Contents
What is Mendelian inheritance
Mendelian inheritance also known as Mendelism refers to the inheritance of traits such as color and shape that are passed down from parents to their offsprings that is controlled by a single gene with two alleles (a variant form of a given gene), one of which may be completely dominant or recessive to the other (Figure 2) 1. Mendelian pattern of inheritance was first explained by the Austrian monk Gregor Mendel, who performed thousands of experiments with pea plants in the 19th century. Mendel developed three principles of inheritance that described the transmission of genetic traits, before anyone knew genes existed 2.
Mendel’s Laws of Heredity are usually stated as:
- The Law of Segregation: Each inherited trait is defined by a gene pair. Parental genes are randomly separated to the sex cells so that sex cells contain only one gene of the pair. Offspring therefore inherit one genetic allele from each parent when sex cells unite in fertilization.
- The Law of Independent Assortment: Genes for different traits are sorted separately from one another so that the inheritance of one trait is not dependent on the inheritance of another.
- The Law of Dominance: An organism with alternate forms of a gene will express the form that is dominant.
Humans have 23 pairs of chromosomes (Figure 1). Each chromosome has many genes. The human genome includes about 21,000 genes, spread out along 3 billion base pairs of DNA 3. This DNA is distributed among 23 chromosomes, of which you have two sets. You inherit one set from each parent. The first 22 pairs of chromosomes are called autosomes (any chromosome that is not a sex chromosome) 3. The last pair of chromosomes are called sex chromosomes (a pair of chromosomes that determines biological sex) and they are different between males and females 3. Most females have two X chromosomes (XX), and most males have an X and a Y (XY) chromosomes. Autosomes are all the chromosomes except the X or Y chromosome (sex chromosomes), and they do not differ between males and females, so autosomal traits are inherited in the same way, regardless of the sex of the parent or offspring. Traits controlled by genes on the sex chromosomes are called sex-linked traits. Because of the small size of the Y chromosome, most sex-linked traits are controlled by genes on the X chromosome. These traits are called X-linked traits. Single-gene X-linked traits have a different pattern of inheritance than single-gene autosomal traits, because males have just one X chromosome. In addition, males always inherit their X chromosome from their mother, and they pass on their X chromosome to all of their daughters, but none of their sons.
The dominant and recessive modes of inheritance of Mendelian traits depends on whether the traits are controlled by genes on autosomes, or by genes on sex chromosomes 4.
In 1800s, growers and breeders were puzzled that they could not fully explain the rules of inheritance. For generations, growers and breeders had produced detailed records of features such as the characteristics of cattle, the color of flowers, or the number of kernels in an ear of maize, but the inheritance process still seemed to be random. Sometimes, characteristic features were missing for several generations and then reappeared, and sometimes they disappeared completely. To get to the bottom of these issues, Mendel decided that it was necessary to first isolate clearly defined phenotypic traits. Mendel did not try to track all phenotypic traits at once but chose a few clearly defined characteristics such as plant height, flower color, or seed shape 5. He decided to use garden peas as his primary experimental model. Luckily, garden peas turned out to be a practical and suitable model, as garden peas took up little space, were cheap, and produced offspring quickly. Between 1854 and 1856, Mendel cultivated and tested thousands of pea plants because he assumed that the more independent measurements he made, the more likely it would be that he could rule out random phenomena. His exhaustive study included tests of 34 varieties of garden peas for trait consistency over several generations.
First, Mendel observed plant forms and their offspring for two years as they self-fertilized and ensured that their outward, measurable characteristics remained constant in each generation. During this time, Mendel observed seven different characteristics in the pea plants, and each of these characteristics had two forms. The characteristics included height (tall or short), pod shape (inflated or constricted), seed shape (smooth or winkled), pea color (green or yellow), and so on. In the years Mendel spent letting the plants self-fertilized, he verified the purity of his plants by confirming, for example, that tall plants had only tall children and grandchildren and so forth. Because the seven pea plant characteristics tracked by Mendel were consistent in generation after generation of self-fertilization, these parental lines of peas could be considered pure-breeders (in modern terminology, homozygous for the traits of interest). Mendel and his assistants eventually developed 22 varieties of pea plants with combinations of these consistent characteristics. Mendel also repeated his experiment to verify his results 6.
Based on his famous experiments of breeding different pea plants together and observing the traits present in their offsprings, Mendel proposed several laws of inheritance and coined key genetics terms like “dominant” and “recessive”. Mendel is famous for discovering the idea that hereditary elements are passed on in discrete units rather than “blended” together at each new generation. Today scientists call those discrete units “genes”. A gene is a segment of DNA instructions for building a particular RNA or protein molecule. Genes are very important, and yet they make up just 3% of your genome 3. Each gene is like an island separated by vast stretches of non-coding DNA—that is DNA that does not code for a product. Some of this DNA holds information about which cells should make proteins, when they should make it, and how much they should make. Other segments were left by viruses that infected our ancestors many generations ago 3. There is some DNA that scientists know is important—it is conserved between species—but scientists are not sure exactly why that is. Some DNA appears to have no function.
- A gene is a hereditary factor that determines (or influences) a particular trait. A gene is comprised of a specific DNA sequence and is located on a specific region of a specific chromosome. Because of its specific location, a gene can also be called a genetic locus.
- An allele is a particular variant of a gene, in the same way that chocolate and vanilla are particular variants of ice cream.
An organism’s genotype is the particular collection of alleles found in its DNA. An organism with two of the same alleles for a particular gene is homozygous at that locus; an organism with two different alleles for a particular gene is heterozygous at that locus. - A phenotype is its observable traits. An organism can have a heterozygous at a particular locus but have a phenotype that looks like only one of the two alleles. This is because some alleles mask the appearance of others in a dominant/recessive pattern.
- A dominant allele produces its phenotype whether the organism is homozygous or heterozygous at that locus. For example, in humans the allele for brown eyes is dominant to the allele for blue eyes, so a person who is heterozygous at the eye color locus will have brown eyes.
- A recessive allele produces its phenotype only when homozygous at the locus; its phenotype is masked if the locus is heterozygous. For example, a person must have two copies of the blue eye color allele to have blue eyes.
- Sometimes specific recessive alleles are associated with diseases. A person who is heterozygous for the gene will be phenotypically normal, but carry a copy of the recessive, disease-associated allele. This person is said to be a carrier and can pass on the disease allele to his or her offspring.
Before Mendel’s experiments, most people believed that traits in offspring resulted from a blending of the traits of each parent. However, when Mendel cross-pollinated one variety of purebred plant with another, these crosses would yield offspring that looked like either one of the parent plants, not a blend of the two. For example, when Mendel cross-fertilized plants with wrinkled seeds to those with smooth seeds, he did not get progeny with semi-wrinkly seeds. Instead, the offspring from this cross had only smooth seeds. For example, if a pea plant with the alleles T and t (T = tallness, t = shortness) is equal in height to a TT individual, the T allele (and the trait of tallness) is “dominant trait“. If the T t individual is shorter than the T T but still taller than the t t individual, T is partially or incompletely dominant—i.e., it has a greater influence than t but does not completely mask the presence of t, which is “recessive trait“. From this simple observation, Mendel proposed his first principle, the principle of uniformity; this principle states that all the offspring of a cross like this where the parents differ by only one trait will appear identical. Exceptions to the principle of uniformity include the phenomena of penetrance, expressivity, and sex-linkage, which were discovered after Mendel’s time.
Mendel’s data did not support the ideas about trait blending that were popular among the biologists of his time. As there were never any semi-wrinkled seeds or greenish-yellow seeds, for example, in the second generation (F2), Mendel concluded that blending should not be the expected outcome of parental trait combinations. Mendel instead hypothesized that each parent contributes some particulate matter to the offspring. He called this heritable substance “elementen”. In 1865, Mendel did not know about DNA or genes. For each of the traits he examined, Mendel focused on how the “elementen” that determined that trait was distributed among the offspring. Scientists now know that a single gene controls seed form, while another controls color, and so on, and that the “elementen” is actually the assembly of physical genes located on chromosomes. Multiple forms of those genes, known as alleles, represent the different traits. For example, one allele results in round seeds, and another allele specifies wrinkled seeds.
All of the concepts above are illustrated in the types of experiments that Mendel carried out with pea plants. Pea plants aren’t a particularly exciting organism to study, but they were very useful in figuring out basic patterns of inheritance. The reason pea plants were so useful is that they have a lot of traits that are caused by a single gene with a simple dominant or recessive inheritance pattern (this is actually pretty rare in general). A classic example is pea shape. Peas can be either round or wrinkly, but not anything in between. Whether peas are round or wrinkly is controlled by a single gene with two alleles, and the round allele is dominant to the wrinkly allele. The inheritance pattern if you cross homozygous round and homozygous wrinkly pea plants: in the first generation (F1) that all offspring produce round seeds, even though they both the round and wrinkly alleles (Figure 3). If the first generation (F1) self-fertilizes (pea plants – like most plants – produce both male and female gametes [male or female germ cell]), then you now see some offspring that produce rounds seeds and some that produce wrinkly seeds. The round:wrinkly seed producers exist in approximately a 3:1 ratio (Figure 3 and 4).
The inheritance patterns of single gene diseases are often referred to as Mendelian since Gregor Mendel first observed the different patterns of gene segregation for selected traits in garden peas and was able to determine probabilities of recurrence of a trait for subsequent generations. If a family is affected by a disease, an accurate family history will be important to establish a pattern of transmission. In addition, a family history can even help to exclude genetic diseases, particularly for common diseases where behavior and environment play strong roles.
Most genes have one or more versions due to mutations or polymorphisms referred to as alleles. Individuals may carry a ‘normal’ allele and/or a ‘disease’ or ‘rare’ allele depending on the impact of the mutation/polymorphism (e.g., disease or neutral) and the population frequency of the allele. Single-gene diseases are usually inherited in one of several patterns depending on the location of the gene and whether one or two normal copies of the gene are needed for the disease phenotype to manifest.
The expression of the mutated allele with respect to the normal allele can be characterized as dominant, co-dominant, or recessive. There are five basic modes of inheritance for single-gene diseases 7: autosomal dominant, autosomal recessive, X-linked dominant, X-linked recessive, and mitochondrial.
Figure 1. Human chromosomes
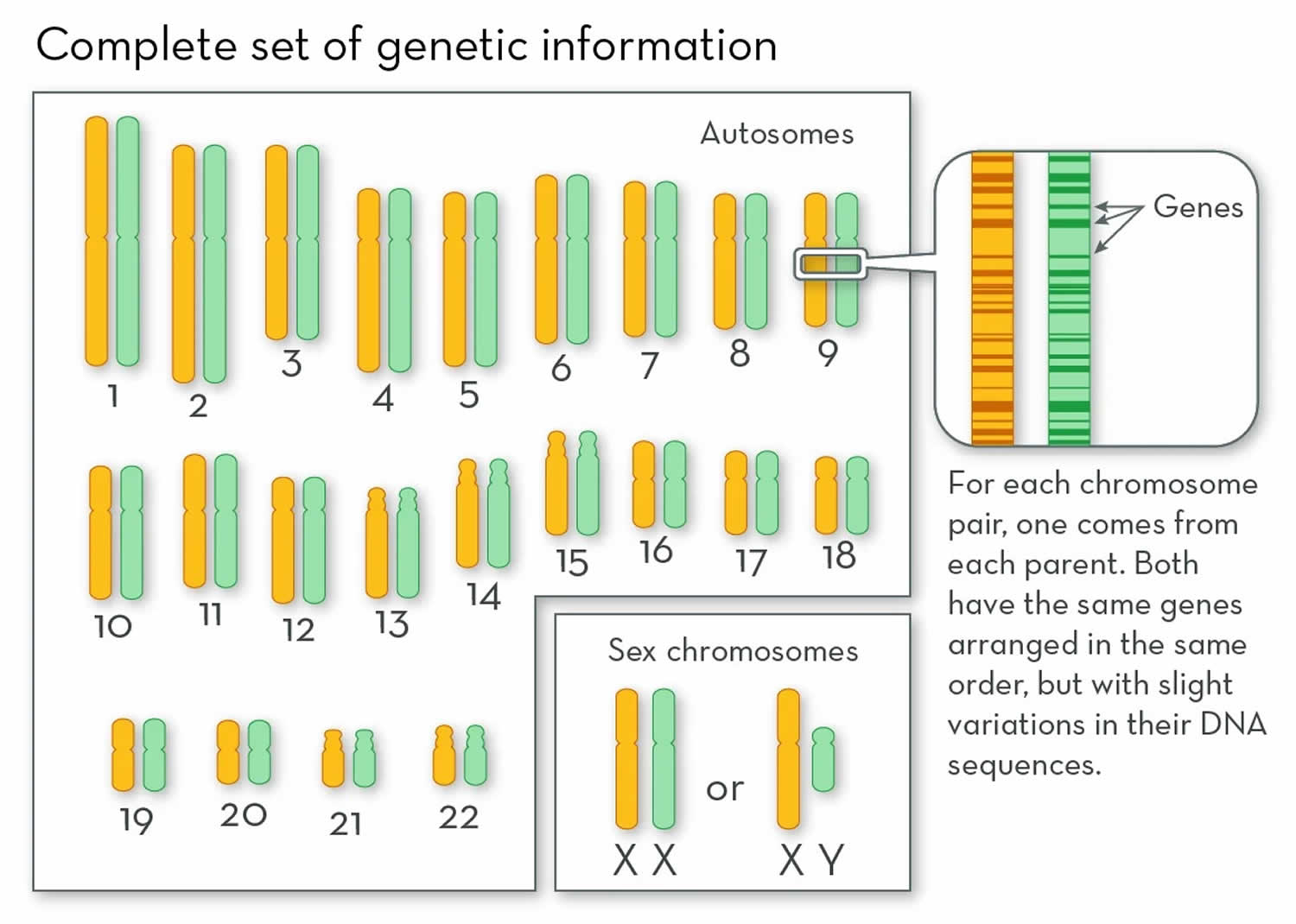
Footnote: Each chromosome includes a single, linear molecule of DNA with its own set of genes. Chromosomes are numbered according to their size, and genes are arranged along them in a specific order. So a gene near the tip of the short arm of chromosome 4 in one person will be found in the same location in everyone. This means that each of your genes can be mapped to a specific place on a specific chromosome.
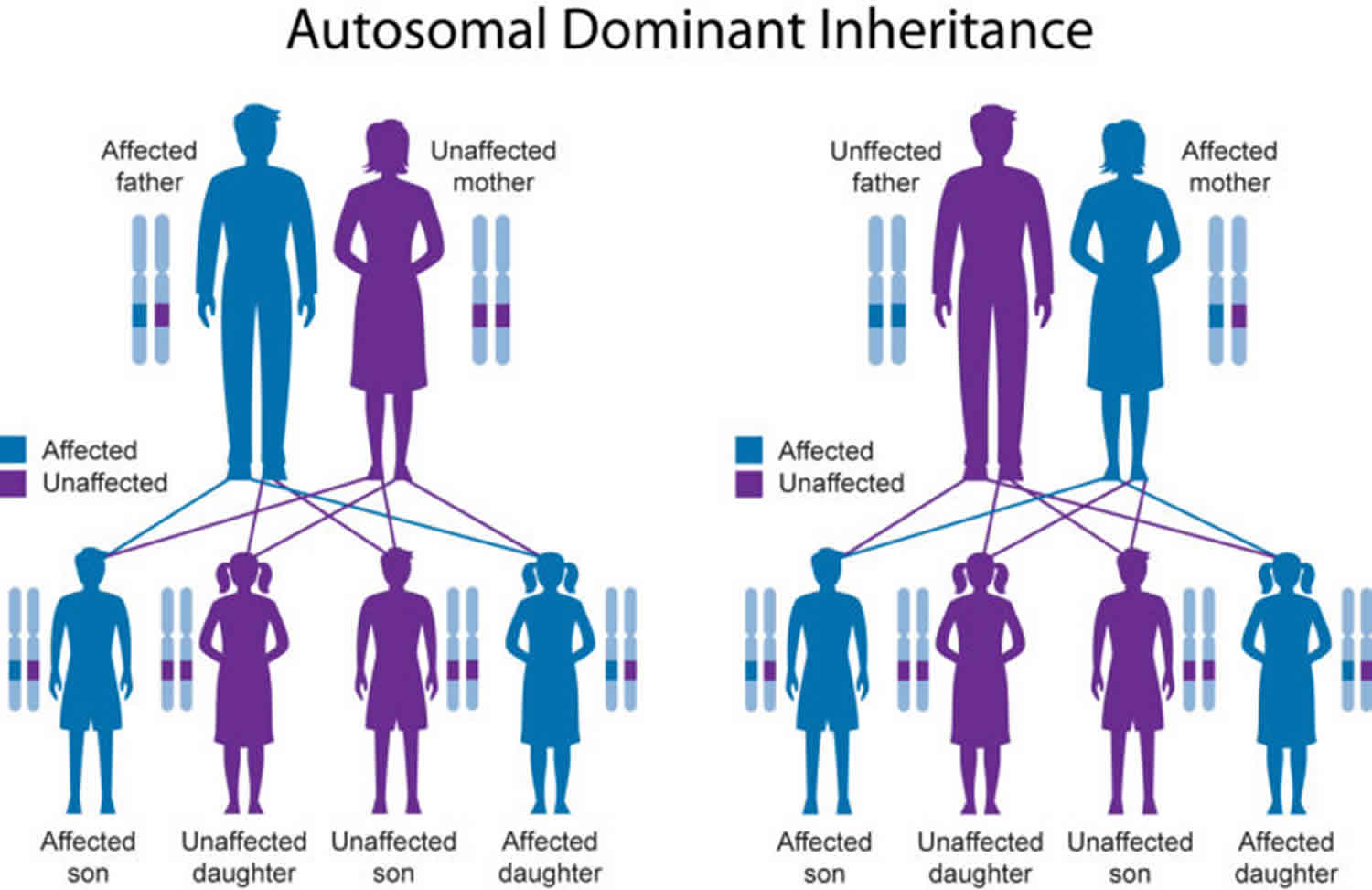
[Source 3 ]Figure 2. Mendelian patterns of inheritance (Pedigree chart)
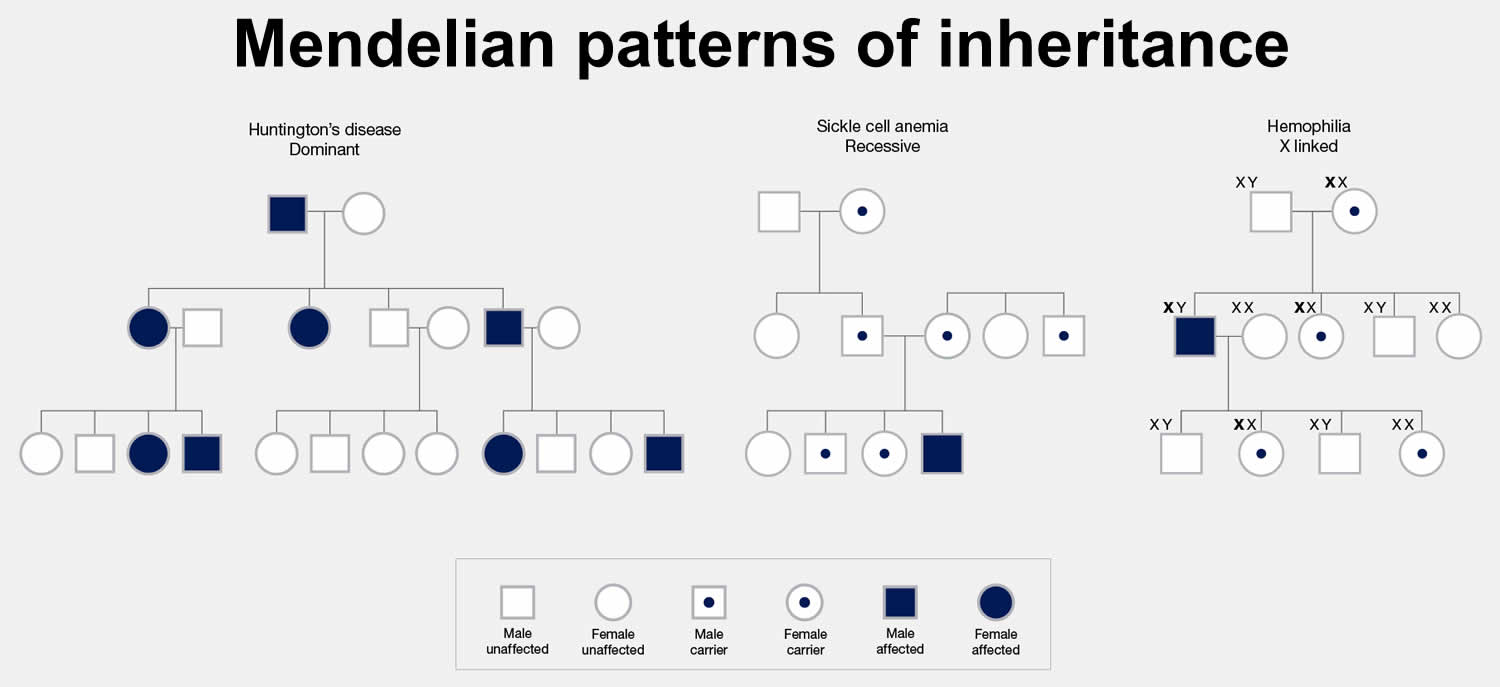
Footnote: The chart above is called a pedigree. A pedigree shows how a trait is passed from generation to generation within a family. A pedigree can show, for example, whether a Mendelian trait is an autosomal or X-linked trait. It can also be used to infer the genotype of different members of the family.
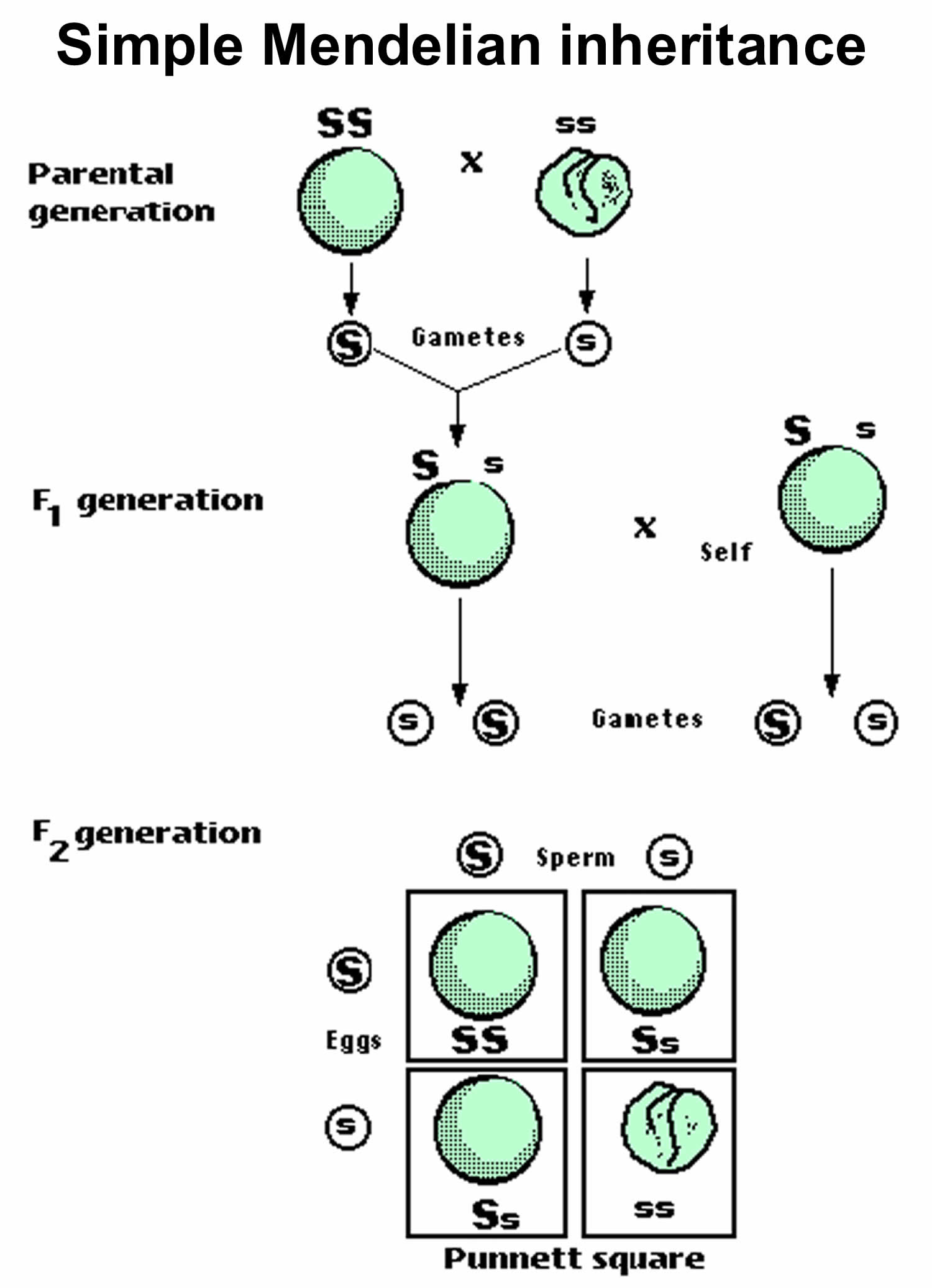
[Source 1 ]Figure 3. Monohybrid cross
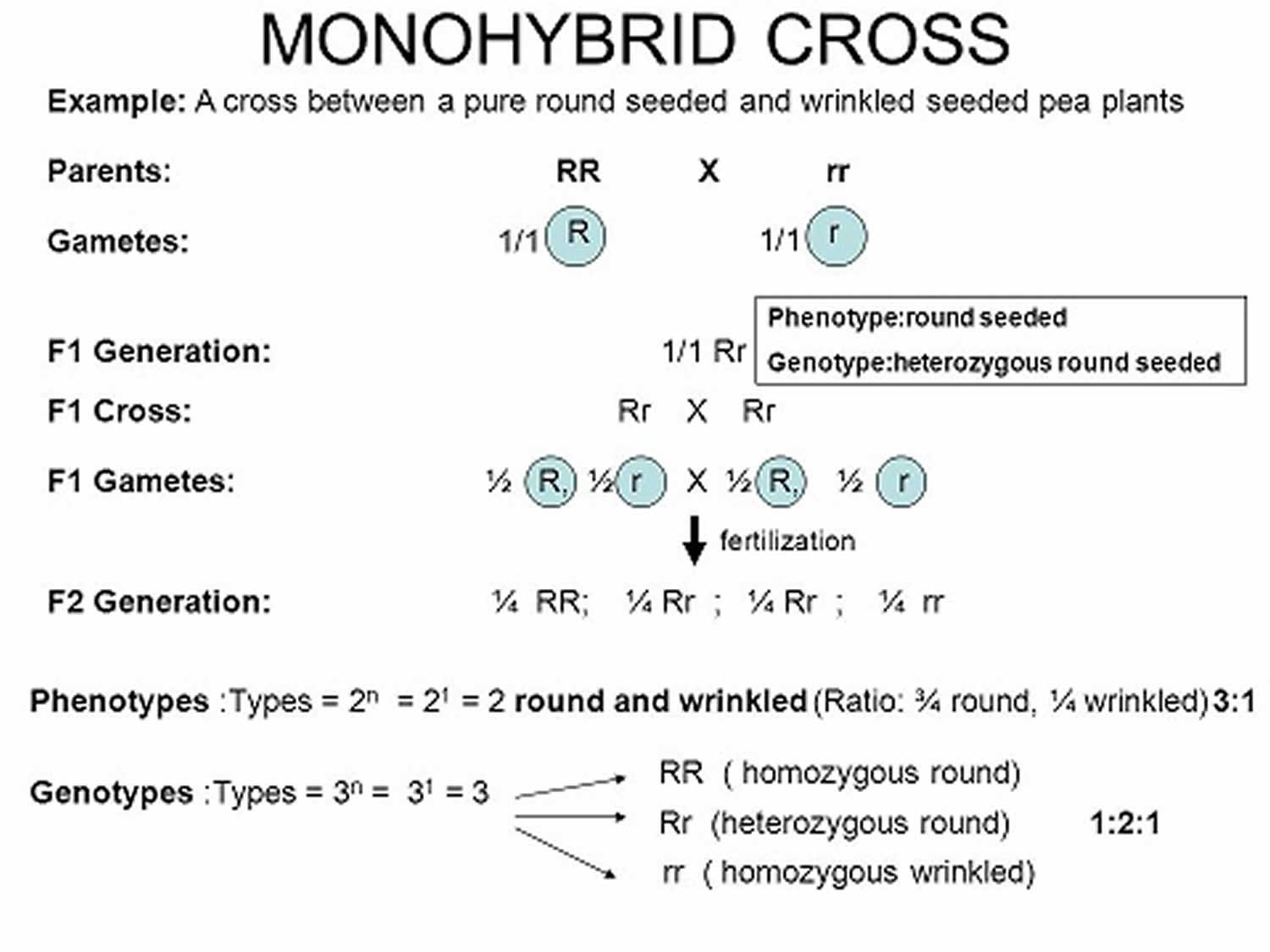
Footnotes: The genes for the traits of parents are pure round (RR) and pure wrinkled (rr). Round (R) which is a dominant trait and Wrinkled (r) which is a recessive trait. In first generation (F1), Mendel cross-pollinated a pure round pea (RR) and a pure wrinkled pea (rr). He called them parental generation (P). He collected the seeds produced by this cross and grew plants from them. All these plants were found to be round. The first generation (F1) plants were pollinated among themselves called selfing. The second generation (F2) plants were found to contain both round and wrinkled individual. They were in the ration of 3 round and 1 wrinkle.
Figure 4. Punnett Square
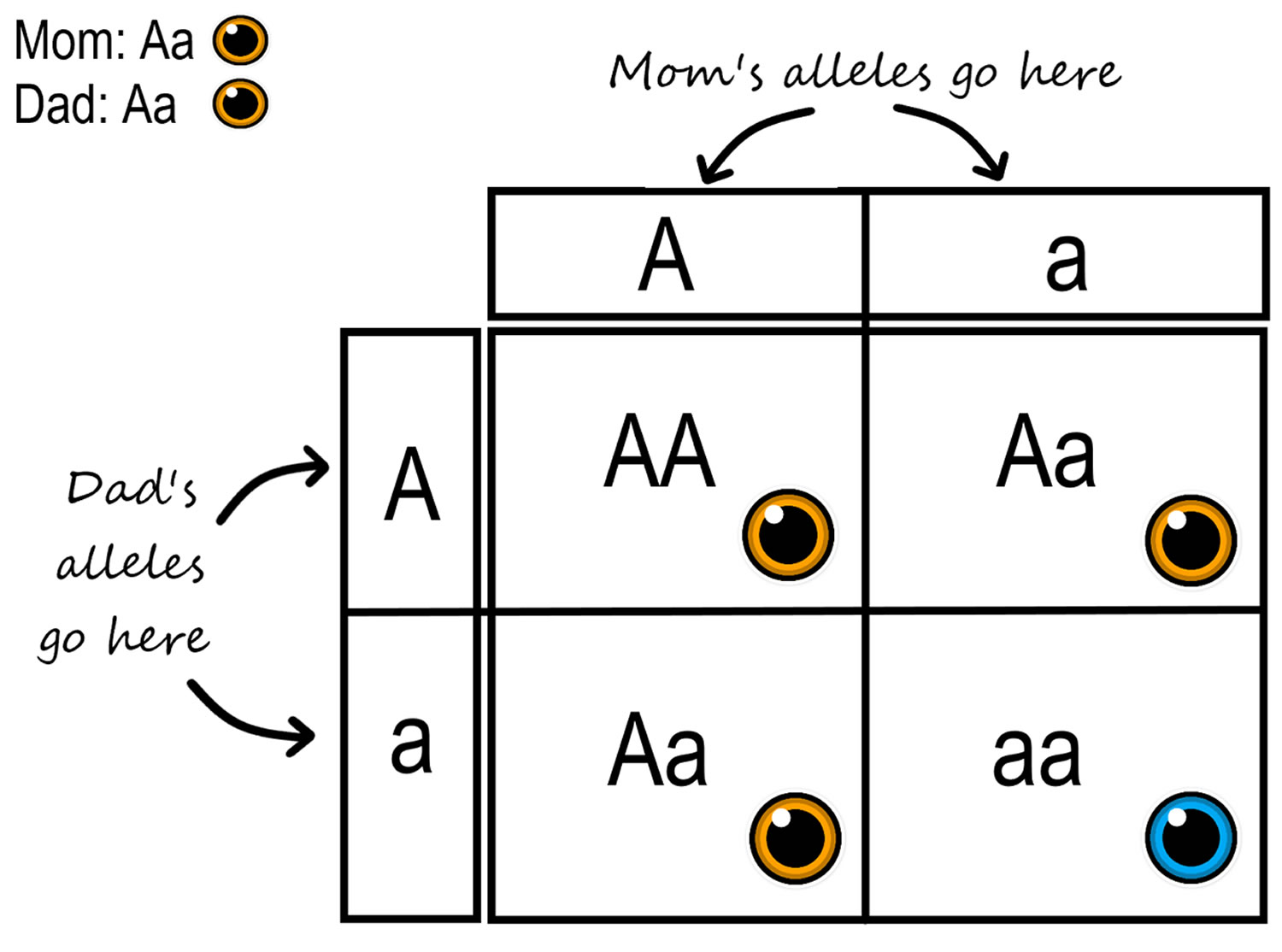
Footnotes: Punnett Square is a chart that allows you to easily determine the expected ratios of possible genotypes in the offspring of two parents. You can see a hypothetical example in figure above. In this case, the gene is autosomal, and both parents are heterozygotes (Aa) for the gene. Half of the gametes (male or female germ cell) produced by each parent will have the A allele, and half will have the a allele. That’s because the two alleles are on homologous chromosomes, which always separate and go to separate gametes during meiosis. The alleles in the gametes from each parent are written down the side and across the top of the Punnett square. Filling in the cells of the Punnett square gives the possible genotypes of their children. It also shows the most likely ratios of the genotypes, which in this case is 25 per cent AA, 50 per cent Aa, and 25 per cent aa.
[Source 4 ]Types of mendelian inheritance
There are five basic modes of inheritance for single-gene diseases 7: autosomal dominant, autosomal recessive, X-linked dominant, X-linked recessive, and mitochondrial.
Genetic heterogeneity is a common phenomenon with both single-gene diseases and complex multi-factorial diseases 7. It should not be surprising that multiple affected family members may experience different levels of disease severity and outcomes. This effect may be due to other genes influencing the disease phenotype or different mutations in the same gene resulting in similar, but not identical phenotypes.
Autosomal Dominant
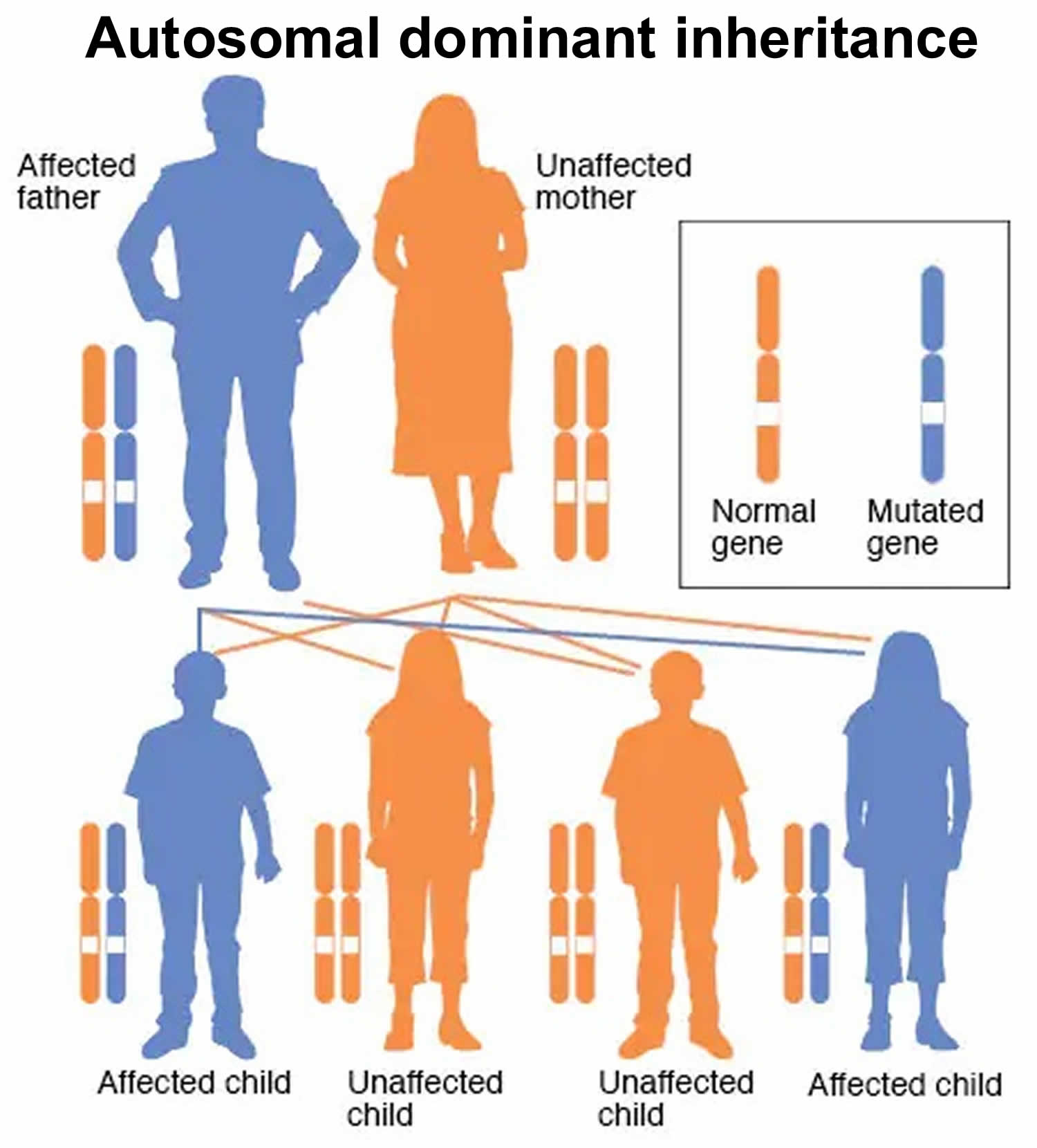
Autosomal dominant means one copy of the abnormal (mutated) dominant gene from only one parent or in each cell is sufficient to cause the disorder or disease. “Autosomal” means that the gene in question is located on one of the chromosome that is not a sex chromosome, or autosomes. “Dominant” means that a single copy of the mutated gene (from one parent) is enough to cause the disorder. In some cases, an affected person inherits the autosomal dominant condition from an affected parent. In others, the autosomal dominant condition may result from a new mutation in the gene and occur in people with no history of the disorder in their family. This is called a de novo mutation.
A child of a person affected by an autosomal dominant condition has a 50% chance of being affected by that condition via inheritance of a dominant allele.
- Each affected person has an affected parent
- Occurs in every generation
- When one parent has the abnormal gene, they will pass on either their normal gene or their abnormal gene to their child. Each of their children therefore has a 50% (1 in 2) chance of inheriting the changed gene and being affected by the condition.
- There is also a 50% (1 in 2) chance that a child will inherit the normal copy of the gene. If this happens the child will not be affected by the disorder and cannot pass it on to any of his or her children.
- A person only needs to inherit one copy of the abnormal gene in order to be affected by the condition (50% chance). These outcomes occur randomly. They remain the same in every pregnancy and are the same for boys and girls.
Autosomal Dominant disease examples:
- Huntington’s disease,
- Marfan syndrome
- Neurofibromatosis,
- Achondroplasia,
- Familial hypercholesterolemia
Figure 5. Autosomal dominant inheritance pattern
Autosomal Recessive
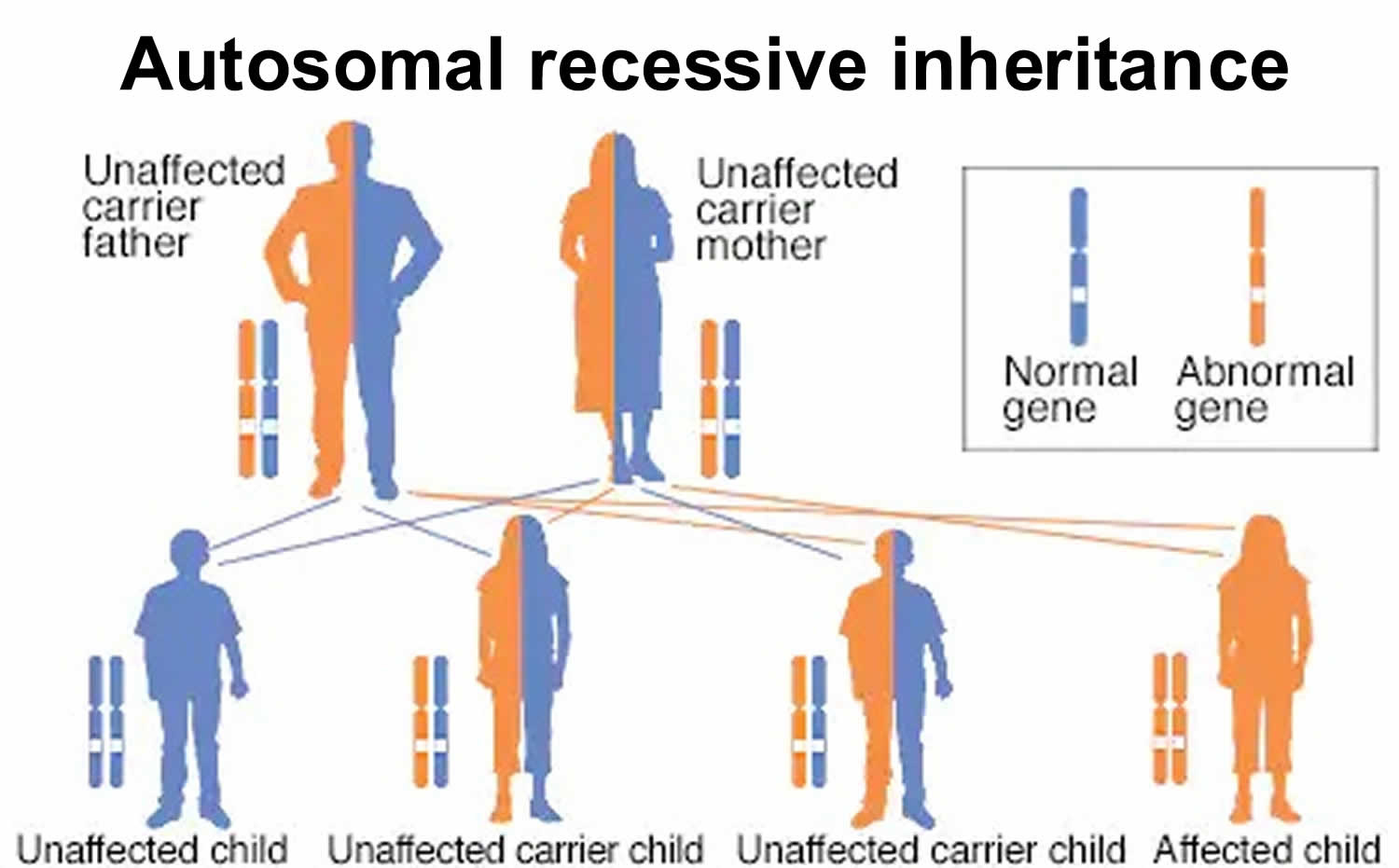
In an autosomal recessive disorder, the abnormal (mutated) recessive gene is located on one of the non-sex chromosomes (autosomes). To inherit an autosomal recessive disorder — such as cystic fibrosis, sickle cell anemia or phenylketonuria (PKU) — both parents must be carriers of the abnormal recessive gene. The affected child must inherits two copies of the abnormal gene — one from each parent.
A carrier is an unaffected individual who has one copy of the abnormal gene and the other copy is normal. With each pregnancy, two carriers have a 25% chance of having an unaffected child with two normal copies of a gene, a 50% chance of having an unaffected child who also is a carrier and a 25% chance of having an affected child with two abnormal copies of the gene.
- Both parents of an affected person are carriers
- Not typically seen in every generation
Autosomal Recessive disease examples:
- Tay-sachs disease,
- Sickle cell anemia,
- Cystic fibrosis,
- Phenylketonuria (PKU)
Figure 6. Autosomal recessive inheritance pattern
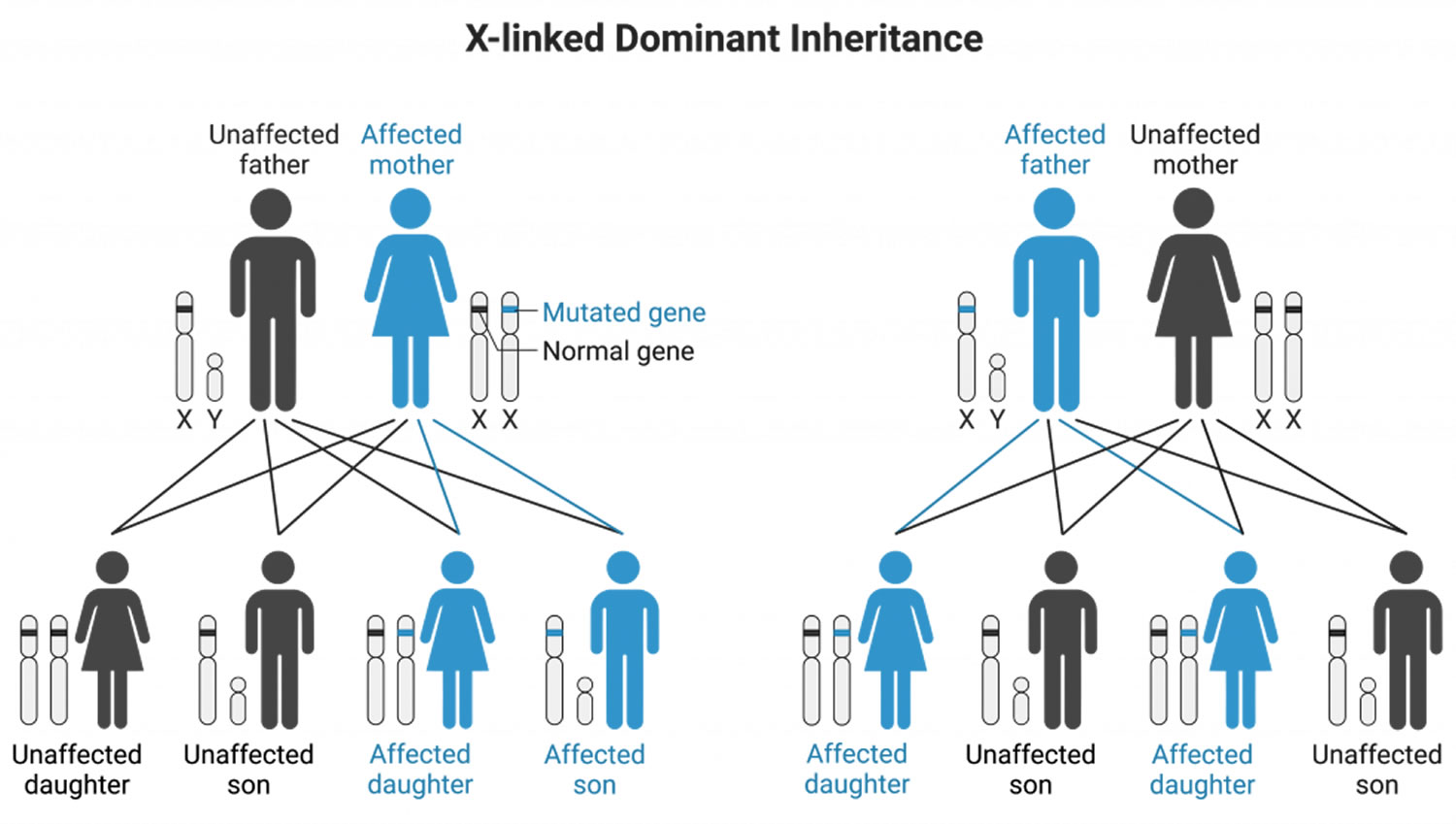
X-linked Dominant
X-linked Dominant Inheritance, the abnormal (mutated) gene is on the X chromosome. Affected father with X-linked Dominant disorder will pass on his X chromosome (with the abnormal gene) and condition to all of his daughters. A man with an X-linked Dominant disorder will pass his unaffected Y chromosome to his sons, and none will be affected. Sons of an affected, heterozygous mother (carrier) have a 50% chance of being affected.
- Females more frequently affected
- Can have affected males and females in same generation
X-linked Dominant disease examples:
- Hypophatemic rickets (vitamin D-resistant rickets),
- Ornithine transcarbamylase deficiency
Figure 7. X-linked Dominant inheritance pattern
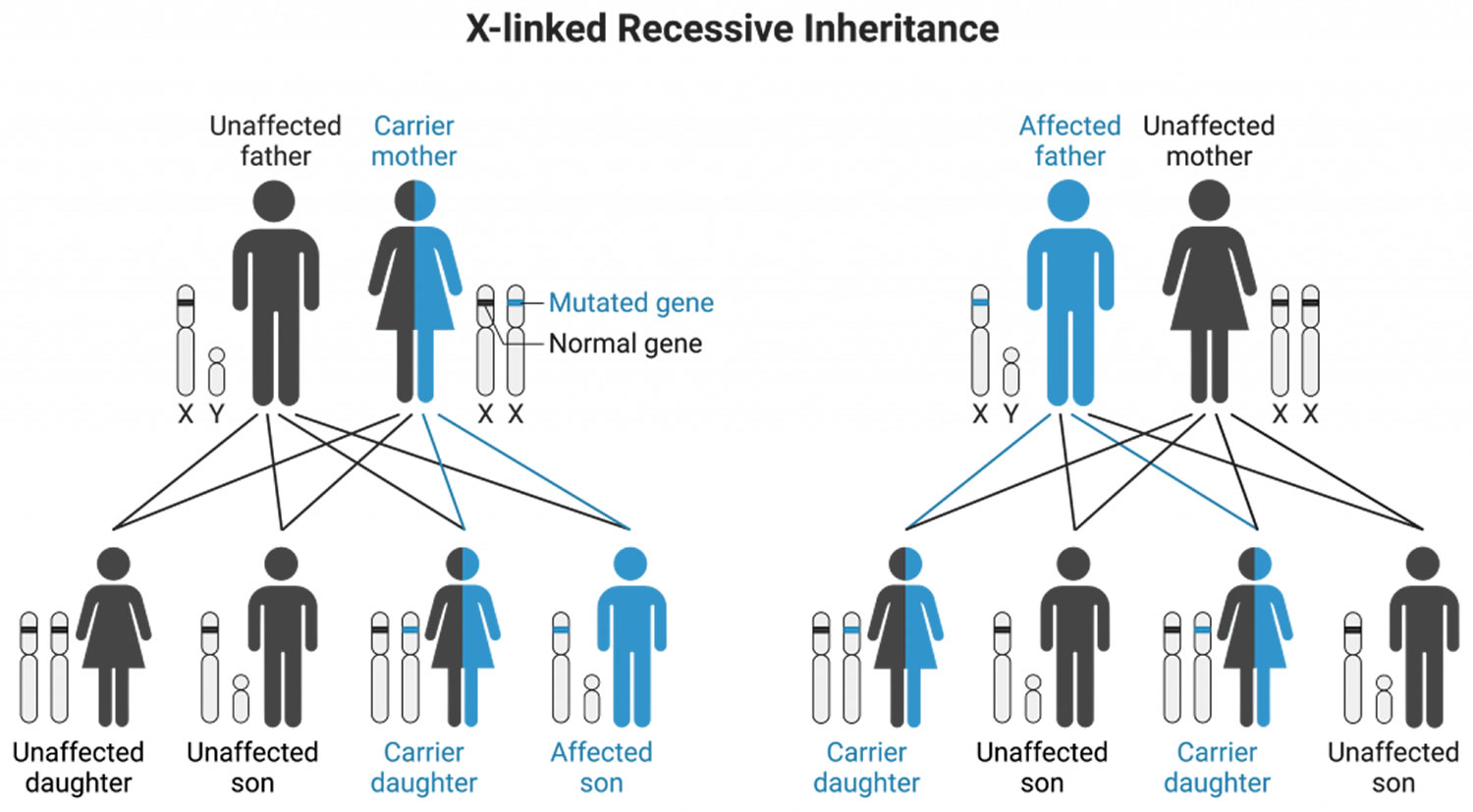
X-linked Recessive
X-linked recessive inheritance pattern, the abnormal (mutated) gene is on the X chromosome. Duchenne muscular dystrophy, some types of colorblindness and hemophilia A are examples of X-linked recessive disorders. A man with an X-linked recessive disorder will pass his unaffected Y chromosome to his sons, and none will be affected. He will pass his X chromosome (with the abnormal gene) to his daughters, and all will be carriers of the disease. His daughters (carrier) may have no symptoms or only mild features of the disorder, and they may pass the abnormal gene to their children.
Female carriers of an X-linked recessive disorder often have no symptoms, though some have mild symptoms of the disorder. This is because female carriers have one normal copy of the gene and one abnormal (mutated) copy. The normal copy typically can compensate for the abnormal copy.
Males have only one X chromosome and will usually be affected if there is an abnormal copy of a gene on the X-chromosome.
A woman who is a carrier of an X-linked recessive disorder has a 25% chance of each of the following results with each pregnancy:
- Having an unaffected son
- Having an unaffected daughter
- Having an unaffected daughter who also is a carrier
- Having an affected son
X-linked Recessive disease examples:
- Hemophilia A,
- Duchenne muscular dystrophy
Figure 8. X-linked Recessive inheritance pattern
Mitochondrial inheritance
Mitochondria are located in a cell’s cytoplasm and contain circular DNA, called mitochondrial DNA (mtDNA) (Figure 9). Mitochondrial DNA (mtDNA) exists separately from a cell’s nuclear
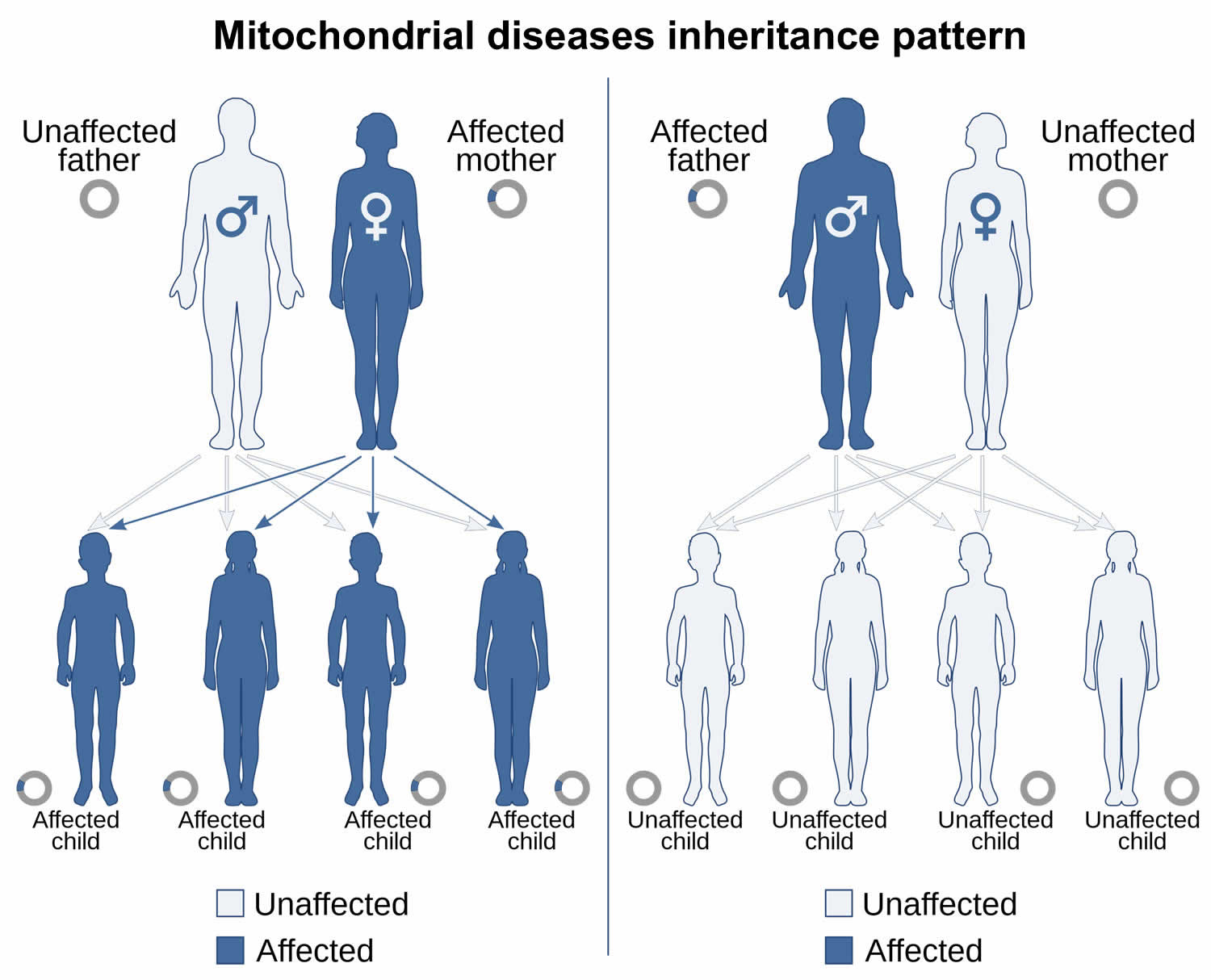
genome and is inherited solely through the maternal lineage non-Mendelian inheritance. Mitochondrial DNA (mtDNA) in humans is always inherited from a person’s mother by a non-Mendelian pattern. Mitochondria can only be passed from one generation to the next only through egg cells (not through sperm cells), only females pass mitochondrial DNA (mtDNA)-associated condition to their children. A family history in which males and females are affected, affected females transmit the disease to all their children, and affected males do not transmit the disease to their children suggests mitochondrial inheritance. Mitochondrial diseases (diseases due to malfunction of mitochondria) can be inherited both maternally and through chromosomal inheritance, the way in which they are passed on from generation to generation can vary greatly depending on the disease. Mitochondrial genetic mutations that occur in the nuclear DNA can occur in any of the chromosomes (depending on the species). Mutations inherited through the chromosomes can be autosomal dominant or recessive and can also be sex-linked dominant or recessive.
- Can affect both males and females, but only passed on by females
- Can appear in every generation
Mitochondrial disease examples:
- Leber’s hereditary optic neuropathy,
- Kearns-Sayre syndrome,
- Chronic progressive external ophthalmoplegia,
- Pearson syndrome
- Leigh syndrome
- Mitochondrial encephalopathy, lactic acidosis, stroke-like episodes (MELAS),
- Myoclonus, epilepsy, and ragged-red fibers (MERRF),
- Neurogenic weakness with ataxia and retinitis pigmentosa (NARP)
Figure 9. Mitochondrial DNA
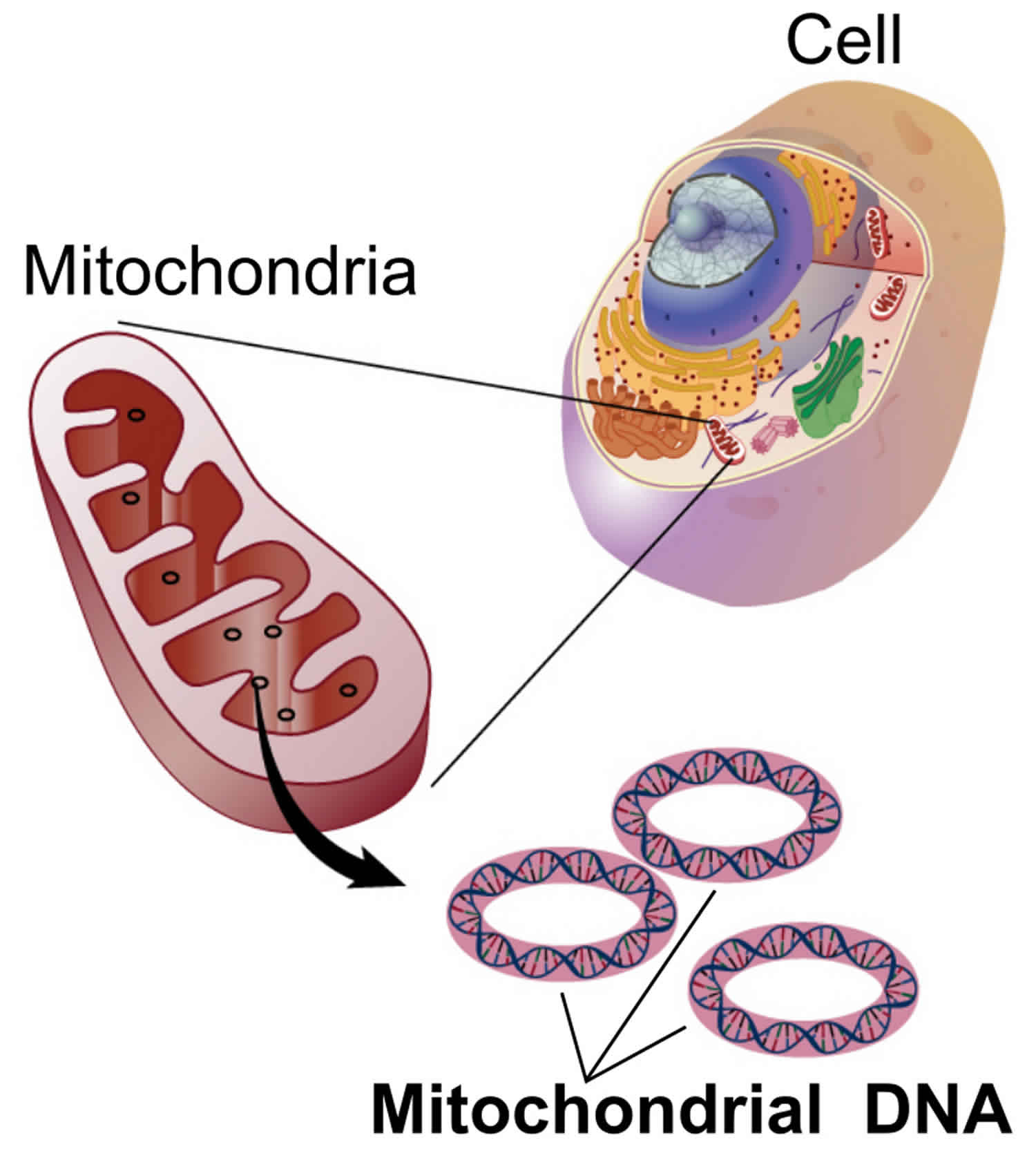
Figure 10. Mitochondrial inheritance
What is Non-Mendelian inheritance?
Non-Mendelian inheritance is any pattern of inheritance that have more complex genetic basis in which traits do not segregate in accordance with Mendel’s laws. Non-Mendelian inheritance includes inheritance of multiple allele traits, codominance, incomplete dominance and polygenic traits.
Multiple allele traits
The majority of human genes are thought to have more than two normal versions also known as alleles. Traits controlled by a single gene with more than two alleles are called multiple allele traits. An example is ABO blood type. Your blood type refers to which of certain proteins called antigens are found on your red blood cells. There are three common alleles for this trait, which are represented by the letters A, B, and O. there are six possible ABO genotypes, because the three alleles, taken two at a time, result in six possible combinations. The A and B alleles are dominant to the O allele. As a result, both AA and AO genotypes have the same phenotype, with the A antigen in their blood (type A blood). Similarly, both BB and BO genotypes have the same phenotype, with the B antigen in their blood (type B blood). No antigen is associated with the O allele, so people with the OO genotype have no antigens for ABO blood type in their blood (type O blood).
As shown in Table 1 below there are six possible ABO genotypes, because the three alleles (A, B, and O), taken two at a time, result in six possible combinations. The A and B alleles are dominant to the O allele. As a result, both AA and AO genotypes have the same phenotype, with the A antigen in their blood (type A blood). Similarly, both BB and BO genotypes have the same phenotype, with the B antigen in their blood (type B blood). No antigen is associated with the O allele, so people with the OO genotype have no antigens for ABO blood type in their blood (type O blood).
Figure 11. Multiple allele traits
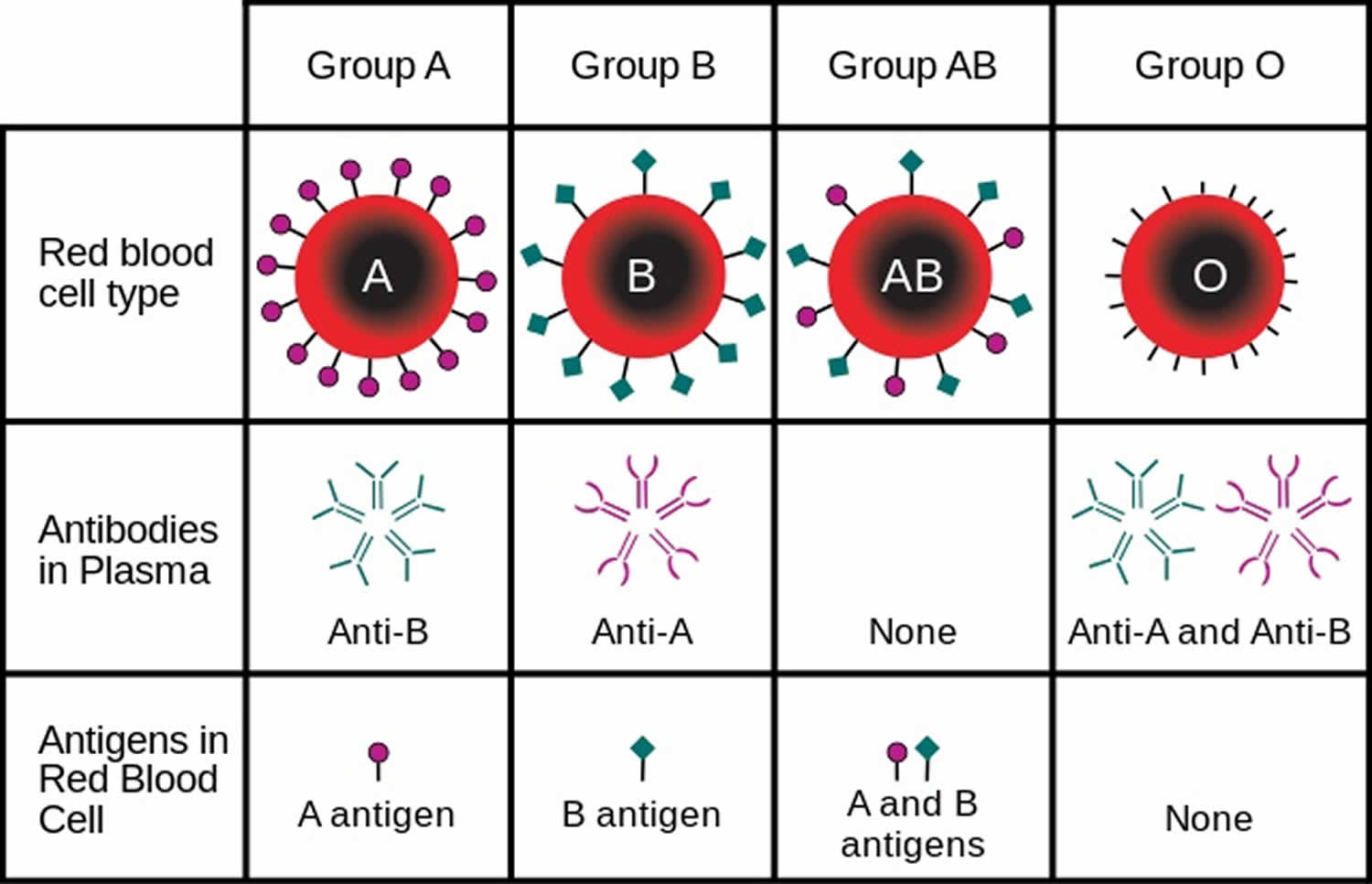
Table 1. ABO Blood Group System
| Genotype | Blood Type (Phenotype) |
|---|---|
| AA | A |
| AO | A |
| BB | B |
| BO | B |
| OO | O |
| AB | AB |
Codominance
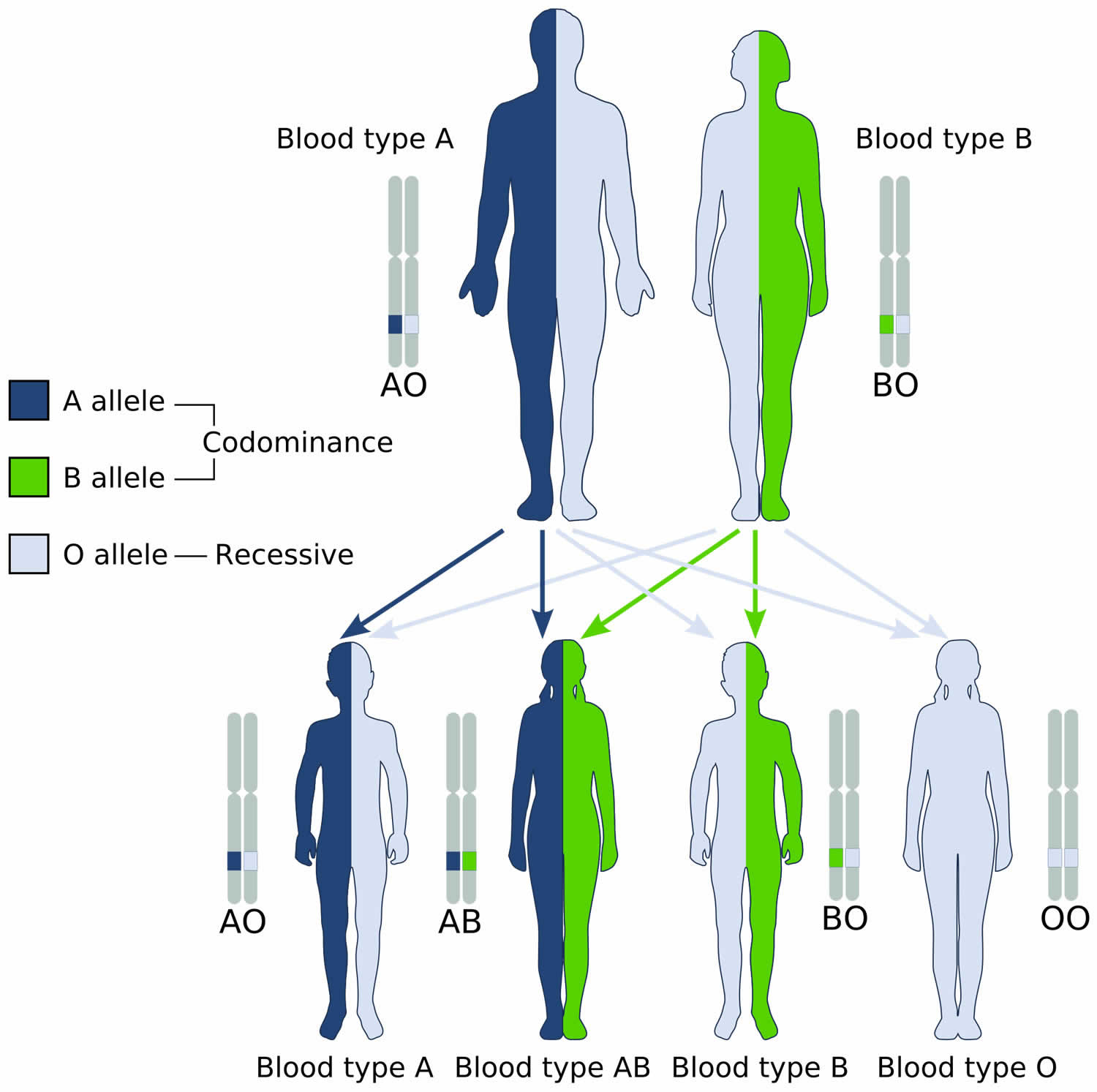
Codominance occurs when the contributions of both alleles for a gene are expressed equally in the phenotype of heterozygotes and neither allele masks another. For example, the genotype AB in the ABO blood group system, alleles A and B for ABO blood type are neither dominant nor recessive to one another. Instead, they are codominant. In the case of ABO blood type, AB heterozygotes have a unique phenotype, with both A and B antigens in their blood (type AB blood).
Figure 12. Codominance
Incomplete Dominance
Another relationship that may occur between alleles for the same gene is incomplete dominance. This occurs when the dominant allele is not completely dominant. In this case, an intermediate phenotype results in heterozygotes who inherit both alleles. Generally, this happens when the two alleles for a given gene both produce proteins, but one protein is not functional. As a result, the heterozygote individual produces only half the amount of normal protein as is produced by an individual who is homozygous for the normal allele.
An example of incomplete dominance in humans is Tay Sachs disease. The normal allele for the gene in this case produces an enzyme that is responsible for breaking down lipids. A defective allele for the gene results in the production of a nonfunctional enzyme. Heterozygotes who have one normal and one defective allele produce half as much functional enzyme as the normal homozygote, and this is enough for normal development. Homozygotes who have only defective allele, however, produce only nonfunctional enzyme. This leads to the accumulation of lipids in the brain starting in utero, which causes significant brain damage. Most individuals with Tay Sachs disease die at a young age, typically by the age of five years.
Another good example of incomplete dominance in humans is hair type. There are genes for straight and curly hair, and if an individual is heterozygous, they will typically have the phenotype of wavy hair.
Figure 13. Incomplete dominance hair type

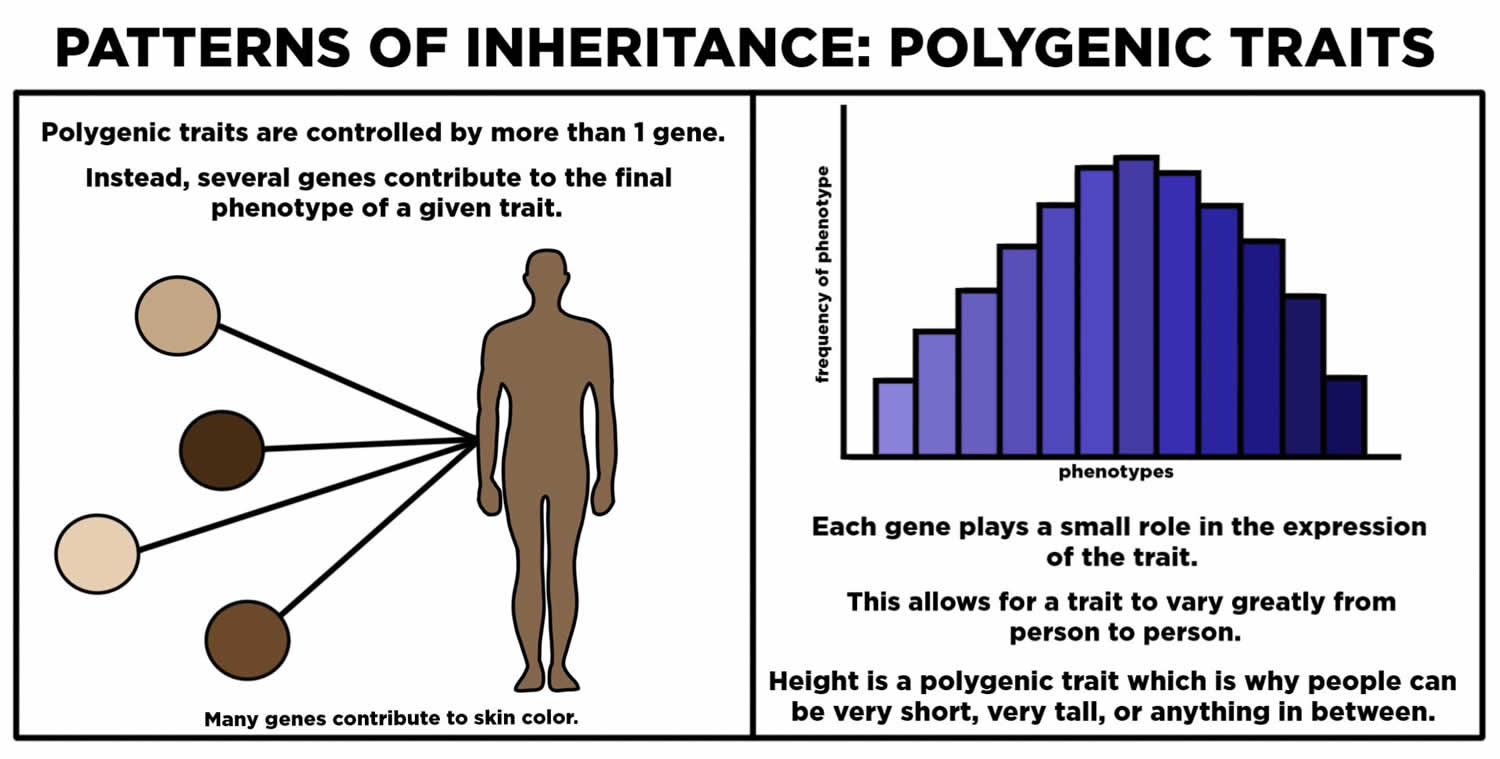
Polygenic Traits
Many human traits are controlled by more than one gene. These traits are called polygenic traits. The alleles of each gene have a minor additive effect on the phenotype. There are many possible combinations of alleles, especially if each gene has multiple alleles. Therefore, a whole continuum of phenotypes is possible.
An example of a human polygenic trait is adult height. Several genes, each with more than one allele, contribute to this trait, so there are many possible adult heights. One adult’s height might be 1.655 m (5.430 feet), and another adult’s height might be 1.656 m (5.433 feet). Adult height ranges from less than 5 feet to more than 6 feet, with males, on average, being somewhat taller than females. The majority of people fall near the middle of the range of heights for their sex.
Figure 14. Polygenic traits
Environmental Effects on Phenotype
Many traits are affected by the environment, as well as by genes. This may be especially true for polygenic traits. Adult height, for example, might be negatively impacted by poor diet or childhood illness. Skin color is another polygenic trait. There is a wide range of skin colors in people worldwide. In addition to differences in genes, differences in exposure to ultraviolet (UV) light cause some variation.
Pleiotropy
Some genes affect more than one phenotypic trait. This is called pleiotropy. There are numerous examples of pleiotropy in humans. They generally involve important proteins that are needed for the normal development or functioning of more than one organ system. An example of pleiotropy in humans occurs with the gene that codes for the main protein in collagen, a substance that helps form bones. This protein is also important in the ears and eyes. Mutations in the gene result in problems not only in bones, but also in these sensory organs, which is how the gene’s pleiotropic effects were discovered.
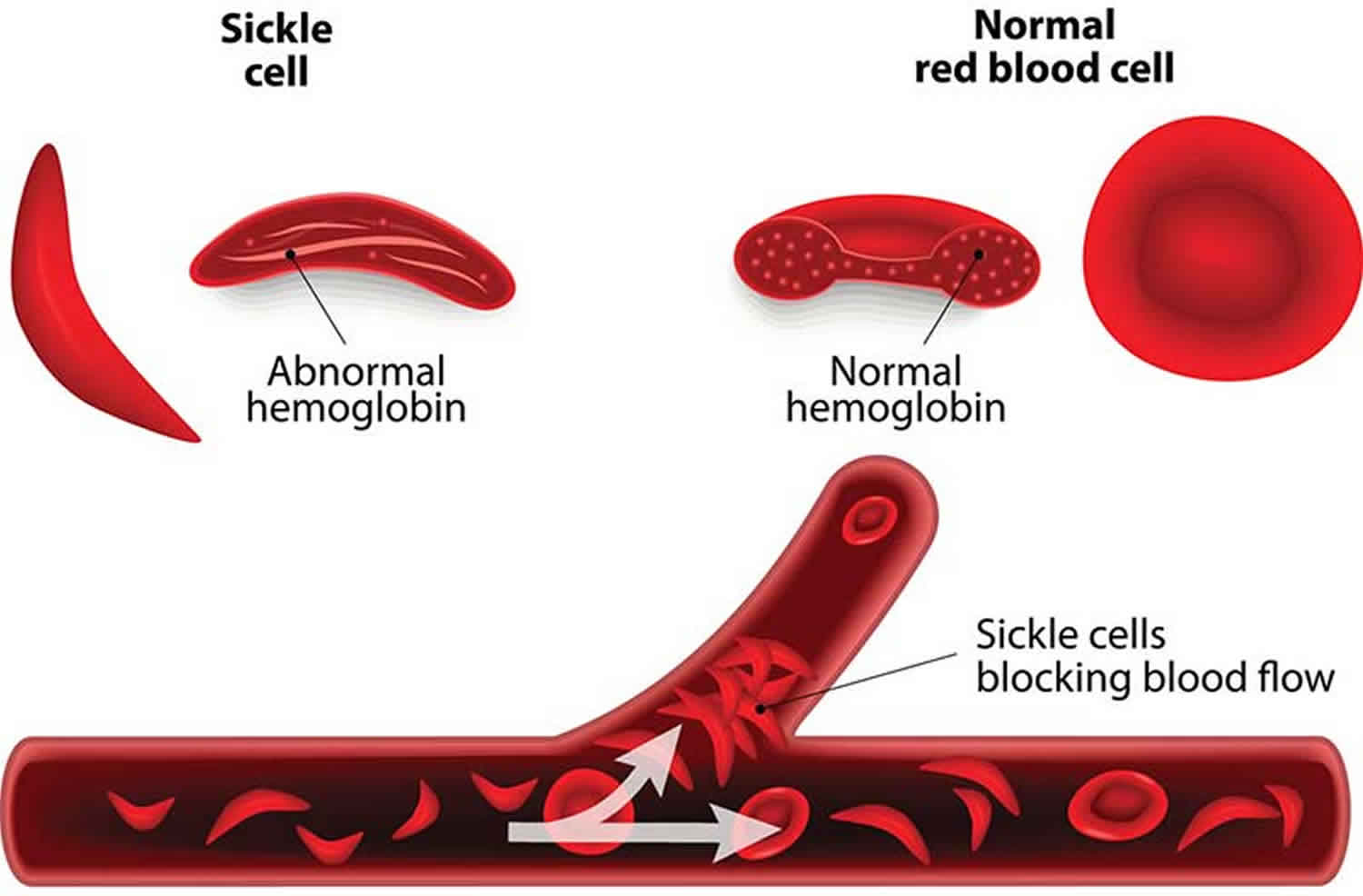
Another example of pleiotropy occurs with sickle cell anemia. This recessive genetic disorder occurs when there is a mutation in the gene that normally encodes the red blood cell protein called hemoglobin. People with sickle cell anemia have two alleles for sickle cell hemoglobin, so named for the sickle shape that their red blood cells take on under certain conditions (like physical exertion). The sickle-shaped red blood cells clog small blood vessels, causing multiple phenotypic effects, including stunting of physical growth, certain bone deformities, kidney failure, and strokes.
Figure 15. Pleiotropy
Epistasis
Epistasis is the case in which a gene affects the expression of other genes. For example, a mutation in one gene may not allow other genes to be expressed in the phenotype. Epistasis is similar to dominance, except that it occurs between different genes, rather than between different alleles for the same gene. Albinism is an example of epistasis. A person with albinism has virtually no pigment in the skin. The condition occurs due to an entirely different gene than the genes that encode skin color. Albinism occurs because a protein called tyrosinase, which is needed for the production of normal skin pigment, is not produced, due to a gene mutation. If an individual has the albinism mutation, he or she will not have any skin pigment, regardless of the skin color genes that were inherited.
Figure 16. Epistasis

What is the difference between mendelian and non mendelian inheritance?
Mendelian inheritance also known as Mendelism refers to the inheritance of traits linked to single genes on chromosomes in the nucleus. In Mendelian inheritance, each parent contributes one of two possible alleles for a trait. If the genotypes of both parents in a genetic cross are known, Mendel’s laws can be used to determine the distribution of phenotypes expected for the population of offspring.
Non-Mendelian inheritance is any pattern of inheritance that have more complex genetic basis in which traits do not segregate in accordance with Mendel’s laws. Non-Mendelian inheritance includes inheritance of multiple allele traits, codominance, incomplete dominance and polygenic traits. For example, the skin color trait has a more complex genetic basis than just one gene with two alleles, which is the type of simple trait that Mendel studied in pea plants.
- Mendelian Inheritance. https://www.genome.gov/genetics-glossary/Mendelian-Inheritance[↩][↩]
- Miko, I. (2008) Gregor Mendel and the principles of inheritance. Nature Education 1(1):134[↩]
- What Are Chromosomes? https://learn.genetics.utah.edu/content/genetics/chromosomes[↩][↩][↩][↩][↩][↩]
- Mendelian Inheritance. https://humanbiology.pressbooks.tru.ca/chapter/5-12-mendelian-inheritance[↩][↩]
- Raudenska M, Vicar T, Gumulec J, Masarik M. Johann Gregor Mendel: the victory of statistics over human imagination. Eur J Hum Genet. 2023 Jul;31(7):744-748. doi: 10.1038/s41431-023-01303-1[↩]
- Abbott S, Fairbanks DJ. Experiments on Plant Hybrids by Gregor Mendel. Genetics. 2016;204:407–22. doi: 10.1534/genetics.116.195198[↩]
- Genetic Alliance; District of Columbia Department of Health. Understanding Genetics: A District of Columbia Guide for Patients and Health Professionals. Washington (DC): Genetic Alliance; 2010 Feb 17. Appendix B, Classic Mendelian Genetics (Patterns of Inheritance) Available from: https://www.ncbi.nlm.nih.gov/books/NBK132145[↩][↩][↩]

{kind=link}