobacteria are absent from Loa loa. Filaria J. 2003 May 9;2(1):10. doi: 10.1186/1475-2883-2-10[↩][↩][↩][↩]
- Grobusch MP, Kombila M, Autenrieth I, Mehlhorn H, Kremsner PG. No evidence of Wolbachia endosymbiosis with Loa loa and Mansonella perstans. Parasitol Res. 2003 Aug;90(5):405-8. doi: 10.1007/s00436-003-0872-z[↩][↩]
- Batsa Debrah L, Phillips RO, Pfarr K, Klarmann-Schulz U, Opoku VS, Nausch N, Owusu W, Mubarik Y, Sander AL, Lämmer C, Ritter M, Layland LE, Jacobsen M, Debrah AY, Hoerauf A. The Efficacy of Doxycycline Treatment on Mansonella perstans Infection: An Open-Label, Randomized Trial in Ghana. Am J Trop Med Hyg. 2019 Jul;101(1):84-92. doi: 10.4269/ajtmh.18-0491[↩][↩][↩][↩]
- Coulibaly YI, Dembele B, Diallo AA, Lipner EM, Doumbia SS, Coulibaly SY, Konate S, Diallo DA, Yalcouye D, Kubofcik J, Doumbo OK, Traore AK, Keita AD, Fay MP, Traore SF, Nutman TB, Klion AD. A randomized trial of doxycycline for Mansonella perstans infection. N Engl J Med. 2009 Oct 8;361(15):1448-58. doi: 10.1056/NEJMoa0900863[↩][↩][↩][↩]
- Pomari E, Voronin D, Alvarez-Martinez MJ, Arsuaga M, Bottieau E, Luzón-García MP, Nickel B, Rubio JM, Salas-Coronas J, Salvador F, Soriano-Pérez MJ, Sulleiro E, Goterris L, Van Esbroeck M, van Hellemond JJ, Wammes LJ, Zammarchi L, Lavezzari D, Degani M, Tais S, Held J, Gobbi F, Tamarozzi F. Wolbachia bacteria in Mansonella perstans isolates from patients infected in different geographical areas: a pilot study from the ESCMID Study Group for Clinical Parasitology. Parasit Vectors. 2025 Mar 10;18(1):97. doi: 10.1186/s13071-025-06723-0[↩][↩]
- Gehringer C, Kreidenweiss A, Flamen A, Antony JS, Grobusch MP, Bélard S. Molecular evidence of Wolbachia endosymbiosis in Mansonella perstans in Gabon, Central Africa. J Infect Dis. 2014 Nov 15;210(10):1633-8. doi: 10.1093/infdis/jiu320[↩][↩]
- Keiser PB, Coulibaly Y, Kubofcik J, Diallo AA, Klion AD, Traoré SF, Nutman TB. Molecular identification of Wolbachia from the filarial nematode Mansonella perstans. Mol Biochem Parasitol. 2008 Aug;160(2):123-8. doi: 10.1016/j.molbiopara.2008.04.012[↩][↩]
- Bordenstein S. R., Paraskevopoulos C., Dunning Hotopp J. C., Sapountzis P., Lo N., Bandi C., et al. (2009). Parasitism and mutualism in wolbachia: what the phylogenomic trees can and cannot say. Mol. Biol. Evol. 26 (1), 231–241. doi: 10.1093/molbev/msn243[↩]
- Sharp, N.A.D. Filaria perstans; its development in Culicoides austeni. Trans. R. Soc. Trop. Med. Hyg. 1928; 21:371-396[↩][↩]
- Brown, M. Manson’s tropical diseases. Lancet Infect. Dis. 2009; 9:407-408.[↩][↩]
- Adolph, P.E., KAGAN IG, McQUAY RM. Diagnosis and treatment of Acanthocheilonema perstans filariasis. Am. J. Trop. Med. Hyg. 1962; 11:76-88. doi: 10.4269/ajtmh.1962.11.76[↩][↩]
- About Mansonellosis. https://www.cdc.gov/filarial-worms/about/mansonellosis.html[↩][↩][↩][↩][↩][↩]
- Mansonellosis. https://www.cdc.gov/dpdx/mansonellosis[↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩]
- Wanji S, Tayong DB, Layland LE, Datchoua Poutcheu FR, Ndongmo WP, Kengne-Ouafo JA, Ritter M, Amvongo-Adjia N, Fombad FF, Njeshi CN, Nkwescheu AS, Enyong PA, Hoerauf A. Update on the distribution of Mansonella perstans in the southern part of Cameroon: influence of ecological factors and mass drug administration with ivermectin. Parasit Vectors. 2016 May 31;9(1):311. doi: 10.1186/s13071-016-1595-1[↩][↩]
- Bregani ER, Rovellini A, Mbaïdoum N, Magnini MG. Comparison of different anthelminthic drug regimens against Mansonella perstans filariasis. Trans R Soc Trop Med Hyg. 2006 May;100(5):458-63. doi: 10.1016/j.trstmh.2005.07.009[↩][↩][↩][↩]
- Ferreira MU, Crainey JL, Gobbi FG. The search for better treatment strategies for mansonellosis: an expert perspective. Expert Opin Pharmacother. 2023 Sep-Dec;24(15):1685-1692. doi: 10.1080/14656566.2023.2240235[↩][↩]
- Hübner MP, Martin C, Specht S, Koschel M, Dubben B, Frohberger SJ, Ehrens A, Fendler M, Struever D, Mitre E, Vallarino-Lhermitte N, Gokool S, Lustigman S, Schneider M, Townson S, Hoerauf A, Scandale I. Oxfendazole mediates macrofilaricidal efficacy against the filarial nematode Litomosoides sigmodontis in vivo and inhibits Onchocerca spec. motility in vitro. PLoS Negl Trop Dis. 2020 Jul 6;14(7):e0008427. doi: 10.1371/journal.pntd.0008427[↩][↩]
- Schiefer A, Hübner MP, Krome A, Lämmer C, Ehrens A, Aden T, Koschel M, Neufeld H, Chaverra-Muñoz L, Jansen R, Kehraus S, König GM, Pogorevc D, Müller R, Stadler M, Hüttel S, Hesterkamp T, Wagner K, Pfarr K, Hoerauf A. Corallopyronin A for short-course anti-wolbachial, macrofilaricidal treatment of filarial infections. PLoS Negl Trop Dis. 2020 Dec 7;14(12):e0008930. doi: 10.1371/journal.pntd.0008930[↩][↩]
- Krome AK, Becker T, Kehraus S, Schiefer A, Gütschow M, Chaverra-Muñoz L, Hüttel S, Jansen R, Stadler M, Ehrens A, Pogorevc D, Müller R, Hübner MP, Hesterkamp T, Pfarr K, Hoerauf A, Wagner KG, König GM. Corallopyronin A: antimicrobial discovery to preclinical development. Nat Prod Rep. 2022 Sep 21;39(9):1705-1720. doi: 10.1039/d2np00012a[↩][↩]
- Risch F, Kazakov A, Specht S, Pfarr K, Fischer PU, Hoerauf A, Hübner MP. The long and winding road towards new treatments against lymphatic filariasis and onchocerciasis. Trends Parasitol. 2024 Sep;40(9):829-845. doi: 10.1016/j.pt.2024.07.005[↩][↩]
- Hegde S, Marriott AE, Pionnier N, Steven A, Bulman C, Gunderson E, Vogel I, Koschel M, Ehrens A, Lustigman S, Voronin D, Tricoche N, Hoerauf A, Hübner MP, Sakanari J, Aljayyoussi G, Gusovsky F, Dagley J, Hong DW, O’Neill P, Ward SA, Taylor MJ, Turner JD. Combinations of the azaquinazoline anti-Wolbachia agent, AWZ1066S, with benzimidazole anthelmintics synergise to mediate sub-seven-day sterilising and curative efficacies in experimental models of filariasis. Front Microbiol. 2024 Feb 1;15:1346068. doi: 10.3389/fmicb.2024.1346068[↩][↩]
- Wanji S., Tayong D.B., Ebai R., Opoku V., Kien C.A., Ndongmo W.P.C., Njouendou A.J., Ghani R.N., Ritter M., Debrah Y.A., et al. Update on the biology and ecology of Culicoides species in the South-West region of Cameroon with implications on the transmission of Mansonella perstans. Parasit. Vectors. 2019;12:166. doi: 10.1186/s13071-019-3432-9[↩]
- Lane RP, Crosskey RW. Medical Insects and Arachnids. Heidelberg: Springer Netherlands; 1993.[↩]
- Crosskey RW. The Natural History of Blackflies. Chichester, UK: Wiley; 1990.[↩]
- Pichon G. Crypto-periodicity in Mansonella ozzardi. Trans R Soc Trop Med Hyg. 1983;77(3):331–333. doi: 10.1016/0035-9203(83)90155-4[↩]
- Sharp NA. Filaria perstans: its development in Culicoides austeni. Trans R Soc Trop Med Hyg. 1928;21:371–396.[↩]
- Simonsen PE, Fischer PU, Hoerauf A, Weil GJ. The filariasis. In: Farrar J, Hotez PJ, Junghanss T, Kang G, Lalloo D, White NJ, editors. Manson’s Tropical Diseases. 23rd ed. London: Saunders; 2014. pp. 737–765.[↩][↩][↩][↩][↩][↩][↩][↩]
- da Silva LB, Crainey JL, da Silva TR, et al. Molecular verification of New World Mansonella perstans Parasitemias. Emerg Infect Dis. 2017;23(3):545–547. doi: 10.3201/eid2303.161159[↩][↩]
- Crainey JL, da Silva TR, Luz SL. Historic accounts of Mansonella parasitaemias in the South Pacific and their relevance to lymphatic filariasis elimination efforts today. Asian Pac J Trop Med. 2016;9(3):205–210. doi: 10.1016/j.apjtm.2016.01.040[↩][↩]
- Muller R. Worms and Human Disease. 2nd ed. Wallingford, UK: CABI International; 2002.[↩][↩][↩]
- Fischer P, Bamuhiiga J, Büttner DW. Occurrence and diagnosis of Mansonella streptocerca in Uganda. Acta Trop. 1997;63(1):43–55. doi: 10.1016/s0001-706x(96)00607-9[↩][↩][↩][↩][↩][↩]
- Duke BO. The uptake of the microfilariae of Acanthocheilonema streptocerca by Culicoides grahamii, and their subsequent development. Ann Trop Med Parasitol. 1954;48(4):416–420. doi: 10.1080/00034983.1954.11685642[↩]
- Duke BO. The intake of the microfilariae of Acanthocheilonema streptocerca by Culicoides milnei, with some observations on the potentialities of the fly as a vector. Ann Trop Med Parasitol. 1958;52(2):123–128. doi: 10.1080/00034983.1958.11685852[↩]
- Bamuhiiga JT. Mansonella streptocerca: another filarial worm in the skin in Western Uganda. Community Eye Health. 1998;11(26):28. https://pmc.ncbi.nlm.nih.gov/articles/PMC1706049[↩]
- Simonsen PE, Fischer PU, Hoerauf A, Weil GJ. The filariasis. In: Farrar J, Hotez PJ, Junghanss T, Kang G, Lalloo D, White NJ, editors. Manson’s Tropical Diseases. 23rd ed. London: Saunders; 2014. pp. 737–765. [↩]
- Shelley AJ, Coscaron S. Simuliid blackflies (Diptera: Simuliidae) and ceratopogonid midges (Diptera: Ceratopogonidae) as vectors of Mansonella ozzardi (Nematoda: Onchocercidae) in northern Argentina. Mem Inst Oswaldo Cruz. 2001;96(4):451–458. doi: 10.1590/s0074-02762001000400003[↩]
- Lowrie RC, Jr, Raccurt CP, Eberhard ML, Katz SP. Assessment of Leptoconops bequaerti as a potential vector of Mansonella ozzardi in Haiti. Am J Trop Med Hyg. 1983;32(5):1013–1015. doi: 10.4269/ajtmh.1983.32.1013[↩]
- Basano SA, Camargo JS, Vera LJ, et al. Investigation of the occurrence of Mansonella ozzardi in the state of Rondonia, western Amazonia, Brazil. Rev Soc Bras Med Trop. 2011;44(5):600–603. doi: 10.1590/s0037-86822011005000055[↩]
- Bain O, Mutafchiev Y, Junker K, et al. Review of the genus Mansonella Faust, 1929 sensu lato (Nematoda: Onchocercidae), with descriptions of a new subgenus and a new subspecies. Zootaxa. 2015;3918(2):151–193. doi: 10.11646/zootaxa.3918.2.1[↩][↩]
- Orihel TC, Lowrie RC, Jr, Eberhard ML, et al. Susceptibility of laboratory primates to infection with Mansonella ozzardi from man. Am J Trop Med Hyg. 1981;30(4):790–794. doi: 10.4269/ajtmh.1981.30.790[↩][↩]
- Marcos LA, Arrospide N, Recuenco S, Cabezas C, Weil GJ, Fischer PU. Genetic characterization of atypical Mansonella (Mansonella) ozzardi microfilariae in human blood samples from northeastern Peru. Am J Trop Med Hyg. 2012;87(3):491–494. doi: 10.4269/ajtmh.2012.11-0379[↩]
- Lefoulon E, Bain O, Bourret J, et al. Shaking the tree: multi-locus sequence typing usurps current onchocercid (filarial nematode) phylogeny. PLoS Negl Trop Dis. 2015;9(11):e0004233. doi: 10.1371/journal.pntd.0004233[↩]
- Holmes GK, Gelfand M, Boyt W, Mackenzie P. A study to investigate the pathogenicity of a parasite resembling Acanthocheilonema perstans. Trans R Soc Trop Med Hyg. 1969;63(4):479-84. doi: 10.1016/0035-9203(69)90035-2[↩]
- Low GC. FILARIA PERSTANS. Br Med J. 1903 Mar 28;1(2204):722-4. doi: 10.1136/bmj.1.2204.722[↩]
- Asio SM, Simonsen PE, Onapa AW. Analysis of the 24-h microfilarial periodicity of Mansonella perstans. Parasitol Res. 2009 Mar;104(4):945-8. doi: 10.1007/s00436-008-1312-x[↩]
- Mediannikov O, Ranque S. Mansonellosis, the most neglected human filariasis. New Microbes New Infect. 2018 Sep 1;26:S19-S22. doi: 10.1016/j.nmni.2018.08.016[↩]
- Eberhard ML, Orihel TC. The genus Mansonella (syn. Tetrapetalonema): a new classification. Ann Parasitol Hum Comp. 1984;59(5):483-96. doi: 10.1051/parasite/1984595483[↩]
- Nelson GS. Filarial infections as zoonoses. J Helminthol. 1965;39(2):229-50. doi: 10.1017/s0022149x00020630[↩]
- Dolo H, Coulibaly YI, Dembele B, et al. Filariasis attenuates anemia and proinflammatory responses associated with clinical malaria: a matched prospective study in children and young adults. PLoS Negl Trop Dis. 2012;6(11):e1890. doi: 10.1371/journal.pntd.0001890[↩][↩]
- Charles LJ. Malaria in the Leeward and Windward islands, British West Indies. Am J Trop Med Hyg. 1952;1(6):941–961. doi: 10.4269/ajtmh.1952.1.941[↩]
- Bouyou-Akotet MK, Boussougou MN, Ovono-Abessolo F, Owono-Medang M, Kombila M. Influence of Mansonella perstans microfilaraemia on total IgE levels in Gabonese patients co-infected with Loa loa. Acta Trop. 2014;131:11–15. doi: 10.1016/j.actatropica.2013.11.012[↩]
- Keiser PB, Coulibaly YI, Keita F, et al. Clinical characteristics of post-treatment reactions to ivermectin/albendazole for Wuchereria bancrofti in a region co-endemic for Mansonella perstans. Am J Trop Med Hyg. 2003;69(3):331–335[↩]
- Moya L, Herrador Z, Ta-Tang TH, et al. Evidence for suppression of onchocerciasis transmission in Bioko Island, Equatorial Guinea. PLoS Negl Trop Dis. 2016;10(7):e0004829. doi: 10.1371/journal.pntd.0004829[↩][↩]
- Phillips RO, Frimpong M, Sarfo FS, et al. Infection with Mansonella perstans nematodes in Buruli ulcer patients, Ghana. Emerg Infect Dis. 2014;20(6):1000–1003. doi: 10.3201/eid2006.131501[↩]
- Rubin RH, Austen KF, Goetzl EJ. Studies of immediate hypersensitivity in a patient with Acanthocheilonema perstans filarial infection. J Infect Dis. 1975;131(Suppl):S98–S103. doi: 10.1093/infdis/131.supplement.s98[↩][↩][↩]
- Asgeirsson H, Harling A, Botero-Kleiven S. Successful treatment of 2 imported cases of Mansonella perstans infection. PLoS Negl Trop Dis. 2017;11(5):e0005452. doi: 10.1371/journal.pntd.0005452[↩][↩][↩]
- Bregani ER. Filariasis due to blood transfusion. Blood Transfus. 2010;8(2):129. doi: 10.2450/2009.0140-09[↩][↩]
- Baird JK, Neafie RC, Connor DH. Nodules in the conjunctiva, bung-eye, and bulge-eye in Africa caused by Mansonella perstans. Am J Trop Med Hyg. 1988;38(3):553–557. doi: 10.4269/ajtmh.1988.38.553[↩][↩][↩][↩][↩]
- Orihel TC, Eberhard ML. Zoonotic filariasis. Clin Microbiol Rev. 1998;11(2):366–381. doi: 10.1128/cmr.11.2.366[↩][↩][↩][↩]
- Otranto D, Eberhard ML. Zoonotic helminths affecting the human eye. Parasit Vectors. 2011;4:41. doi: 10.1186/1756-3305-4-41[↩]
- Martins M, Pessoa FA, de Medeiros MB, de Andrade EV, Medeiros JF. Mansonella ozzardi in Amazonas, Brazil: prevalence and distribution in the municipality of Coari, in the middle Solimões River. Mem Inst Oswaldo Cruz. 2010;105(3):246–253. doi: 10.1590/s0074-02762010000300002[↩]
- Medeiros JF, Py-Daniel V, Barbosa UC, Ogawa GM. Current profile of Mansonella ozzardi (Nematoda: Onchocercidae) in communities along the Ituxi river, Lábrea municipality, Amazonas, Brazil. Mem Inst Oswaldo Cruz. 2008;103(4):409–411. doi: 10.1590/s0074-02762008000400017[↩]
- Vianna LM, Martins M, Cohen MJ, Cohen JM, Belfort R., Jr Mansonella ozzardi corneal lesions in the Amazon: a cross-sectional study. BMJ Open. 2012;2(6):e001266. doi: 10.1136/bmjopen-2012-001266[↩]
- Garrido C, Campos M. First report of presumed parasitic keratitis in Indians from the Brazilian Amazon. Cornea. 2000;19(6):817–819. doi: 10.1097/00003226-200011000-00011[↩]
- Medeiros JF, Py-Daniel V, Barbosa UC. Prevalence of Mansonella ozzardi among riverine communities in the municipality of Lábrea, state of Amazonas, Brazil. Rev Soc Bras Med Trop. 2011;44(2):186–190. doi: 10.1590/s0037-86822011005000006[↩]
- Weller PF, Simon HB, Parkhurst BH, Medrek TF. Tourism-Acquired Mansonella ozzardi Microfilaremia in a Regular Blood Donor. JAMA. 1978;240(9):858–859. doi:10.1001/jama.1978.03290090052018[↩]
- Brown, M. Manson’s tropical diseases. Lancet Infect. Dis. 2009; 9:407-408. https://www.thelancet.com/pdfs/journals/laninf/PIIS1473-3099(09)70170-7.pdf[↩]
- ADOLPH PE, KAGAN IG, McQUAY RM. Diagnosis and treatment of Acanthocheilonema perstans filariasis. Am J Trop Med Hyg. 1962 Jan;11:76-88. doi: 10.4269/ajtmh.1962.11.76[↩][↩]
- Olumide YM, Obembe BM. Dipetalonema perstans in a patient with chronic lymphoedema. Case report. East Afr Med J. 1983 Mar;60(3):186-9.[↩][↩]
- Lansoud-Soukate J, Dupont A, De Reggi ML, Roelants GE, Capron A. Hypogonadism and ecdysteroid production in Loa loa and Mansonella perstans filariasis. Acta Trop. 1989 Jul;46(4):249-56. doi: 10.1016/0001-706x(89)90025-9[↩]
- Baird JK, Neafie RC, Connor DH. Nodules in the conjunctiva, bung-eye, and bulge-eye in Africa caused by Mansonella perstans. Am J Trop Med Hyg. 1988 May;38(3):553-7. doi: 10.4269/ajtmh.1988.38.553[↩][↩]
- Wiseman RA. Acanthocheilonema perstans, a cause of significant eosinophilia in the tropics: comments on its pathogenicity. Trans R Soc Trop Med Hyg. 1967;61(5):667-73. doi: 10.1016/0035-9203(67)90131-9[↩]
- Mpairwe H, Muhangi L, Ndibazza J, Tumusiime J, Muwanga M, Rodrigues LC, Elliott AM. Skin prick test reactivity to common allergens among women in Entebbe, Uganda. Trans R Soc Trop Med Hyg. 2008 Apr;102(4):367-73. doi: 10.1016/j.trstmh.2008.01.017[↩][↩]
- Almaviva M, Galli M, Rizzi M, Simonelli E, Negri C, Vigevani GM. Immune response in a symptomatic case of Tetrapetalonema perstans infection. Trans R Soc Trop Med Hyg. 1984;78(4):489-91. doi: 10.1016/0035-9203(84)90069-5[↩]
- Metenou S, Dembélé B, Konate S, Dolo H, Coulibaly SY, Coulibaly YI, Diallo AA, Soumaoro L, Coulibaly ME, Sanogo D, Doumbia SS, Wagner M, Traoré SF, Klion A, Mahanty S, Nutman TB. Patent filarial infection modulates malaria-specific type 1 cytokine responses in an IL-10-dependent manner in a filaria/malaria-coinfected population. J Immunol. 2009 Jul 15;183(2):916-24. doi: 10.4049/jimmunol.0900257[↩]
- Soboslay PT, Hamm DM, Pfäfflin F, Fendt J, Banla M, Schulz-Key H. Cytokine and chemokine responses in patients co-infected with Entamoeba histolytica/dispar, Necator americanus and Mansonella perstans and changes after anti-parasite treatment. Microbes Infect. 2006 Jan;8(1):238-47. doi: 10.1016/j.micinf.2005.06.019[↩]
- Semnani RT, Mahapatra L, Dembele B, Konate S, Metenou S, Dolo H, Coulibaly ME, Soumaoro L, Coulibaly SY, Sanogo D, Seriba Doumbia S, Diallo AA, Traoré SF, Klion A, Nutman TB, Mahanty S. Expanded numbers of circulating myeloid dendritic cells in patent human filarial infection reflect lower CCR1 expression. J Immunol. 2010 Nov 15;185(10):6364-72. doi: 10.4049/jimmunol.1001605[↩]
- Dolo H, Coulibaly YI, Dembele B, Konate S, Coulibaly SY, Doumbia SS, Diallo AA, Soumaoro L, Coulibaly ME, Diakite SA, Guindo A, Fay MP, Metenou S, Nutman TB, Klion AD. Filariasis attenuates anemia and proinflammatory responses associated with clinical malaria: a matched prospective study in children and young adults. PLoS Negl Trop Dis. 2012;6(11):e1890. doi: 10.1371/journal.pntd.0001890[↩]
- Lechner CJ, Komander K, Hegewald J, Huang X, Gantin RG, Soboslay PT, Agossou A, Banla M, Köhler C. Cytokine and chemokine responses to helminth and protozoan parasites and to fungus and mite allergens in neonates, children, adults, and the elderly. Immun Ageing. 2013 Jul 15;10(1):29. doi: 10.1186/1742-4933-10-29[↩]
- Ritter M, Ndongmo WPC, Njouendou AJ, Nghochuzie NN, Nchang LC, Tayong DB, Arndts K, Nausch N, Jacobsen M, Wanji S, Layland LE, Hoerauf A. Mansonella perstans microfilaremic individuals are characterized by enhanced type 2 helper T and regulatory T and B cell subsets and dampened systemic innate and adaptive immune responses. PLoS Negl Trop Dis. 2018 Jan 11;12(1):e0006184. doi: 10.1371/journal.pntd.0006184[↩][↩][↩]
- Stensgaard AS, Vounatsou P, Onapa AW, Utzinger J, Pedersen EM, Kristensen TK, Simonsen PE. Ecological Drivers of Mansonella perstans Infection in Uganda and Patterns of Co-endemicity with Lymphatic Filariasis and Malaria. PLoS Negl Trop Dis. 2016 Jan 21;10(1):e0004319. doi: 10.1371/journal.pntd.0004319[↩]
- Hillier SD, Booth M, Muhangi L, Nkurunziza P, Khihembo M, Kakande M, Sewankambo M, Kizindo R, Kizza M, Muwanga M, Elliott AM. Plasmodium falciparum and helminth coinfection in a semi urban population of pregnant women in Uganda. J Infect Dis. 2008 Sep 15;198(6):920-7. doi: 10.1086/591183[↩]
- Phillips RO, Frimpong M, Sarfo FS, Kretschmer B, Beissner M, Debrah A, Ampem-Amoako Y, Abass KM, Thompson W, Duah MS, Abotsi J, Adjei O, Fleischer B, Bretzel G, Wansbrough-Jones M, Jacobsen M. Infection with Mansonella perstans Nematodes in Buruli Ulcer Patients, Ghana. Emerg Infect Dis. 2014 Jun;20(6):1000-3. doi: 10.3201/eid2006.131501[↩]
- Elliott AM, Mawa PA, Webb EL, Nampijja M, Lyadda N, Bukusuba J, Kizza M, Namujju PB, Nabulime J, Ndibazza J, Muwanga M, Whitworth JA. Effects of maternal and infant co-infections, and of maternal immunisation, on the infant response to BCG and tetanus immunisation. Vaccine. 2010 Dec 16;29(2):247-55. doi: 10.1016/j.vaccine.2010.10.047[↩]
- Fernandes JS, Cardoso LS, Pitrez PM, Cruz ÁA. Helminths and Asthma: Risk and Protection. Immunol Allergy Clin North Am. 2019 Aug;39(3):417-427. doi: 10.1016/j.iac.2019.03.009[↩]
- Berbudi A, Ajendra J, Wardani AP, Hoerauf A, Hübner MP. Parasitic helminths and their beneficial impact on type 1 and type 2 diabetes. Diabetes Metab Res Rev. 2016 Mar;32(3):238-50. doi: 10.1002/dmrr.2673[↩]
- Lenz B, Andrew BA, Ritter M, Karunakaran I, Gandjui NVT, Nchang LC, Surendar J, Ebob AOB, Ehrens A, Klarmann-Schulz U, Ricchiuto A, Kuehlwein JM, Fombad FF, Ngwa AM, Katcho TD, Hoerauf A, Wanji S, Hübner MP. The design and development of a study protocol to investigate Onchocerca volvulus, Loa loa and Mansonella perstans-mediated modulation of the metabolic and immunological profile in lean and obese individuals in Cameroon. PLoS One. 2023 Jun 2;18(6):e0285689. doi: 10.1371/journal.pone.0285689[↩]
- Tang TH, López-Vélez R, Lanza M, Shelley AJ, Rubio JM, Luz SL. Nested PCR to detect and distinguish the sympatric filarial species Onchocerca volvulus, Mansonella ozzardi and Mansonella perstans in the Amazon Region. Mem Inst Oswaldo Cruz. 2010 Sep;105(6):823-8. doi: 10.1590/s0074-02762010000600016[↩]
- Nuchprayoon S, Junpee A, Poovorawan Y, Scott AL. Detection and differentiation of filarial parasites by universal primers and polymerase chain reaction-restriction fragment length polymorphism analysis. Am J Trop Med Hyg. 2005 Nov;73(5):895-900.[↩][↩]
- Pilotte N, Thomas T, Zulch MF, Sirois AR, Minetti C, Reimer LJ, Williams SA, Saunders LJ. Targeting a highly repetitive genomic sequence for sensitive and specific molecular detection of the filarial parasite Mansonella perstans from human blood and mosquitoes. PLoS Negl Trop Dis. 2022 Dec 29;16(12):e0010615. doi: 10.1371/journal.pntd.0010615. Erratum in: PLoS Negl Trop Dis. 2023 Jan 26;17(1):e0011096. doi: 10.1371/journal.pntd.0011096[↩]
- Crainey JL, Marín MA, Silva TRRD, de Medeiros JF, Pessoa FAC, Santos YV, Vicente ACP, Luz SLB. Mansonella ozzardi mitogenome and pseudogene characterisation provides new perspectives on filarial parasite systematics and CO-1 barcoding. Sci Rep. 2018 Apr 18;8(1):6158. doi: 10.1038/s41598-018-24382-3[↩]
- Chung M, Aluvathingal J, Bromley RE, Nadendla S, Fombad FF, Kien CA, Gandjui NVT, Njouendou AJ, Ritter M, Sadzewicz L, Tallon LJ, Wanji S, Hoerauf A, Pfarr K, Dunning Hotopp JC. Complete Mitochondrial Genome Sequence of Mansonella perstans. Microbiol Resour Announc. 2020 Jul 23;9(30):e00490-20. doi: 10.1128/MRA.00490-20[↩]
- Poole CB, Li Z, Alhassan A, Guelig D, Diesburg S, Tanner NA, Zhang Y, Evans TC Jr, LaBarre P, Wanji S, Burton RA, Carlow CK. Colorimetric tests for diagnosis of filarial infection and vector surveillance using non-instrumented nucleic acid loop-mediated isothermal amplification (NINA-LAMP). PLoS One. 2017 Feb 15;12(2):e0169011. doi: 10.1371/journal.pone.0169011[↩][↩][↩][↩]
- Amambo GN, Innocentia N, Abong RA, Fombad FF, Njouendou AJ, Nietcho F, Ekanya R, Kien CA, Ebai R, Lenz B, Ritter M, Esum ME, Deribe K, Cho JF, Beng AA, Enyong PI, Li Z, Hübner MP, Pfarr K, Hoerauf A, Carlow C, Wanji S. Application of loop mediated isothermal amplification (LAMP) assays for the detection of Onchocerca volvulus, Loa loa and Mansonella perstans in humans and vectors. Front Trop Dis. 2023 Jan 9;3:1016176. doi: 10.3389/fitd.2022.1016176[↩][↩]
- Rosenblatt JE. Laboratory diagnosis of infections due to blood and tissue parasites. Clin Infect Dis. 2009;49(7):1103–1108. doi: 10.1086/605574[↩]
- World Health Organization . Bench Aids for the Diagnosis of Filarial Infections. Geneva: WHO; 1997.[↩]
- Fischer P, Bamuhiiga J, Büttner DW. Treatment of human Mansonella streptocerca infection with ivermectin. Trop Med Int Health. 1997;2(2):191–199. doi: 10.1046/j.1365-3156.1997.d01-233.x[↩]
- Post RJ, Adams Z, Shelley AJ, Maia-Herzog M, Dias AP, Coscarón S. The morphological discrimination of microfilariae of Onchocerca volvulus from Mansonella ozzardi. Parasitology. 2003;127(Pt 1):21–27. doi: 10.1017/s003118200300324x[↩]
- Egwang TG, Nguiri C, Kombila M, Duong TH, Richard-Lenoble D. Elevated antifilarial IgG4 antibody levels in microfilaremic and microfilaridermic Gabonese adults and children. Am J Trop Med Hyg. 1993;49(1):135–142. doi: 10.4269/ajtmh.1993.49.135[↩]
- Alhassan A, Li Z, Poole CB, Carlow CK. Expanding the MDx toolbox for filarial diagnosis and surveillance. Trends Parasitol. 2015;31(8):391–400. doi: 10.1016/j.pt.2015.04.006[↩]
- Walther M, Muller R. Diagnosis of human filariases (except onchocerciasis) Adv Parasitol. 2003;53:149–193. doi: 10.1016/s0065-308x(03)53004-4[↩]
- Tang TH, Lopez-Velez R, Lanza M, Shelley AJ, Rubio JM, Luz SL. Nested PCR to detect and distinguish the sympatric filarial species Onchocerca volvulus, Mansonella ozzardi and Mansonella perstans in the Amazon region. Mem Inst Oswaldo Cruz. 2010;105(6):823–828. doi: 10.1590/s0074-02762010000600016[↩]
- Thiele EA, Cama VA, Lakwo T, et al. Detection of Onchocerca volvulus in skin snips by microscopy and real-time polymerase chain reaction: implications for monitoring and evaluation activities. Am J Trop Med Hyg. 2016;94(4):906–911. doi: 10.4269/ajtmh.15-0695[↩]
- Jimenez M, Gonzalez LM, Carranza C, et al. Detection and discrimination of Loa loa, Mansonella perstans and Wuchereria bancrofti by PCR-RFLP and nested-PCR of ribosomal DNA ITS1 region. Exp Parasitol. 2011;127(1):282–286. doi: 10.1016/j.exppara.2010.06.019[↩]
- Alhassan A, Osei-Atweneboana MY, Kyeremeh KF, et al. Comparison of a new visual isothermal nucleic acid amplification test with PCR and skin snip analysis for diagnosis of onchocerciasis in humans. Mol Biochem Parasitol. 2016;210(1–2):10–12. doi: 10.1016/j.molbiopara.2016.07.006[↩]
- Poole CB, Li Z, Alhassan A, et al. Colorimetric tests for diagnosis of filarial infection and vector surveillance using non-instrumented nucleic acid loop-mediated isothermal amplification (NINA-LAMP) PLoS One. 2017;12(2):e0169011. doi: 10.1371/journal.pone.0169011[↩]
- Ta-Tang TH, Luz SLB, Crainey JL, Rubio JM. An Overview of the Management of Mansonellosis. Res Rep Trop Med. 2021 May 24;12:93-105. doi: 10.2147/RRTM.S274684[↩]


{kind=link}