
Contents
What is Gherlin Hormone ?
Ghrelin is 28-amino-acid peptide (a chain of natural biological amino acids) that was discovered from the rat and human stomach in 1999 1. Since the discovery of ghrelin, various functions of ghrelin, including growth hormone release, feeding behavior, glucose metabolism, memory, and also antidepressant effects, have been studied. Although there are several neuropeptides stimulating food intake, ghrelin is the only known appetite-stimulating hormone in humans. Its circulating levels increase during fasting and decrease following a meal 2. Ghrelin is considered to be a gut-brain peptide in that ghrelin is found in the stomach and hypothalamus but is abundantly produced from endocrine cells in the gastrointestinal mucosa (the stomach). In the gastrointestinal tract, ghrelin cells are most abundant in the stomach and are localized in gastric mucosal layers. Ghrelin cells are also widely distributed throughout the gastrointestinal tract. And gastric ghrelin is thought to play important multiple physiological roles in peripheral signaling. Many factors including nutrients as well as circulating and gastric hormones were found to be involved in the regulation of mRNA and release of ghrelin, indicating that ghrelin cells are under the control of complicated mechanisms, which are still obscure 1. In addition, characteristics of ghrelin cells in the stomach and other parts of the gastrointestinal tract are different; closed-type ghrelin cells are localized in the stomach and many opened-type ghrelin cells are localized throughout the small and large intestines. Since the length of the gastrointestinal tract is long, considerable amounts of plasma ghrelin can be assumed to be derived from ghrelin cells in small and large intestines. Further studies are needed to understand the physiological and pathological roles of ghrelin cells in both small and large intestines 1.
Ghrelin has at least two different forms: acylated (‘active’ or ‘octanoylated’) and non-acylated (‘des-n-octanoyl’ or ‘desacyl’) 3. Acylated ghrelin is thought to be more potent than non-acylated ghrelin in stimulating appetite. Whereas acylated ghrelin increases appetite and feeding, non-acylated ghrelin may actually decrease feeding behaviour 4. In fact, researchers have proposed inhibition of ghrelin-O-acyltransferace for reducing appetite by preventing acylation of ghrelin 5. Although acylated ghrelin makes up only 10–20% of total circulating ghrelin levels, much research has relied on measurements of total ghrelin; researchers have recently emphasized the need to assess both acylated and non-acylated forms. Ghrelin binds to the growth-hormone secretagogue receptors (GHSR) 6.
The growth hormone secretagogue receptor (GHSR), also known as the ghrelin receptor, is involved in mediating a wide variety of biological effects of ghrelin, including: stimulation of growth hormone release, increase of food intake and body weight, modulation of glucose and lipid metabolism, regulation of gastrointestinal motility and secretion, protection of neuronal and cardiovascular cells, and regulation of immune function 7. Dependent on the tissues and cells, activation of growth-hormone secretagogue receptors may trigger a diversity of signaling mechanisms and subsequent distinct physiological responses.
The ghrelin receptor, growth hormone secretagogue receptor (GHSR) is widely expressed at the mRNA level in various peripheral tissues (pituitary, stomach, intestine, pancreas, adipose tissue, adrenal gland, thyroid gland, myocardium, spleen, immune cells and on vagal afferents) and the brain 8, 9, 10, 11, 12, 13 indicative of broader functions besides the regulation of food intake. In the brain, growth hormone secretagogue receptor (GHSR) mRNA is localized in the arcuate nucleus, hypothalamic ventromedial nucleus, hippocampus (CA2 and CA3 areas and dentate gyrus), ventral tegmental area, substantia nigra as well as raphe magnus nucleus 14 and receptor expression has been confirmed at the protein level in the hypothalamus 15, 16.
To date, the physiological functions of growth hormone secretagogue receptor have been extended to include:
- The release of various hormones such as growth hormone, adrenocorticotropic hormone, cortisol, and prolactin 17;
- Modulation of food intake and energy metabolism 18;
- Influences on glucose and lipid metabolism 17;
- Regulation of gastrointestinal motility and secretion 19 and pancreatic function 20;
- Regulation of cell proliferation and survival 21, 22;
- Attenuation of proinflammatory cascades and regulation of immune function that play important roles in aging and gastrointestinal homeostasis 23; and
- Cell protection in the nervous and the cardiovascular systems 24, 25, 26. Such diversified functions of growth hormone secretagogue receptor suggest the complexity of GHSR-mediated intracellular signaling. Numerous intracellular signaling pathways have been proposed upon activation of growth hormone secretagogue receptor.
- The body’s response to hypoglycemia (low blood glucose) is to mount a stress response, including the release of corticotropin releasing hormone and growth hormone releasing hormone (GnRH) from the hypothalamus to stimulate corticotropin and growth hormone (GH) that act to raise blood glucose. This study 27 suggests that the ghrelin-producing cell senses glucose to regulate ghrelin release. Ghrelin is essential for maintaining the level of GH required in prevention of neuroglycopenia (shortage of glucose in the brain) and thus, preservation of life.
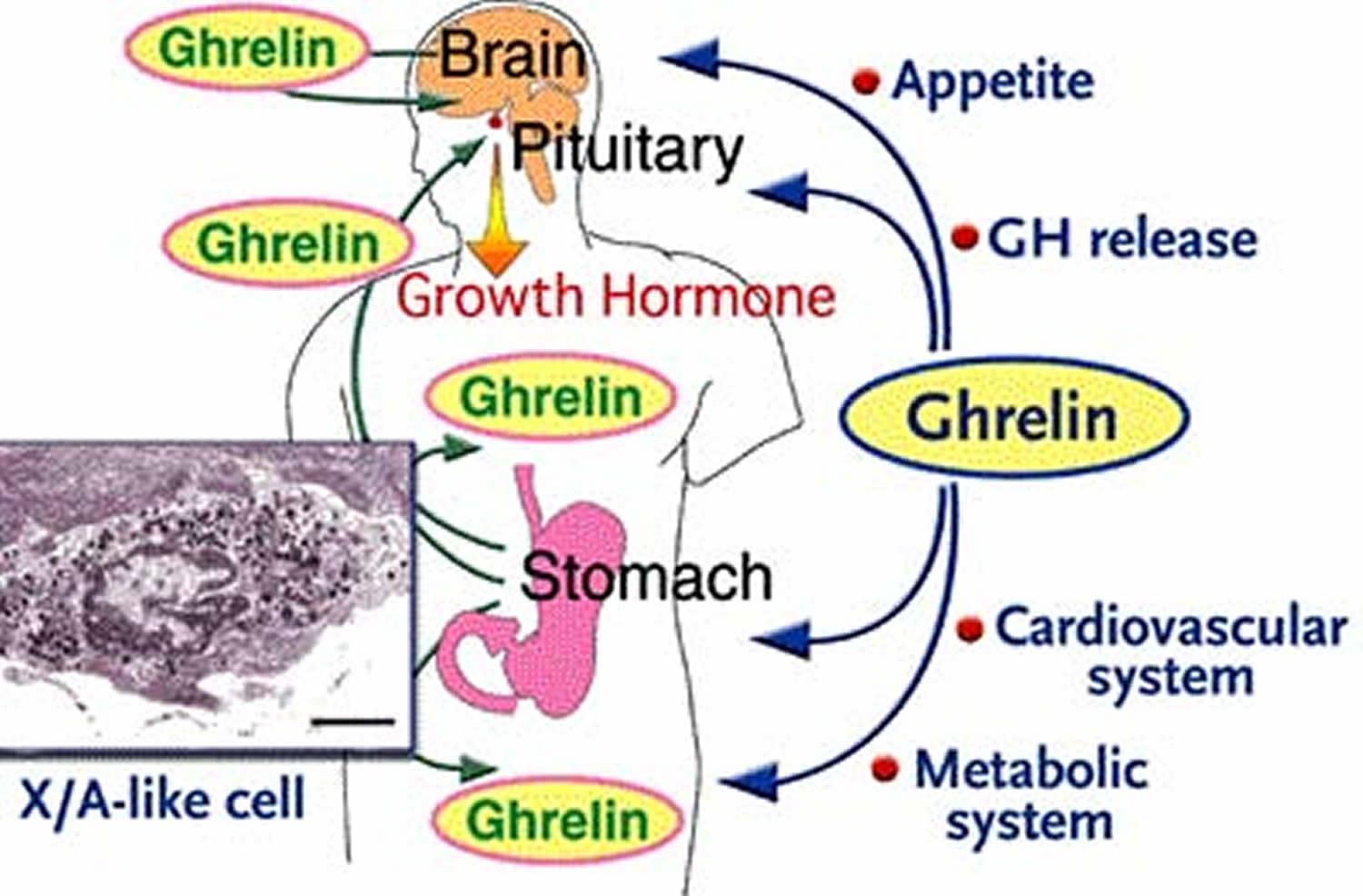
Ghrelin, is the only established appetite stimulant gut peptide to date, may be one factor involved in increased appetite, cravings and food intake following weight loss. Ghrelin levels, which increase following diet-induced weight loss and stimulate appetite, may contribute to weight regain 28. Ghrelin injections induce stress and anxiety-like responses in rats, and stress stimulates ghrelin secretion in both rats and humans 29, 30, 31. As stress can trigger overeating 32, ghrelin may be one mechanism by which appetite and food intake increase during stress. Indeed, individuals attempting to lose weight often cite life stressors as reasons for abandoning diet plans, and following strict diet/exercise programmes is a stressful endeavour. Rigidly restricting food intake often leads to compensatory overeating and psychological distress associated with diet-breaking may further increase disinhibited eating 33. Recent research has highlighted relationships between ghrelin, stress and lifestyle factors 34. Stress-induced increases in ghrelin, which further stimulate appetite, might impede efforts to maintain weight loss. Interventions to prevent increased ghrelin levels in response to stress and weight loss are needed 35.
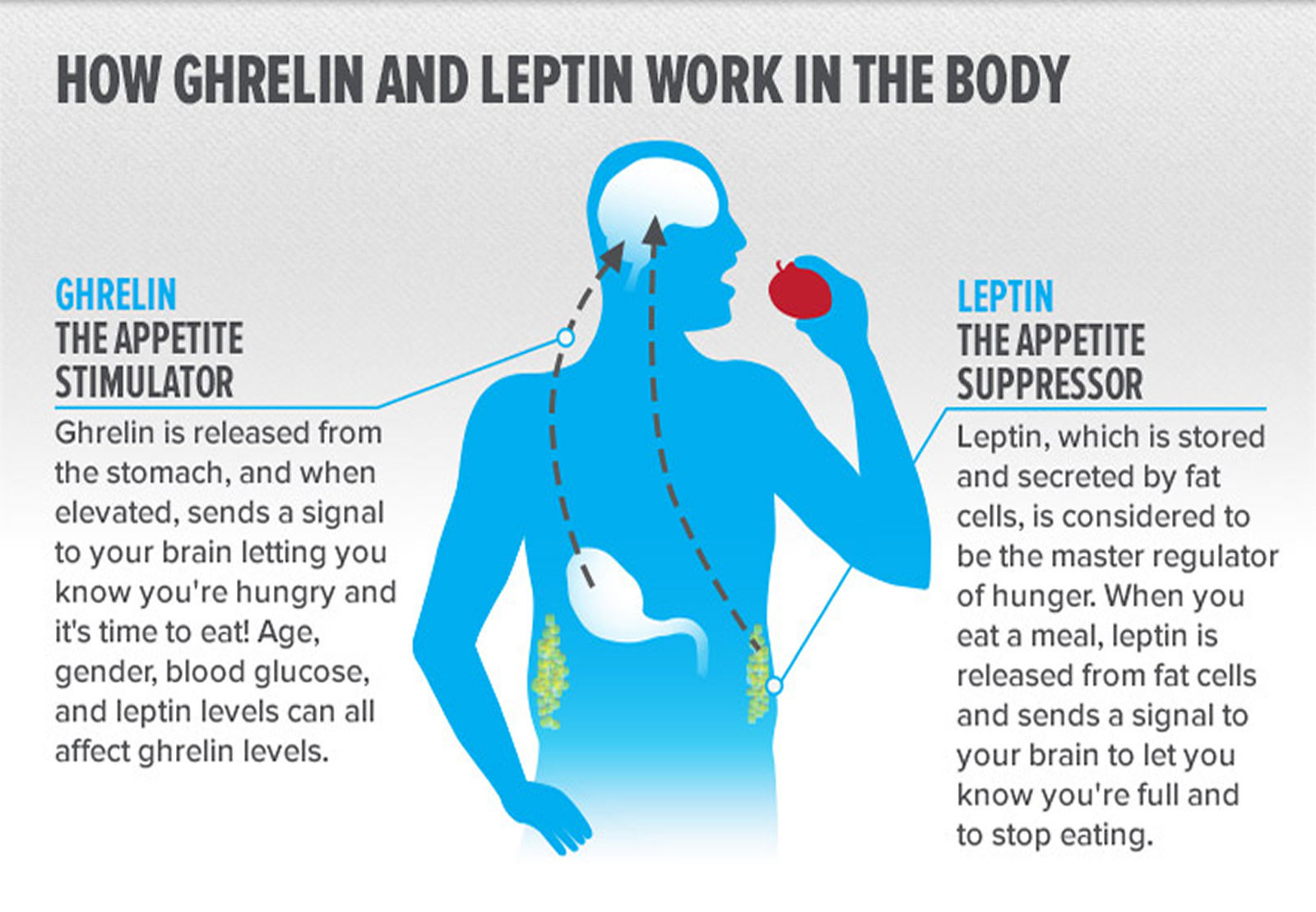
Ghrelin and Leptin
Leptin and ghrelin are two hormones that have been recognized to have a major influence on energy balance. Ghrelin is a fast-acting “go” hormone that tells you to eat, seemingly playing a role in meal initiation. Ghrelin is considered a classical orexigenic (appetite stimulant) peptide hormone since its plasma levels rise before and decrease after ingestion of food in animals and humans 36. Leptin on the other hand is the hormone that tells you to stop eating and thereby inducing weight loss.
After rapid passage through the oral cavity and the esophagus, ingested food has contact with the stomach first and consecutively with parts of the small intestine. Along this way, specialized endocrine cells respond with the release of gut peptide hormones, many of which play a prime role in modulating nutrient intake. Most of them induce satiation (meal termination) and/or satiety (postponing next meal initiation) while ghrelin is the only known gut orexigenic (appetite stimulant) hormone.
More ghrelin and less leptin set the stage for weight regain. Known as the hunger hormone, ghrelin is produced in your hypothalamus (located in your brain), kidneys, and pituitary gland, but most of it is synthesized in and released by the stomach, and it is released in a pulsatile manner throughout the night, peaking when you wake. No matter where it’s produced, it always has the same effect: snack attack.
You see, ghrelin both induces hunger and is also secreted by it. Your stomach produces ghrelin when it’s feeling empty. Whenever you start producing ghrelin, you’ll feel hungry—and anytime you get hungry, ghrelin is secreted. From an evolutionary standpoint, these changes are supposed to prevent starvation when food is hard to come by. But in an environment in which high-calorie processed food is abundant and physical activity is largely unnecessary, the high rate of relapse after weight loss is not surprising.
Now, here’s the important thing: your ghrelin secretion schedule largely follows your eating schedule, because ghrelin is what we might call a trainable hormone—the more often you eat, the more often you’ll produce ghrelin.
This is one of the many reasons why the multiple-meal hypothesis—the one that states you need to eat five to six mini-meals per day—is flawed. This style of eating—from a hormonal standpoint and in light of the way our bodies are built—is designed to make you hungrier. This might be great if your goal is to gain weight. In that case, you want to be hungrier. But if losing weight and becoming leaner are your goals, training your body to eat less frequently, while still feeling full, is a more sustainable and enjoyable approach to eating without feeling frustrated or constantly hungry.
A better meal schedule will help you balance your ghrelin levels, and so will sleep. Research published in the American Journal of Human Biology found a direct link between a lack of sleep, overeating, and obesity. Many people think the reason less sleep leads to more eating is because when you sleep less, you’re awake longer; the more hours you’re up, the more time you have to eat. The real reason is that a lack of sleep impacts hormone levels and brain functioning in a way that pushes you toward more foods—and in particular the crap you should avoid.
It’s a three-pronged attack designed to make you fat. When you don’t sleep enough:
- Cortisol levels rise, which activates reward centers in your brain that make you crave food.
- You produce more ghrelin and it’s harder to decrease the levels, meaning you feel hungry all the freaking time.
- The lack of sleep and higher levels of ghrelin appear to make you more likely to grab for dessert foods and fattening, sugary snacks.
While cheat foods will have their place, the goal is for you to eat them when you want—not because your brain creates a desire that you can’t resist. But that’s exactly what happens when you don’t get enough sleep. When researchers from Columbia University used MRI testing on sleep-deprived participants, they found that the areas of the brain that desire junk food were more activated with less than six hours of sleep. So unless you want to be a mind-control experiment that is at the beck and call of every fast food commercial on television, your best protection is ensuring more rest.
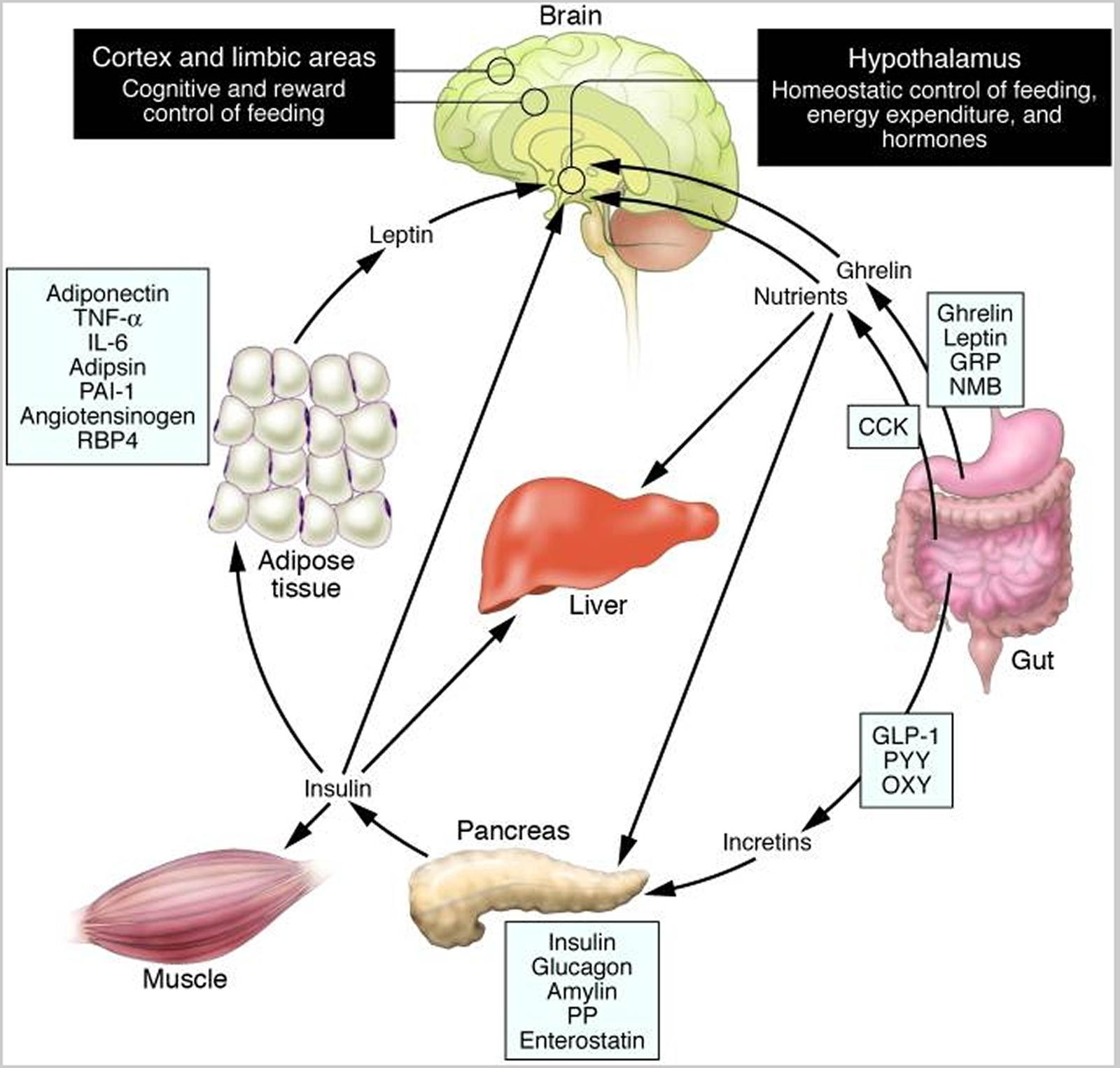
Schematic illustration of peptides secreted by the gut and adipose tissue (fat) that control energy balance. (Source 37)).
Note: Ghrelin is produced in the stomach and is a potent stimulator of appetite in the brain. In addition to increasing the uptake of nutrients by muscle, liver, and fat, insulin acts in the brain to suppress food intake. Gut-derived peptides such as GLP-1 augment insulin release from the pancreas. Leptin levels decline during weight loss and signal to the hypothalamus to stimulate feeding, reduce energy expenditure, and promote weight regain. As Rosenbaum et al. demonstrate in this issue of the The Journal of Clinical Investigation 38, low leptin levels during weight loss also increase the activity of brain areas involved in the decision-making and reward aspects of eating behavior. Thus, preventing the decline of leptin levels during weight loss by hormone replacement may be a means of overriding the homeostatic and behavioral tendencies toward energy conservation and weight regain during dieting. CCK, cholecystokinin; GLP-1, glucagon-like peptide–1; GRP, gastrin-releasing peptide; NMB, neuromedin B; OXY, oxyntomodulin; PAI-1, plasminogen activator inhibitor 1; PP, pancreatic polypeptide; PYY, peptide YY; RBP4, retinol-binding protein–4.
Here is How to Reduce Ghrelin Level
Food intake can have significant effects on circulating leptin and ghrelin levels. Overfeeding results in an increase in adipocyte leptin expression and circulating leptin in healthy human subjects 39, 40. Fasting (for 20 or 36 h or 3 days) results in a decrease of adipocyte leptin mRNA and serum leptin levels, with a greater decline in leptin levels in lean subjects than in obese subjects 41, 42, 43. Refeeding is again associated with a rise in serum leptin levels, and leptin levels return to baseline after 24 h 43, 41. On the other hand, fasting results in an increase in plasma ghrelin levels, with a nearly twofold increase immediately before each meal 44, 45. This preprandial increase in ghrelin levels correlates with hunger scores in humans 46. Feeding results in a decrease in plasma ghrelin levels within 1 to 2 h 44, 47.
Not only the size and frequency of meals have an effect on circulating leptin and ghrelin levels, but also the composition of a meal is a determinant of leptin and ghrelin levels in humans (Table 1). For example, low-fat/high-carbohydrate meals result in an increase in circulating leptin concentrations, which is larger, compared with high-fat/low-carbohydrate meals 48. In addition, high-fat meals lower 24-h circulating leptin levels relative to high-carbohydrate meals 49. Hydrolysed guar fibre or protein intake does not seem to have influence on circulating leptin concentrations 50, 51.
Table 1. Effects of diet composition on circulating leptin and ghrelin levels
| Diet | Effect on circulating leptin | Effect on circulating ghrelin |
|---|---|---|
| ||
| High-fat | 24-h circulating leptin levels ↓ relative to high-carbohydrate meal | ↓ |
| High-carbohydrate | ↓ (Larger drop compared with high-fat diet) | |
| Low-fat/high-carbohydrate | ↑(Larger compared with high-fat/low-carbohydrate meal) | No increase |
| High-fat/low-carbohydrate | ↑ | |
| Protein | No effect | Conflicting results |
| Hydrolysed guar fibre | No effect | |
| Non-caloric Psyllian fibres | ↓ | |
A low-fat diet seems to have an inhibitory effect on ghrelin levels, as one study reported that a low-fat/high-carbohydrate diet resulted in weight loss, without an increase in plasma ghrelin levels 52. Another study demonstrated that a high-carbohydrate diet caused a larger drop in ghrelin levels than a high-fat diet in healthy women 53. The effect of protein ingestion on ghrelin levels gives conflicting results 51, 54, 55. Finally, the use of non-caloric Psyllian fibres results in a decrease of plasma ghrelin levels in healthy women 56. Together, these data indicate that for obese subjects it is important to follow a specific diet in order to regulate food intake and body weight.
- Ensure enough sleep.
- Avoid eating too much sugar.
- Don’t eat too often.
- Eat Low Fat/high carb (high GI) diet.
- Eat more fiber.
- Get Physically Active.
- Eat more protein.
- Practice intermittent fasting.
Ghrelin, Anxiety and Stress
Several studies suggest that the peptide hormone ghrelin mediates some of the usual behavioral responses to acute and chronic stress 57. Circulating ghrelin levels have been found to rise following stress. It has been proposed that this elevated ghrelin helps animals cope with stress by generating antidepressant-like behavioral adaptations, although another study suggests that decreasing central nervous system ghrelin expression has antidepressant-like effects. Ghrelin also seems to have effects on anxiety, although these have been shown to be alternatively anxiogenic or anxiolytic. As an example, in a large cross sectional epidemiological U.S. study, a body mass index ≥30 was found to be associated with a 25% higher rate of mood disorders 58, 59. Conversely, other studies suggest that psychological stress can increase the risk of developing obesity. For instance, a longitudinal study found that major depression in late adolescent girls was associated with a 2.3-fold increased risk of obesity in adulthood 60. Also, a review of U.S. veterans with posttraumatic stress disorder showed a significantly increased rate of obesity 61. The mechanisms by which ghrelin affects mood-related behaviors have not yet been fully elucidated, but likely include interaction with its receptors in one or more brain sites critical to mood determination 57.
Ghrelin appears to increase anxiety via its actions on the (corticotropin-releasing hormone) CRH/ACTH (adrenocorticotropin hormone) system. Consistent with animal research, Takaya et al. 62 reported that intravenous injections of synthetic human ghrelin increased ACTH (adrenocorticotropin hormone) and cortisol (often called the stress hormone, cortisol principal steroid hormone produced by the adrenal cortex, regulates carbohydrate metabolism and the immune system and maintains blood pressure). Similarly, Arvat and colleagues 63 found that injecting ghrelin increased ACTH and cortisol.
Using the Trier Social Stress Test (5 min of performing a speech and 5 min of mental calculation in front of three observers), Rouach et al. 31 observed that among participants whose cortisol levels increased in response to stress, ghrelin levels increased. These participants also reported higher stress and anxiety after the TSST. Self-reported urges to eat did not differ between participants in upper and lower quartiles of ghrelin levels. However, the participants who reported an increased urge to overeat after the TSST had marginally higher ghrelin levels than participants who reported no change in urge to overeat. This highlights the importance of individual differences in the ghrelin response to stress.
The study on the effects of food deprivation, leptin and ghrelin upon noradrenaline release in the hypothalamic paraventricular nucleus and plasma adrenocorticotropic hormone (ACTH) concentrations after stressful stimuli. These data suggest that leptin inhibits and ghrelin facilitates neuroendocrine stress responses via noradrenaline release and indicate that a decrease in leptin and an increase in ghrelin release after food deprivation might contribute to augmentation of stress-induced ACTH release in a fasting state 64.
Summary of Ghrelin on Anxiety and Stress
Limited research is available on relationships between ghrelin and stress in humans. Existing research suggests that injecting ghrelin leads to increased stress hormone secretion. Due to the limited number of studies, small sample sizes and inconsistent results, research on the relationship between stressful life events and ghrelin is less clear. However, specific behavioural techniques designed to reduce ghrelin (e.g. increasing sleep and exercise), added to traditional weight loss maintenance protocols, might help maintain lower ghrelin levels and appetite while body weight stabilizes. A ghrelin-reduction programme might include stress management techniques such as time management, relaxation (e.g. diaphragmatic breathing, progressive muscle relaxation), cognitive restructuring, meditation and biofeedback 65.
Ghrelin and Exercise
Research on exercise and ghrelin in humans can be divided into three categories: acute aerobic, acute resistance and chronic exercise 66.
- Ghrelin and Acute aerobic exercise
Short-term aerobic exercise does not appear to affect total ghrelin 67, 68. Erdmann and colleagues 69 found that total ghrelin levels rose after low-intensity exercise but did not change after high-intensity exercise. Total ghrelin levels were not affected by the duration of exercise.
Broom et al. 70 found that acylated ghrelin levels decreased after 1 h of treadmill running. Ghrelin levels were lower in the exercise group, compared with controls, for at least 9 h after exercise. In addition, acylated ghrelin levels were directly related to self-reported hunger. Consistent with this notion, Marzullo et al. 71 found that acute aerobic exercise reduced acylated ghrelin but did not affect total ghrelin. This study also revealed that the acylated ghrelin-suppressing effect of exercise was even stronger in obese than in normal-weight participants.
However, discrepancies still exist; Ueda et al. 72 reported that acute aerobic exercise did not produce significant changes in acylated ghrelin. Notably though, acute exercise did lead to reduced post-exercise food intake.
In addition to direct effects of exercise on ghrelin and appetite, exercise might also influence ghrelin responses to meals. In one study, 1 h of cycling led to lower total ghrelin levels after exercise as well as after a meal 73.
- Ghrelin and Acute resistance exercise
Ghanbari-Niaki 74 found that a single episode of resistance exercise decreased immunoreactive ghrelin immediately following exercise and increased ghrelin 24 h after exercise. Ballard et al. 75 reported that one session of resistance exercise reduced total ghrelin as well as food intake. Additional research 76 investigating resistance exercise differentiated between concentric (lifting weights) and eccentric (lowering weights) muscle contractions. There were no changes in total ghrelin after eccentric contraction exercises; however, total ghrelin decreased after concentric exercises. However, Takano and colleagues 77 found no changes in ghrelin within 30 min after short-term low-intensity resistance exercise.
- Ghrelin and Chronic exercise
Results of several studies suggest that chronic exercise leads to increased total ghrelin levels, specifically when participants lose weight 78, 79. Ravussin and colleagues 80 reported that cycling twice daily for 93 days (designed to induce negative energy balance resulting in average weight loss of 5 kg) resulted in a non-significant increase in total ghrelin. Leidy and colleagues 81 reported that among women who lost weight in a 3-month diet and exercise programme, total ghrelin increased and was positively correlated with amount of weight lost. Among exercisers who did not lose weight; however, ghrelin did not increase.
In their study of effects of a 12-week exercise programme on ghrelin among overweight Korean boys, Kim et al. 82 assessed fasting total and acylated ghrelin levels. Participants showed decreases in body mass index (BMI) and per cent body fat, and non-acylated ghrelin levels increased and were inversely related to weight loss and decreased body fat percentage. However, acylated ghrelin did not change, and acylated ghrelin levels were comparable between experimental and control groups throughout treatment. Kim et al. proposed that different effects of food deprivation and exercise on appetite and feeding may be due to the different effects of acylated vs. non-acylated ghrelin. Specifically, this exercise intervention increased non-acylated ghrelin (decreasing energy intake) without affecting acylated ghrelin (not enhancing appetite). Similarly, Jones et al. 83 found that among overweight adolescents, although 8 months of aerobic exercise reduced per cent body fat, active ghrelin levels did not change.
Summary of Ghrelin and Exercise
Short-term aerobic exercise does not appear to affect ghrelin 35. However, there is some evidence that resistance exercise decreases ghrelin. Most studies on long-term exercise show that total ghrelin increases as participants lose weight; however, there is some evidence that long-term exercise prevents weight loss-induced increases in acylated ghrelin 35.
Ghrelin and Sleep
In humans, sleep deprivation has been associated with higher ghrelin levels, increased hunger and higher BMI 84, 85. Even a single night of sleep deprivation appears sufficient to increase ghrelin and hunger 86. Spiegel et al. 87 found that restricting sleep increased ghrelin as well as hunger and appetite, especially for calorie-dense, high-carbohydrate foods. This increase in hunger was directly related to the increase in ghrelin-to-leptin ratio. Schüssler et al. 88 found that sleep deprivation led to increased plasma total ghrelin levels and postulated that increased ghrelin levels in response to sleep deprivation are adaptive in that they promote sleep.
Consistent with this notion, Weikel et al. 89 found that ghrelin injections led to increased slow-wave sleep (especially Stage IV sleep) and less REM sleep during the second third of the night. Similarly, Kluge et al. 90 found that peripheral injections of acylated ghrelin led to decreased REM sleep and increased non-REM sleep.
Whereas ghrelin levels are stimulated by sleep deprivation and promote slow-wave sleep, ghrelin appears to decrease in response to improved sleep. Takahashi et al. 91 investigated the effects of continuous positive airway pressure treatment for sleep apnoea on ghrelin. Before treatment, the obstructive sleep apnoea (OSA) groups had higher BMIs and ghrelin levels than non-obstructive sleep apnoea healthy controls. After 1 month of treatment, the obstructive sleep apnoea treatment group had reduced acylated ghrelin, decreased ghrelin was directly related to improvements in respiratory disturbance, and obstructive sleep apnoea patients’ ghrelin were comparable with controls. Among overweight sleep apnoea patients, improving breathing at night (and thus improving sleep quality) may decrease acylated ghrelin.
Kimata 92 successfully influenced ghrelin levels and sleep using a behavioural intervention. Children with atopic dermatitis (and comorbid night-time wakening) and healthy children were presented with either a humorous film or a control film before bedtime. Watching the humorous film decreased night-time wakening and nocturnal salivary ghrelin levels in children with atopic dermatitis. Neither of the films affected sleep or ghrelin in the healthy children, who did not have difficulties with sleeping at baseline.
- Ghrelin, sleep and eating
Rosenhagen et al. 93 reported that a 27-year-old woman with Night-Eating syndrome had elevated ghrelin levels. In a study of 11 healthy, normal-weight participants who each slept 5.5 h in a sleep laboratory on one night and 8.5 h on another night, shorter sleep duration led to higher caloric intake (specifically from snacking between meals) during the day 94.
Summary of Ghrelin and Sleep
In animal studies, ghrelin increases wakefulness and decreases REM sleep. However, research with humans suggests that injecting ghrelin increases slow-wave sleep and decreases REM sleep. In addition, sleep deprivation stimulates ghrelin secretion and improved sleep reduces ghrelin. Taken together, ghrelin seems to increase in response to sleep deprivation (and perhaps then act as a sleep-promoting agent) and decrease in response to improved sleep quality in humans. Finally, ghrelin may be involved in problematic night-time eating.
More research is needed on relationships between ghrelin, sleep and eating behaviour. Although there is emerging consensus that ghrelin rises in response to sleep deprivation and may be related to increased snacking, future research should investigate whether increases in ghrelin either partially or fully mediate the relationship between sleep deprivation and eating.
Conclusion
Ghrelin has profound appetite stimulant, fat forming (adipogenic), and growth hormone like properties, increasing food intake and body weight. Secreted predominantly from the stomach, ghrelin is the natural ligand for the growth hormone secretagogue receptor in the pituitary gland, thus fulfilling criteria of a brain-gut peptide. However, the wide tissue distribution of ghrelin suggests that it may have other functions as well. Innovative strategies for suppressing ghrelin and decreasing appetite during weight loss maintenance are needed.
It is now established that obese patients are leptin-resistant and endogenous ghrelin is a potentially important new regulator of the complex systems controlling food intake and body weight. However, the manner in which both the leptin and ghrelin systems contribute to the development or maintenance of obesity is as yet not clear.
- Sakata I, Sakai T. Ghrelin Cells in the Gastrointestinal Tract. International Journal of Peptides. 2010;2010:945056. doi:10.1155/2010/945056. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2925405/[↩][↩][↩]
- Appetite regulation and energy balance. Erlanson-Albertsson C. Acta Paediatr Suppl. 2005 Jun; 94(448):40-1. https://www.ncbi.nlm.nih.gov/pubmed/16175806/[↩]
- Hosoda H, Kojima M, Matsuo H, Kengawa K. Ghrelin and des-acyl ghrelin: two major forms of rat ghrelin peptide in gastrointestinal tissue. Biochem Biophys Res Commun 2000; 279: 909–913. https://www.ncbi.nlm.nih.gov/pubmed/11162448[↩]
- Asakawa A, Inui A, Fujimiya M, Sakamaki R, Shinfuku N, Ueta Y, Meguid MM, Kasuga M. Stomach regulates energy balance via acylated ghrelin and desacyl ghrelin. Gut 2005; 54: 18–24. https://www.ncbi.nlm.nih.gov/pubmed/15591499[↩]
- Yang J, Zhao T, Goldstein JL, Brown MS. Inhibition of ghrelin O-acyltransferase (GOAT) by octanoylated pentapeptides. Proc Natl Acad Sci 2008; 10750–10755. https://www.ncbi.nlm.nih.gov/pubmed/18669668[↩]
- Kojima M, Hosoda H, Date Y, Nakazato M, Kangawa K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999; 402: 656–660. https://www.ncbi.nlm.nih.gov/pubmed/10604470[↩]
- Yin Y, Li Y, Zhang W. The Growth Hormone Secretagogue Receptor: Its Intracellular Signaling and Regulation. International Journal of Molecular Sciences. 2014;15(3):4837-4855. doi:10.3390/ijms15034837. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3975427/[↩]
- Gnanapavan S, Kola B, Bustin SA, Morris DG, McGee P, Fairclough P, Bhattacharya S, Carpenter R, Grossman AB, Korbonits M. The tissue distribution of the mRNA of ghrelin and subtypes of its receptor, GHS-R, in humans. J. Clin. Endocrinol. Metab. 2002;87(6):2988. https://www.ncbi.nlm.nih.gov/pubmed/12050285[↩]
- Howard AD, Feighner SD, Cully DF, Arena JP, Liberator PA, Rosenblum CI, Hamelin M, Hreniuk DL, Palyha OC, Anderson J, Paress PS, Diaz C, Chou M, Liu KK, McKee KK, Pong SS, Chaung LY, Elbrecht A, Dashkevicz M, Heavens R, Rigby M, Sirinathsinghji DJ, Dean DC, Melillo DG, Patchett AA, Nargund R, Griffin PR, DeMartino JA, Gupta SK, Schaeffer JM, Smith RG, Van der Ploeg LH. A receptor in pituitary and hypothalamus that functions in growth hormone release. Science. 1996;273(5277):974–977. https://www.ncbi.nlm.nih.gov/pubmed/8688086[↩]
- Guan XM, Yu H, Palyha OC, McKee KK, Feighner SD, Sirinathsinghji DJ, Smith RG, Van der Ploeg LH, Howard AD. Distribution of mRNA encoding the growth hormone secretagogue receptor in brain and peripheral tissues. Brain Res. Mol. Brain Res. 1997;48(1):23–29. https://www.ncbi.nlm.nih.gov/pubmed/9379845[↩]
- Nakazato M, Murakami N, Date Y, Kojima M, Matsuo H, Kangawa K, Matsukura S. A role for ghrelin in the central regulation of feeding. Nature. 2001;409(6817):194–198. https://www.ncbi.nlm.nih.gov/pubmed/11196643[↩]
- Hattori N, Saito T, Yagyu T, Jiang BH, Kitagawa K, Inagaki C. GH, GH receptor, GH secretagogue receptor, and ghrelin expression in human T cells, B cells, and neutrophils. J. Clin. Endocrinol. Metab. 2001;86(9):4284–4291. https://www.ncbi.nlm.nih.gov/pubmed/11549663[↩]
- Schellekens H, Dinan TG, Cryan JF. Lean Mean Fat Reducing “Ghrelin” Machine: Hypothalamic Ghrelin and Ghrelin Receptors as Therapeutic Targets in Obesity. Neuropharmacology. 2010;58(1):2–16. https://www.ncbi.nlm.nih.gov/pubmed/19573543[↩]
- Zigman JM, Jones JE, Lee CE, Saper CB, Elmquist JK. Expression of ghrelin receptor mRNA in the rat and the mouse brain. J. Comp. Neurol. 2006;494(3):528–548. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4524499/[↩]
- Shuto Y, Shibasaki T, Wada K, Parhar I, Kamegai J, Sugihara H, Oikawa S, Wakabayashi I. Generation of polyclonal antiserum against the growth hormone secretagogue receptor (GHS-R): evidence that the GHS-R exists in the hypothalamus, pituitary and stomach of rats. Life Sci. 2001;68(9):991–996. https://www.ncbi.nlm.nih.gov/pubmed/11212874[↩]
- Mondal MS, Date Y, Yamaguchi H, Toshinai K, Tsuruta T, Kangawa K, Nakazato M. Identification of ghrelin and its receptor in neurons of the rat arcuate nucleus. Regul. Pept. 2005;126(1-2):55–59. https://www.ncbi.nlm.nih.gov/pubmed/15620414[↩]
- Biological, physiological, and pharmacological aspects of ghrelin. Hosoda H, Kojima M, Kangawa K. J Pharmacol Sci. 2006; 100(5):398-410. https://www.ncbi.nlm.nih.gov/pubmed/16612045/[↩][↩]
- Ghrelin stimulation of growth hormone release and appetite is mediated through the growth hormone secretagogue receptor. Sun Y, Wang P, Zheng H, Smith RG. Proc Natl Acad Sci U S A. 2004 Mar 30; 101(13):4679-84. https://www.ncbi.nlm.nih.gov/pubmed/15070777/[↩]
- Ghrelin enhances gastric emptying in diabetic gastroparesis: a double blind, placebo controlled, crossover study. Murray CD, Martin NM, Patterson M, Taylor SA, Ghatei MA, Kamm MA, Johnston C, Bloom SR, Emmanuel AV. Gut. 2005 Dec; 54(12):1693-8. https://www.ncbi.nlm.nih.gov/pubmed/16085693/[↩]
- Ghrelin is present in pancreatic alpha-cells of humans and rats and stimulates insulin secretion. Date Y, Nakazato M, Hashiguchi S, Dezaki K, Mondal MS, Hosoda H, Kojima M, Kangawa K, Arima T, Matsuo H, Yada T, Matsukura S. Diabetes. 2002 Jan; 51(1):124-9. https://www.ncbi.nlm.nih.gov/pubmed/11756331/[↩]
- The mitogenic and antiapoptotic actions of ghrelin in 3T3-L1 adipocytes. Kim MS, Yoon CY, Jang PG, Park YJ, Shin CS, Park HS, Ryu JW, Pak YK, Park JY, Lee KU, Kim SY, Lee HK, Kim YB, Park KS. Mol Endocrinol. 2004 Sep; 18(9):2291-301. https://www.ncbi.nlm.nih.gov/pubmed/15178745/[↩]
- Ghrelin enhances the growth of cultured human adrenal zona glomerulosa cells by exerting MAPK-mediated proliferogenic and antiapoptotic effects. Mazzocchi G, Neri G, Rucinski M, Rebuffat P, Spinazzi R, Malendowicz LK, Nussdorfer GG. Peptides. 2004 Aug; 25(8):1269-77. https://www.ncbi.nlm.nih.gov/pubmed/15350694/[↩]
- Developments in ghrelin biology and potential clinical relevance. Smith RG, Jiang H, Sun Y. Trends Endocrinol Metab. 2005 Nov; 16(9):436-42. https://www.ncbi.nlm.nih.gov/pubmed/16213742/[↩]
- Altered myocardial expression of ghrelin and its receptor (GHSR-1a) in patients with severe heart failure. Beiras-Fernandez A, Kreth S, Weis F, Ledderose C, Pöttinger T, Dieguez C, Beiras A, Reichart B. Peptides. 2010 Dec; 31(12):2222-8. https://www.ncbi.nlm.nih.gov/pubmed/20804798/[↩]
- Baldanzi G., Filigheddu N., Cutrupi S., Catapano F., Bonissoni S., Fubini A., Malan D., Baj G., Granata R., Broglio F., et al. Ghrelin and des-acyl ghrelin inhibit cell death in cardiomyocytes and endothelial cells through ERK1/2 and PI 3-kinase/AKT. J. Cell Biol. 2002;159:1029–1037. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2173981/[↩]
- Ghrelin amplifies dopamine signaling by cross talk involving formation of growth hormone secretagogue receptor/dopamine receptor subtype 1 heterodimers. Jiang H, Betancourt L, Smith RG. Mol Endocrinol. 2006 Aug; 20(8):1772-85. https://www.ncbi.nlm.nih.gov/pubmed/16601073/[↩]
- Ghrelin O-acyltransferase (GOAT) is essential for growth hormone-mediated survival of calorie-restricted mice. Zhao TJ, Liang G, Li RL, Xie X, Sleeman MW, Murphy AJ, Valenzuela DM, Yancopoulos GD, Goldstein JL, Brown MS. Proc Natl Acad Sci U S A. 2010 Apr 20; 107(16):7467-72. https://www.ncbi.nlm.nih.gov/pubmed/20231469/[↩]
- Cummings DE, Weigle DS, Frayo RS, Breen PA, Ma MK, Dellinger EP, Purnell JQ. Plasma ghrelin levels after diet-induced weight loss or gastric bypass surgery. N Engl J Med 2002; 346: 1623–1630. https://www.ncbi.nlm.nih.gov/pubmed/12023994[↩]
- Carlini VP, Monzon ME, Varas MM, Cragnolini AB, Schioth HB, Scimonelli TN, de Barioglio SR. Ghrelin increases anxiety-like behavior and memory retention in rats. Biochem Biophys Res Commun 2002; 299: 739–743. https://www.ncbi.nlm.nih.gov/pubmed/12470640[↩]
- Kawakami A, Okada N, Rokkaku K, Honda K, Ishibashi S, Onaka T. Leptin inhibits and ghrelin augments hypothalamic noradrenaline release after stress. Stress 2008; 11: 363–369. https://www.ncbi.nlm.nih.gov/pubmed/18800308[↩]
- Rouach V, Bloch M, Rosenberg N, Gilad S, Limor R, Stern N, Greenman Y. The acute ghrelin response to a psychological stress challenge does not predict the post-stress urge to eat. Psychoneuroendocrinology 2007; 32: 693–702. https://www.ncbi.nlm.nih.gov/pubmed/17560728[↩][↩]
- Greeno CG, Wing RR. Stress-induced eating. Psychol Bull 1994; 115: 444–464. https://www.ncbi.nlm.nih.gov/pubmed/8016287[↩]
- Adams CE, Leary MR. Promoting self-compassionate attitudes toward eating among restrictive and guilty eaters. J Soc Clin Psychol 2007; 26: 1120–1144.[↩]
- Obesity Reviews Volume 12, Issue 5, May 2011; Pages e211–e218. Lifestyle factors and ghrelin: critical review and implications for weight loss maintenance. http://onlinelibrary.wiley.com/doi/10.1111/j.1467-789X.2010.00776.x/full[↩]
- Obesity Reviews Volume 12, Issue 5, May 2011. Pages e211–e218. Lifestyle factors and ghrelin: critical review and implications for weight loss maintenance. http://onlinelibrary.wiley.com/doi/10.1111/j.1467-789X.2010.00776.x/full[↩][↩][↩]
- A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Cummings DE, Purnell JQ, Frayo RS, Schmidova K, Wisse BE, Weigle DS. Diabetes. 2001 Aug; 50(8):1714-9. https://www.ncbi.nlm.nih.gov/pubmed/11473029/[↩]
- Gastroenterology 2007. 132(Issue 6[↩]
- Rosenbaum M., Sy M., Pavlovich K., Leibel R.L., Hirsch J. Leptin reverses weight loss–induced changes in regional neural activity responses to visual food stimuli. J. Clin. Invest. 2008;118:2583–2591. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2430499/[↩]
- Kolaczynski JW, Ohannesian JP, Considine RV, Marco CC, Caro JF. Response of leptin to short-term and prolonged overfeeding in humans. J Clin Endocrinol Metab 1996; 81: 4162–4165. https://www.ncbi.nlm.nih.gov/pubmed/8923877[↩]
- Levine JA, Eberhardt NL, Jensen MD. Leptin responses to overfeeding: relationship with body fat and nonexercise activity thermogenesis. J Clin Endocrinol Metab 1999; 84: 2751–2754. https://www.ncbi.nlm.nih.gov/pubmed/10443673[↩]
- Kolaczynski JW, Considine RV, Ohannesian J, Marco C, Opentanova I, Nyce MR, Myint M, Caro JF. Responses of leptin to short-term fasting and refeeding in humans: a link with ketogenesis but not ketones themselves. Diabetes 1996; 45: 1511–1515. https://www.ncbi.nlm.nih.gov/pubmed/8866554[↩][↩]
- Korbonits M, Trainer PJ, Little JA, Edwards R, Kopelman PG, Besser GM, Svec F, Grossman AB. Leptin levels do not change acutely with food administration in normal or obese subjects, but are negatively correlated with pituitary-adrenal activity. Clin Endocrinol 1997; 46: 751–757. https://www.ncbi.nlm.nih.gov/pubmed/9274707[↩]
- Weigle DS, Duell PB, Connor WE, Steiner RA, Soules MR, Kuijper JL. Effect of fasting, refeeding, and dietary fat restriction on plasma leptin levels. J Clin Endocrinol Metab 1997; 82: 561–565. https://www.ncbi.nlm.nih.gov/pubmed/9024254[↩][↩]
- Cummings DE, Purnell JQ, Frayo RS, Schmidova K, Wisse BE, Weigle DS. A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes 2001; 50: 1714–1719. https://www.ncbi.nlm.nih.gov/pubmed/11473029[↩][↩]
- Ariyasu H, Takaya K, Tagami T, Ogawa Y, Hosoda K, Akamizu T, Suda M, Koh T, Natsui K, Toyooka S, Shirakami G, Usui T, Shimatsu A, Doi K, Hosoda H, Kojima M, Kangawa K, Nakao K. Stomach is a major source of circulating ghrelin, and feeding state determines plasma ghrelin-like immunoreactivity levels in humans. J Clin Endocrinol Metab 2001; 86: 4753–4758. https://www.ncbi.nlm.nih.gov/pubmed/11600536[↩]
- Cummings DE, Frayo RS, Marmonier C, Aubert R, Chapelot D. Plasma ghrelin levels and hunger scores in humans initiating meals voluntarily without time- and food-related cues. Am J Physiol Endocrinol Metab 2004; 287: E297–E304. https://www.ncbi.nlm.nih.gov/pubmed/15039149[↩]
- Tschop M, Wawarta R, Riepl RL, Friedrich S, Bidlingmaier M, Landgraf R, Folwaczny C. Post-prandial decrease of circulating human ghrelin levels. J Endocrinol Invest 2001; 24: RC19–RC21. https://www.ncbi.nlm.nih.gov/pubmed/11434675[↩]
- Havel PJ, Townsend R, Chaump L, Teff K. High-fat meals reduce 24-h circulating leptin concentrations in women. Diabetes 1999; 48: 334–341. https://www.ncbi.nlm.nih.gov/pubmed/10334310[↩]
- Havel PJ. Role of adipose tissue in body-weight regulation: mechanisms regulating leptin production and energy balance. Proc Nutr Soc 2000; 59: 359–371. https://www.ncbi.nlm.nih.gov/pubmed/10997652[↩]
- Heini AF, Lara-Castro C, Schneider H, Kirk KA, Considine RV, Weinsier RL. Effect of hydrolyzed guar fiber on fasting and postprandial satiety and satiety hormones: a double-blind, placebo-controlled trial during controlled weight loss. Int J Obes Relat Metab Disord 1998; 22: 906–909. https://www.ncbi.nlm.nih.gov/pubmed/9756250[↩]
- Groschl M, Knerr I, Topf HG, Schmid P, Rascher W, Rauh M. Endocrine responses to the oral ingestion of a physiological dose of essential amino acids in humans. J Endocrinol 2003; 179: 237–244. https://www.ncbi.nlm.nih.gov/pubmed/14596675[↩][↩]
- Weigle DS, Cummings DE, Newby PD, Breen PA, Frayo RS, Matthys CC, Callahan HS, Purnell JQ. Roles of leptin and ghrelin in the loss of body weight caused by a low fat, high carbohydrate diet. J Clin Endocrinol Metab 2003; 88: 1577–1586. https://www.ncbi.nlm.nih.gov/pubmed/12679442[↩]
- Monteleone P, Bencivenga R, Longobardi N, Serritella C, Maj M. Differential responses of circulating ghrelin to high-fat or high-carbohydrate meal in healthy women. J Clin Endocrinol Metab 2003; 88: 5510–5514. https://www.ncbi.nlm.nih.gov/pubmed/14602798[↩]
- Erdmann J, Lippl F, Schusdziarra V. Differential effect of protein and fat on plasma ghrelin levels in man. Regul Pept 2003; 116: 101–107. https://www.ncbi.nlm.nih.gov/pubmed/14599721[↩]
- Greenman Y, Golani N, Gilad S, Yaron M, Limor R, Stern N. Ghrelin secretion is modulated in a nutrient- and gender-specific manner. Clin Endocrinol 2004; 60: 382–388. https://www.ncbi.nlm.nih.gov/pubmed/15009005[↩]
- Nedvidkova J, Krykorkova I, Bartak V, Papezova H, Gold PW, Alesci S, Pacak K. Loss of meal-induced decrease in plasma ghrelin levels in patients with anorexia nervosa. J Clin Endocrinol Metab 2003; 88: 1678–1682. https://www.ncbi.nlm.nih.gov/pubmed/12679456[↩]
- Chuang J-C, Zigman JM. Ghrelin’s Roles in Stress, Mood, and Anxiety Regulation. International Journal of Peptides. 2010;2010:460549. doi:10.1155/2010/460549. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2915752/[↩][↩]
- Simon GE, Von Korff M, Saunders K, et al. Association between obesity and psychiatric disorders in the US adult population. Archives of General Psychiatry. 2006;63(7):824–830. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1913935/[↩]
- Kloiber S, Ising M, Reppermund S, et al. Overweight and obesity affect treatment response in major depression. Biological Psychiatry. 2007;62(4):321–326. https://www.ncbi.nlm.nih.gov/pubmed/17241618[↩]
- Richardson LP, Davis R, Poulton R, et al. A longitudinal evaluation of adolescent depression and adult obesity. Archives of Pediatrics and Adolescent Medicine. 2003;157(8):739–745. https://www.ncbi.nlm.nih.gov/pubmed/12912778[↩]
- Vieweg WV, Julius DA, Benesek J, et al. Posttraumatic stress disorder and body mass index in military veterans. Preliminary findings. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 2006;30(6):1150–1154. https://www.ncbi.nlm.nih.gov/pubmed/16647795[↩]
- Takaya K, Ariyasu H, Kanamoto N, Iwakura H, Yoshimoto A, Harada M, Mori K, Komatsu Y, Usui T, Shimatsu A, Ogawa Y, Hosoda K, Akamizu T, Kojima M, Kangawa K, Nakao K. Ghrelin strongly stimulates growth hormone (GH) release in humans. J Clin Endocrinol Metab 2000; 85: 4908–4911. https://www.ncbi.nlm.nih.gov/pubmed/11134161[↩]
- Arvat E, Maccario M, Di Vito L, Broglio F, Benso A, Gottero C, Papotti M, Muccioli G, Dieguez C, Casanueva FF, Deghenghi R, Camanni F, Ghigo E. Endocrine activities of ghrelin, a natural growth hormone secretagogue (GHS), in humans: comparison and interactions with hexarelin, a nonnatural peptidyl GHS, and GH-releasing hormone. J Clin Endocrinol Metab 2001; 86: 1169–1174. https://www.ncbi.nlm.nih.gov/pubmed/11238504[↩]
- The International Journal on the Biology of Stress. Published online: 07 Jul 2009. Volume 11, 2008 – Issue 5. Leptin inhibits and ghrelin augments hypothalamic noradrenaline release after stress. http://www.tandfonline.com/doi/citedby/10.1080/10253890701820257[↩]
- Obesity Reviews Volume 12, Issue 5, May 2011. Pages e211–e218. http://onlinelibrary.wiley.com/doi/10.1111/j.1467-789X.2010.00776.x/full[↩]
- Kraemer RR, Castracane VD. Exercise and humoral mediators of peripheral energy balance: ghrelin and adiponectin. Exp Biol Med 2007; 232: 184–194. https://www.ncbi.nlm.nih.gov/pubmed/17259325[↩]
- Kraemer RR, Durand RJ, Acevefo EO, Johnson LG, Kraemer GR, Hebert EP, Castracane VD. Rigorous running increases growth hormone and insulin-like growth factor-I without altering ghrelin. Exp Biol Med 2004; 229: 240–246. https://www.ncbi.nlm.nih.gov/pubmed/14988516[↩]
- Burns SF, Broom DR, Miyashita M, Mundy C, Stensel DJ. A single session of treadmill running has no effect on plasma total ghrelin concentrations. J Sports Sci 2007; 25: 635–642. https://www.ncbi.nlm.nih.gov/pubmed/17454530[↩]
- Erdmann J, Tahbaz R, Lippl F, Wagenpfeil S, Schusdziarra V. Plasma ghrelin levels during exercise – effects of intensity and duration. Regul Pept 2007; 143: 127–135. https://www.ncbi.nlm.nih.gov/pubmed/17570540[↩]
- Broom DR, Stensel DJ, Bishop NC, Burns SF, Miyashita M. Exercise-induced suppression of acylated ghrelin in humans. J Appl Physiol 2007; 102: 2165–2171. https://www.ncbi.nlm.nih.gov/pubmed/17347386[↩]
- Marzullo P, Salvadori A, Brunani A, Verti B, Walker GE, Fanari P, Tovaglieri I, de Medici CD, Savia G, Liuzzi A. Acylated ghrelin decreases during acute exercise in the lean and obese state. Clin Endocrinol 2008; 69: 970–971. https://www.ncbi.nlm.nih.gov/pubmed/18419786[↩]
- Ueda S, Yoshikawa T, Katsura Y, Usui T, Nakao H, Fujimoto S. Changes in gut hormone levels and negative energy balance during aerobic exercise in obese young males. J Endocrinol 2009; 201: 151–159. https://www.ncbi.nlm.nih.gov/pubmed/19158129[↩]
- Malkova D, McLaughlin R, Manthou E, Wallace AM, Nimmo MA. Effect of moderate-intensity exercise session on preprandial and postprandial responses of circulating ghrelin and appetite. Horm Metab Res 2008; 40: 410–415. https://www.ncbi.nlm.nih.gov/pubmed/18401836[↩]
- Ghanbari-Niaki A. Ghrelin and glucoregulatory hormone responses to a single circuit resistance exercise in male college students. Clin Biochem 2006; 39: 966–970. https://www.ncbi.nlm.nih.gov/pubmed/16979150[↩]
- Ballard TP, Melby CL, Camus H, Cianciulli M, Pitts J, Schmidt S, Hickey MS. Effect of resistance exercise, with or without carbohydrate supplementation, on plasma ghrelin concentrations and postexercise hunger and food intake. Metabolism 2009; 58: 1191–1199. https://www.ncbi.nlm.nih.gov/pubmed/19497597[↩]
- Kraemer RR, Durand RJ, Hollander DB, Tryniecki JL, Hebert EP, Castracane VD. Ghrelin and other glucoregulatory hormone responses to eccentric and concentric muscle contractions. Endocrine 2004; 24: 93–98. https://www.ncbi.nlm.nih.gov/pubmed/15249708[↩]
- Takano H, Morita T, Iida H, Asada K, Kato M, Uno K, Hirose K, Matsumoto A, Takenaka K, Hirata Y, Eto F, Nagai R, Sato Y, Nakajima T. Hemodynamic and hormonal responses to a short-term low-intensity resistance exercise with the reduction of muscle blood flow. Eur J Appl Physiol 2005; 95: 65–73. https://www.ncbi.nlm.nih.gov/pubmed/15959798[↩]
- Santosa S, Demonty I, Lichtenstein AH, Cianflone K, Jones PJH. An investigation of hormone and lipid associations after weight loss in women. J Am Coll Nutr 2007; 26: 250–258. https://www.ncbi.nlm.nih.gov/pubmed/17634170[↩]
- Littman AJ, Vitiello MV, Foster-Schubert K, Ulrich CM, Tworoger SS, Potter JD, Weigle DS, McTiernan A. Sleep, ghrelin, leptin and changes in body weight during a 1-year moderate-intensity physical activity intervention. Int J Obes 2007; 31: 466–475. https://www.ncbi.nlm.nih.gov/pubmed/16909130[↩]
- Ravussin E, Tschop M, Morales S, Bouchard C, Heiman ML. Plasma ghrelin concentration and energy balance: overfeeding and negative energy balance studies in twins. J Clin Endocrinol Metab 2001; 86: 4547–4551. https://www.ncbi.nlm.nih.gov/pubmed/11549706[↩]
- Leidy HJ, Gardner JK, Frye BR, Snook ML, Schuchert MK, Richard EL, Williams NI. Circulating ghrelin is sensitive to changes in body weight during a diet and exercise program in normal-weight young women. J Clin Endocrinol Metab 2004; 89: 2659–2664. https://www.ncbi.nlm.nih.gov/pubmed/15181038[↩]
- Kim HJ, Lee S, Kim TW, Kim HH, Jeon YJ, Yoon YS, Oh SW, Kwak H, Lee JG. Effects of exercise-induced weight loss on acylated and non-acylated ghrelin in overweight children. Clin Endocrinol 2008; 68: 416–422. https://www.ncbi.nlm.nih.gov/pubmed/17888020[↩]
- Jones TE, Basilio JL, Brophy PM, McCammon MR, Hickner RC. Long-term exercise training in overweight adolescents improves plasma peptide YY and resistin. Obesity 2009; 17: 1189–1195. https://www.ncbi.nlm.nih.gov/pubmed/19247279[↩]
- Taheri S, Lin L, Austin D, Young T, Mignot E. Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index. PLoS Med 2004; 1: e62. https://www.ncbi.nlm.nih.gov/pubmed/15602591[↩]
- Spiegel K, Tasali E, Penev P, Cauter EV. Brief Communication: Sleep Curtailment in Healthy Young Men Is Associated with Decreased Leptin Levels, Elevated Ghrelin Levels, and Increased Hunger and Appetite. Ann Intern Med. 2004;141:846-850. doi: 10.7326/0003-4819-141-11-200412070-00008. http://annals.org/aim/article/717987/brief-communication-sleep-curtailment-healthy-young-men-associated-decreased-leptin[↩]
- Schmid SM, Hallschmid M, Jauch-Chara K, Born J, Schultes B. A single night of sleep deprivation increases ghrelin levels and feelings of hunger in normal-weight healthy men. J Sleep Res 2008; 17: 331–334. https://www.ncbi.nlm.nih.gov/pubmed/18564298[↩]
- Spiegel K, Tasali E, Penev P, Van Cauter E. Brief communication: sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite. Ann Intern Med 2004; 141: 846–851. https://www.ncbi.nlm.nih.gov/pubmed/15583226[↩]
- Schüssler P, Uhr M, Ising M, Weikel JC, Schmid DA, Held K, Mathias S, Steiger A. Nocturnal ghrelin, ACTH, GH and cortisol secretion after sleep deprivation in humans. Psychoneuroendocrinology 2006; 31: 915–923. https://www.ncbi.nlm.nih.gov/pubmed/16814473[↩]
- Weikel JC, Wichniak A, Ising M, Brunner H, Friess E, Held K, Mathias S, Schmid DA, Uhr M, Steiger A. Ghrelin promotes slow-wave sleep in humans. Am J Physiol Endocrinol Metab 2002; 284: E407–E415. https://www.ncbi.nlm.nih.gov/pubmed/12388174[↩]
- Kluge M, Schüssler P, Bleninger P, Kleyer S, Uhr M, Weikel JC, Yassouridis A, Zuber V, Steiger A. Ghrelin alone or co-administered with GHRH or CRH increases non-REM sleep and decreases REM sleep in young males. Psychoneuroendocrinology 2008; 33: 497–506. https://www.ncbi.nlm.nih.gov/pubmed/18329818[↩]
- Takahashi K, Chin K, Akamizu T, Morita S, Sumi K, Oga T, Matsumoto H, Niimi A, Tsuboi T, Fukuhara S, Kangawa K, Mishima M. Acylated ghrelin level in patients with OSA before and after nasal CPAP treatment. Respirology 2008; 13: 810–816. https://www.ncbi.nlm.nih.gov/pubmed/18811879[↩]
- Kimata H. Viewing humorous film improves night-time wakening in children with atopic dermatitis. Indian Pediatr 2007; 44: 281–285. https://www.ncbi.nlm.nih.gov/pubmed/17468523[↩]
- Rosenhagen MC, Uhr M, Schüssler P, Steiger A. Elevated plasma ghrelin levels in night-eating syndrome. Am J Psychiatry 2005; 162: 813. https://www.ncbi.nlm.nih.gov/pubmed/15800168[↩]
- Nedeltcheva AV, Kilkus JM, Imperial J, Kasza K, Schoeller DA, Penev PD. Sleep curtailment is accompanied by increased intake of calories from snacks. Am J Clin Nutr 2009; 89: 126–133. https://www.ncbi.nlm.nih.gov/pubmed/19056602[↩]


{kind=link}