What is secretin
Secretin is a hormone that was initially identified by Bayliss and Starling in 1902 1. It was known that the pancreas secreted digestive juices in response to the passage of food (chyme) through the pyloric sphincter into the duodenum. Bayliss and Starling discovered (by cutting all the nerves to the pancreas in their experimental animals) that this process was not, in fact, governed by the nervous system. They determined that a substance secreted by the intestinal lining stimulates the pancreas after being transported via the bloodstream. They named this intestinal secretion secretin 1. Secretin was the first such “chemical messenger” identified. This type of substance is now called a hormone, a term coined by Bayliss in 1905 2. Secretin is a 27 amino acid basic peptide produced by the secretin-containing S-cells of the duodenal epithelium and in smaller amount in the jejunum of the small intestine 3 and secretin is released by acid delivered into the duodenum 4. Secretin is released into the blood when duodenal pH drops below 4 5. Secretin hormone acts to neutralize the pH of the duodenum by stimulating water and bicarbonate secretion into the duodenum. This aids in the digestion process as pancreatic amylase and lipase require a certain pH to function optimally.
Prosecretin is a precursor to secretin, which is present in digestion. Secretin is stored in this unusable form, and is activated by gastric hydrochloric acid in the lower intestine to neutralize the pH and ensure no damage is done to the small intestine by the gastric acid 6.
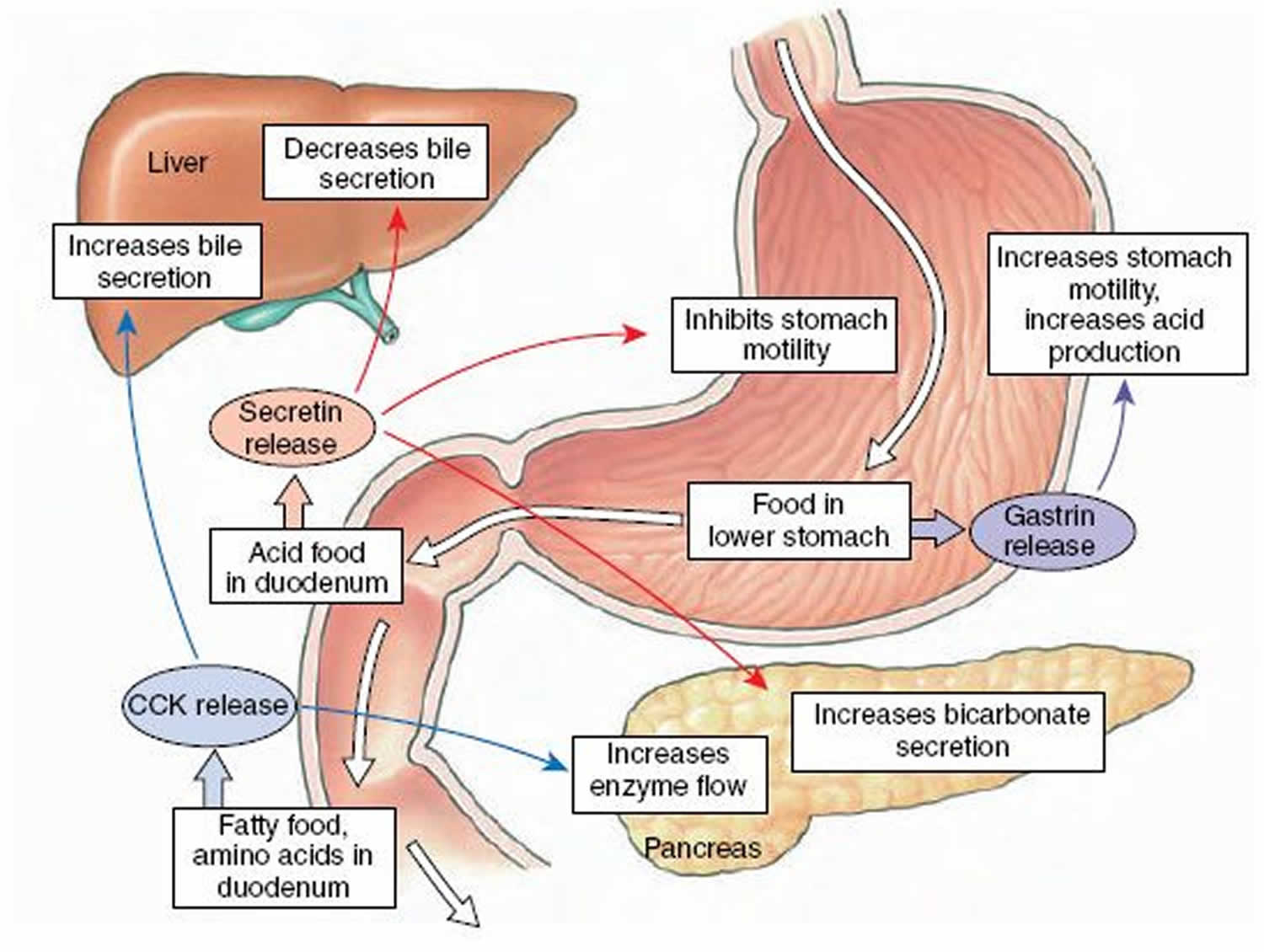
The quantity of secretin released into the blood and the volume of pancreatic secretion are directly related to the load of titratable acid delivered to the duodenum 7. The role of secretin in meal-stimulated pancreatic fluid and bicarbonate secretion has been confirmed by showing that immune-neutralization of secretin with specific anti-secretin antibody decreases these responses by as much as 80% 8. The anti-secretin antibody also inhibits meal-stimulated enzyme secretion by as much as 50%, suggesting that secretin also has a role in enzyme secretion, possibly by potentiating the action of agonists, such as acetylcholine 7. These findings are consistent with those described earlier, showing that gastric acid stimulated during the cephalic phase that enters the duodenum increases fluid and bicarbonate secretion and augments pancreatic digestive enzyme secretion 9. The results indicate that secretin plays key roles in regulating both ductal and acinar components of exocrine pancreatic secretion during a meal. Secretin release is inhibited by H2 antagonists, which reduce gastric acid secretion. As a result, if the pH in the duodenum increases above 4.5, secretin cannot be released 10.
Of note, the bicarbonate response to secretin also depends on cholinergic input because atropine partially inhibits pancreatic bicarbonate secretion stimulated by intra-duodenal application of acid without an effect on the increase in blood concentration of secretin that occurs with the acid instillation 11. Furthermore, if exogenous secretin is infused to reproduce the plasma concentrations of secretin during a meal, the pancreatic bicarbonate output is less than the bicarbonate output observed with a meal. These results indicate that the bicarbonate and fluid response during a meal is mediated by a combination of cholinergic and hormonal secretin actions on the pancreatic ductal system.
The pancreatic enzyme secretory response during the intestinal phase is mediated by both neural and hormonal pathways. Both removing the vagus nerve and administration of the cholinergic antagonist atropine markedly inhibit the digestive enzyme (and bicarbonate) responses to low intestinal loads of amino acids and fatty acids as well as infusion of physiologic concentrations of the hormone cholecystokinin (CCK) 12. These results indicate a prominent role for the cholinergic nervous system utilizing vagovagal reflexes in the regulation of digestive enzyme secretion during the intestinal phase. Further, the results confirm a role for the cholinergic nervous system in bicarbonate secretion stimulated by secretin 7.
Similar to the hormone cholecystokinin (CCK)-releasing factors, secretin-releasing factors have been described to regulate the release of secretin from the secretin-containing enteroendocrine S cell of the intestine 13. Phospholipase A2 represents one secretin-releasing factor 7. Phospholipase A2 is present in the mucosa of the upper small intestine and is released into the duodenum with intestinal acidification, and it causes secretin release into the circulation and stimulates a bicarbonate-rich pancreatic secretion with application to the upper small intestinal lumen. These results provide support for a secretin-releasing factor 7.
Secretin shares structural similarity with glucagon, gastric inhibitory polypeptide, vasoactive intestinal polypeptide, and growth hormone-releasing hormone (GHRH). Secretin is a potent stimulus for bicarbonate secretion. Secretin also stimulates secretion of bile, release of insulin, and release of gastric pepsin in the stomach. Secretin inhibits glucagon release, intestinal motility, and prevents the uptake of water and sodium ions by the intestine. In normal patients, secretin has little effect on gastrin levels, but stimulates gastrin greatly in Zollinger-Ellison patients. Secretin is also elevated in Zollinger-Ellison patients and in patients with duodenal ulcer. Secretin levels are low in patients with pernicious anemia and achlorhydria. Secretin secretion can be suppressed by somatostatin, cimetidine, and methionine-enkephalin.
In the past century, the research of secretin has gone by many milestones, which includes isolation, purification, structural characterization, and chemical synthesis of secretin, establishment of its hormonal status, identification of the specific receptor, cloning of secretin and its receptor genes, and identification of secretin-releasing peptides. Secretin has been identified as a hormone-regulating pancreatic exocrine secretion of fluid and bicarbonate, gastric acid secretion, and gastric motility 14.
There are a few observations showing the involvement of secretin in the regulation of gastrointestinal mucosal growth so far. Treatment with cholecystokinin (CCK) and secretin is shown to prevent atrophy in the jejunum and ileum of dogs given total parenteral nutrition (TPN) as a sole nutrient source 15. Generally, secretin is shown to inhibit the trophic action of gastrin but it has no a direct antitrophic activity in the gastrointestinal mucosa 16. Secretin inhibits the gastrin-mediated stimulation of DNA synthesis in the gastric oxyntic gland region, duodenum, and colon 17 . This inhibitory effect of secretin is independent of its ability to inhibit gastrin-stimulated acid secretion. It is likely that secretin indirectly regulates gastrointestinal mucosal growth by blocking the trophic effect of gastrin 18.
Where is secretin produced?
Secretin is a 27 amino acid basic peptide produced by the secretin-containing S-cells of the duodenal epithelium and in smaller amount in the jejunum of the small intestine 3 and secretin is released by acid delivered into the duodenum 4
Secretin function
Secretin is a potent stimulus for bicarbonate secretion. Secretin hormone acts to neutralize the pH of the duodenum by stimulating water and bicarbonate secretion into the duodenum. This aids in the digestion process as pancreatic amylase and lipase require a certain pH to function optimally. Secretin also stimulates secretion of bile, release of insulin, and release of gastric pepsin in the stomach. Secretin inhibits glucagon release, intestinal motility, and prevents the uptake of water and sodium ions by the intestine. In normal patients, secretin has little effect on gastrin levels, but stimulates gastrin greatly in Zollinger-Ellison patients. Secretin is also elevated in Zollinger-Ellison patients and in patients with duodenal ulcer. Secretin levels are low in patients with pernicious anemia and achlorhydria. Secretin secretion can be suppressed by somatostatin, cimetidine, and methionine-enkephalin.
There are a few observations showing the involvement of secretin in the regulation of gastrointestinal mucosal growth so far. Treatment with cholecystokinin (CCK) and secretin is shown to prevent atrophy in the jejunum and ileum of dogs given total parenteral nutrition (TPN) as a sole nutrient source 15. Generally, secretin is shown to inhibit the trophic action of gastrin but it has no a direct antitrophic activity in the gastrointestinal mucosa 16. Secretin inhibits the gastrin-mediated stimulation of DNA synthesis in the gastric oxyntic gland region, duodenum, and colon 17 . This inhibitory effect of secretin is independent of its ability to inhibit gastrin-stimulated acid secretion. It is likely that secretin indirectly regulates gastrointestinal mucosal growth by blocking the trophic effect of gastrin 18.
Secretin was also recently discovered to play a role in osmoregulation by acting on the hypothalamus, pituitary gland, and kidney 19.
pH regulation
Secretin primarily functions to neutralize the pH in the duodenum, allowing digestive enzymes from the pancreas (e.g., pancreatic amylase and pancreatic lipase) to function optimally 20.
Secretin targets the pancreas; pancreatic centroacinar cells have secretin receptors in their plasma membrane. As secretin binds to these receptors, it stimulates adenylate cyclase activity and converts ATP to cyclic AMP 21. Cyclic AMP acts as second messenger in intracellular signal transduction and causes the organ to secrete a bicarbonate-rich fluid that flows into the intestine. Bicarbonate is a base that neutralizes the acid, thus establishing a pH favorable to the action of other digestive enzymes in the small intestine 22.
Secretin also increases water and bicarbonate secretion from duodenal Brunner’s glands to buffer the incoming protons of the acidic chyme 20. and also reduces acid secretion by parietal cells of the stomach 23. It does this through at least three mechanisms: 1) By stimulating release of somatostatin, 2) By inhibiting release of gastrin in the pyloric antrum, and 3) By direct downregulation of the parietal cell acid secretory mechanics 24.
It counteracts blood glucose concentration spikes by triggering increased insulin release from pancreas, following oral glucose intake 25.
Osmoregulation
Secretin modulates water and electrolyte transport in pancreatic duct cells 26, liver cholangiocytes 27 and epididymis epithelial cells 28, as part of the vasopressin-independent mechanisms in regulating renal water reabsorption 29.
Secretin is found in the magnocellular neurons of the paraventricular and supraoptic nuclei of the hypothalamus and along the neurohypophysial tract to neurohypophysis. During increased osmolality, it is released from the posterior pituitary. In the hypothalamus, it activates vasopressin release 30. Secretin is also needed to carry out the central effects of angiotensin 2. In the absence of secretin or its receptor in the gene knockout animals, central injection of angiotensin 2 was unable to stimulate water intake and vasopressin release 31.
It has been suggested that abnormalities in such secretin release could explain the abnormalities underlying type D syndrome of inappropriate antidiuretic hormone hypersecretion (SIADH) 32. In these individuals, vasopressin release and response are normal, although abnormal renal expression, translocation of aquaporin 2, or both are found 32.
Food intake
Secretin and its receptor are found in discrete nuclei of the hypothalamus, including the paraventricular nucleus and the arcuate nucleus, which are the primary brain sites for regulating body energy homeostasis. It was found that both central and peripheral injection of secretin reduce food intake in mouse, indicating an anorectic role of the peptide. This function of the peptide is mediated by the central melanocortin system 33.
Secretin test
Secretin is used in a diagnostic tests for pancreatic function; secretin is injected and the pancreatic output can then be imaged with magnetic resonance imaging, a noninvasive procedure, or secretions generated as a result can gathered either through an endoscope or through tubes inserted through the mouth, down into the duodenum 34.
- Secretin-stimulated ultrasound estimation of pancreatic secretion in cystic fibrosis validated by magnetic resonance imaging 35. A recombinant human secretin has been available since 2004 for these diagnostic purposes 36.
- Bayliss HP, Starling EH. Mechanism of pancreatic secretion. J Physiol Lond 28: pp. 325–53, 1902[↩][↩]
- Secretin and the exposition of hormonal control. J Physiol. 2004;560(Pt 2):339. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1665254/[↩]
- Immunofluorescent localization of secretin and enteroglucagon in human intestinal mucosa. Scand J Gastroenterol. 1971;6(8):739-44. https://www.ncbi.nlm.nih.gov/pubmed/4945081[↩][↩]
- Lopez PP, Bhimji SS. Anatomy, Abdomen and Pelvis, Duodenum. [Updated 2018 Sep 19]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2018 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK482390[↩][↩]
- Chey, W.Y., and Konturek, S.J., Plasma secretion and pancreatic secretion in response to liver extract meal with varied pH and exogenous secretin in the dog. J Physiol, 1982. 324: pp. 263–72.[↩]
- Gafvelin G, Jörnvall H, Mutt V. Processing of prosecretin: isolation of a secretin precursor from porcine intestine. Proc Natl Acad Sci U S A. 1990;87(17):6781-5. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC54621/pdf/pnas01042-0314.pdf[↩]
- Pandol SJ. The Exocrine Pancreas. San Rafael (CA): Morgan & Claypool Life Sciences; 2010. Regulation of Whole-Organ Pancreatic Secretion. Available from: https://www.ncbi.nlm.nih.gov/books/NBK54132[↩][↩][↩][↩][↩]
- Song, Y., et al., Canine pancreatic juice stimulates the release of secretin and pancreatic secretion in the dog. Am J Physiol, 1999. 277(3 Pt 1): pp. G731–5[↩]
- Anagnostides, A., et al., Sham feeding and pancreatic secretion. Evidence for direct vagal stimulation of enzyme output. Gastroenterology, 1984. 87(1): pp. 109–14.[↩]
- Plasma secretin concentrations and gastric pH in healthy subjects and patients with digestive diseases. Dig Dis Sci. 1981 Jul;26(7):591-7. https://www.ncbi.nlm.nih.gov/pubmed/7249893[↩]
- You, C.H., Rominger, J.M., and Chey, W.Y., Effects of atropine on the action and release of secretin in humans. Am J Physiol, 1982. 242(6): pp. G608–11[↩]
- Singer, M.V., Solomon, T.E., and Grossman, M.I., Effect of atropine on secretion from intact and transplanted pancreas in dog. Am J Physiol, 1980. 238(1): pp. G18–22.[↩]
- Li, J.P., et al., Pancreatic phospholipase A2 from the small intestine is a secretin-releasing factor in rats. Am J Physiol Gastrointest Liver Physiol, 2001. 281(2): pp. G526–32.[↩]
- William CY, Ta-Min C. Secretin, 100 years later. J Gastroenterol, 38: pp. 1025–35, 2003.[↩]
- Thomas RP, Hellmich MR, Townsend CM, Evers BM. Role of gastrointestinal hormones in the proliferation of normal and neoplastic tissues. Endocrine Rev 24: pp. 571–99, 2003.10.1210/er.2002-0028.[↩][↩]
- Johnson LR. Regulation of gastrointestinal mucosal growth. Physiol Rev 68: pp. 456–502, 1988.[↩][↩]
- Johnson LR, Guthrie PD. Secretin inhibition of gastrin-stimulated deoxyribonucleic acid synthesis. Gastroenterology 67: pp. 601–6, 1974.[↩][↩]
- Rao JN, Wang JY. Regulation of Gastrointestinal Mucosal Growth. San Rafael (CA): Morgan & Claypool Life Sciences; 2010. Role of GI Hormones on Gut Mucosal Growth. Available from: https://www.ncbi.nlm.nih.gov/books/NBK54093/[↩][↩]
- Chu JY, Chung SC, Lam AK, Tam S, Chung SK, Chow BK. Phenotypes developed in secretin receptor-null mice indicated a role for secretin in regulating renal water reabsorption. Mol Cell Biol. 2007;27(7):2499-511. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1899889/[↩]
- Hall, John E.; Guyton, Arthur C. (2006). Textbook of medical physiology. St. Louis, Mo: Elsevier Saunders. pp. 800–1. ISBN 0-7216-0240-1.[↩][↩]
- Gardner, JD (1978). “Receptors and gastrointestinal hormones”. In Sleisenger, MH; Fordtran, JS. Gastrointestinal Disease (2nd ed.). Philadelphia: WB Saunders Company. pp. 179–95.[↩]
- Osnes M, Hanssen LE, Flaten O, Myren J. Exocrine pancreatic secretion and immunoreactive secretin (IRS) release after intraduodenal instillation of bile in man. Gut. 1978;19(3):180-4. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1411891/pdf/gut00460-0032.pdf[↩]
- Palmer, KR; Penman, ID (2010). “Alimentary track and pancreatic disease”. In Colledge, NR; Walker, BR; Ralston, SH. Davidson’s Principles and Practice of Medicine (20th ed.). Edinburgh: Churchill Livingstone. p. 844. ISBN 0-7020-3085-6.[↩]
- Boron, Walter F.; Boulpaep, Emile L. (2012). “Acid secretion”. Medical Physiology (2nd ed.). Philadelphia: Saunders. p. 1352. ISBN 978-1-4377-1753-2.[↩]
- Kraegen EW, Chisholm DJ, Young JD, Lazarus L. The gastrointestinal stimulus to insulin release. II. A dual action of secretin. J Clin Invest. 1970;49(3):524-9. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC322500/pdf/jcinvest00219-0116.pdf[↩]
- Secretin causes H+/HCO3- secretion from pig pancreatic ductules by vacuolar-type H(+)-adenosine triphosphatase. Villanger O, Veel T, Raeder MG. Gastroenterology. 1995 Mar; 108(3):850-9.[↩]
- Agonist-induced coordinated trafficking of functionally related transport proteins for water and ions in cholangiocytes. Tietz PS, Marinelli RA, Chen XM, Huang B, Cohn J, Kole J, McNiven MA, Alper S, LaRusso NF. J Biol Chem. 2003 May 30; 278(22):20413-9.[↩]
- Secretin controls anion secretion in the rat epididymis in an autocrine/paracrine fashion. Chow BK, Cheung KH, Tsang EM, Leung MC, Lee SM, Wong PY. Biol Reprod. 2004 Jun; 70(6):1594-9.[↩]
- Phenotypes developed in secretin receptor-null mice indicated a role for secretin in regulating renal water reabsorption. Chu JY, Chung SC, Lam AK, Tam S, Chung SK, Chow BK. Mol Cell Biol. 2007 Apr; 27(7):2499-511.[↩]
- Chu JY, Lee LT, Lai CH, et al. Secretin as a neurohypophysial factor regulating body water homeostasis. Proc Natl Acad Sci U S A. 2009;106(37):15961-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2747226/ [↩]
- Lee VH, Lee LT, Chu JY, et al. An indispensable role of secretin in mediating the osmoregulatory functions of angiotensin II. FASEB J. 2010;24(12):5024-32. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2992369/[↩]
- Chu JY, Lee LT, Lai CH, et al. Secretin as a neurohypophysial factor regulating body water homeostasis. Proc Natl Acad Sci U S A. 2009;106(37):15961-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2747226/[↩][↩]
- Cheng CY, Chu JY, Chow BK. Central and peripheral administration of secretin inhibits food intake in mice through the activation of the melanocortin system. Neuropsychopharmacology. 2010;36(2):459-71. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3055665/[↩]
- Engjom T, Erchinger F, Lærum BN, et al. Diagnostic Accuracy of a Short Endoscopic Secretin Test in Patients With Cystic Fibrosis. Pancreas. 2015;44(8):1266-72. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4947543/[↩]
- Engjom T, Tjora E, Wathle G, et al. Secretin-stimulated ultrasound estimation of pancreatic secretion in cystic fibrosis validated by magnetic resonance imaging. Eur Radiol. 2017;28(4):1495-1503. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5834563/[↩]
- https://www.fda.gov/downloads/Drugs/GuidanceComplianceRegulatoryInformation/Guidances/UCM072008.pdf[↩]

{kind=link}