Contents
What is Luteolin
Luteolin (3′,4′,5,7-tetrahydroxy flavone) belongs to a group of naturally occurring compounds called flavonoids, a type of flavones or polyphenols, found in many plants, including fruits, vegetables, and herbs, that has a variety of potential health benefits with various studies reporting luteolin with anti-inflammatory, antioxidant, anticancer, cytoprotective, anti-microbial, cardioprotective, anti-diabetic, neuroprotective, and anti-allergic effects 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18. Luteolin, a flavonoid, is mainly found in various vegetables and fruits, including celery, apple skins, carrots, cabbages, onion leaves, parsley, cabbages, broccoli, peppers and chrysanthemum flowers 5.
Plants rich in luteolin have been used for a long time in Iranian, Brazilian, and Chinese traditional medicines to treat high blood pressure (hypertension), inflammatory diseases, and cancer 19, 1. For example, Zygophyllum simplex L. (commonly known as hureim or simple-leaved bean caper), Chrysanthemum indicum var. albescens (also known as white wild chrysanthemum), Cymbopogon citratus (commonly known as lemon grass), which all have high luteolin content, have been prescribed for inflammatory diseases, including gout, asthma, skin psoriasis, and erythema (a condition that causes the skin or mucous membranes to appear red) 1.
Luteolin is considered non-toxic. However, luteolin has not been granted Generally Recognized as Safe (GRAS) status by the U.S. Food and Drug Administration (FDA) 1. Currently, luteolin has been used as a dietary supplement and is also incorporated into cosmetic products, as demonstrated by its nontoxic side effects. Luteolin safety is supported by data showing that the oral median lethal dose (LD50, which is the amount of a substance that kills 50% of a group of test animals) in mice and rats exceeded 2500 and 5000 mg/kg, respectively, roughly translating to 219.8−793.7 mg/kg in humans 20. In animal and test tube cells investigations, luteolin demonstrated a lack of detrimental effects on healthy cells and did not result in noteworthy side effects, even at a relatively high concentration of 30 μM 21, 22. Research intended to examine luteolin’s ability to reverse changes caused by exposure to lead in rats showed no side effects at a dose of 50 mg/kg, oral, daily; therefore, the researchers concluded that luteolin could possibly be administered as a dietary supplement in humans 23. In research designed to examine the effectiveness and tolerability in white children with autism spectrum disorder of a dietary supplement containing two flavonoids (>95% pure), including luteolin (100 mg per 10 kg weight per day), for 26 weeks found transient irritability in subjects as the only side effect of treatment and a significant improvement in adaptive functioning as measured by using the Vineland Adaptive Behavior Scales age-equivalent scores 24.
Figure 1. Flavones
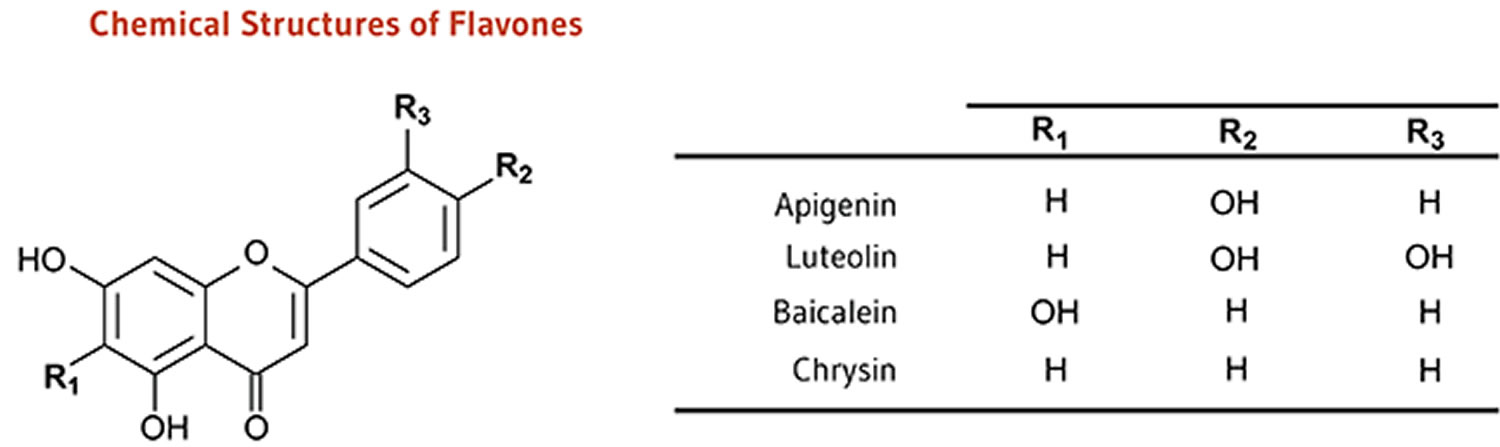
Footnotes: Flavones include luteolin, luteolin glucosides, apigenin, chrysin and rutin. Flavones like the other flavonoids, exert different their biological activities due to modifications in the chemical structure as hydroxylation, O-/C-glycosylation, O-methylation, and acylation 25. Structurally, luteolin has a hydroxyl (-OH) group attached at the 5-, 7-, 3′-, and 4′- positions of the flavone backbone structure. The chemical structure of luteolin is shown in Figure 2.
Figure 2. Luteolin chemical structure
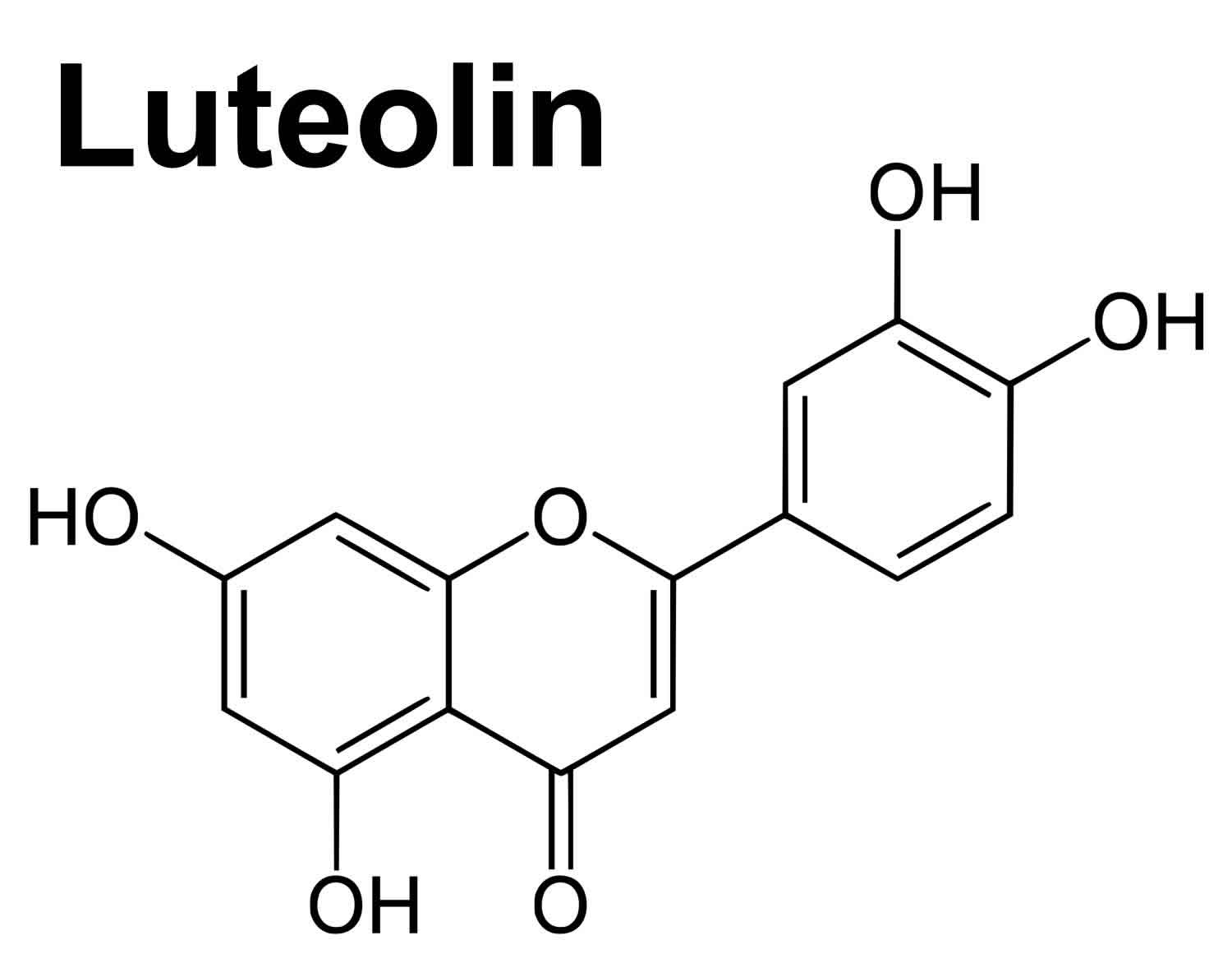
Figure 3. Luteolin biological activities
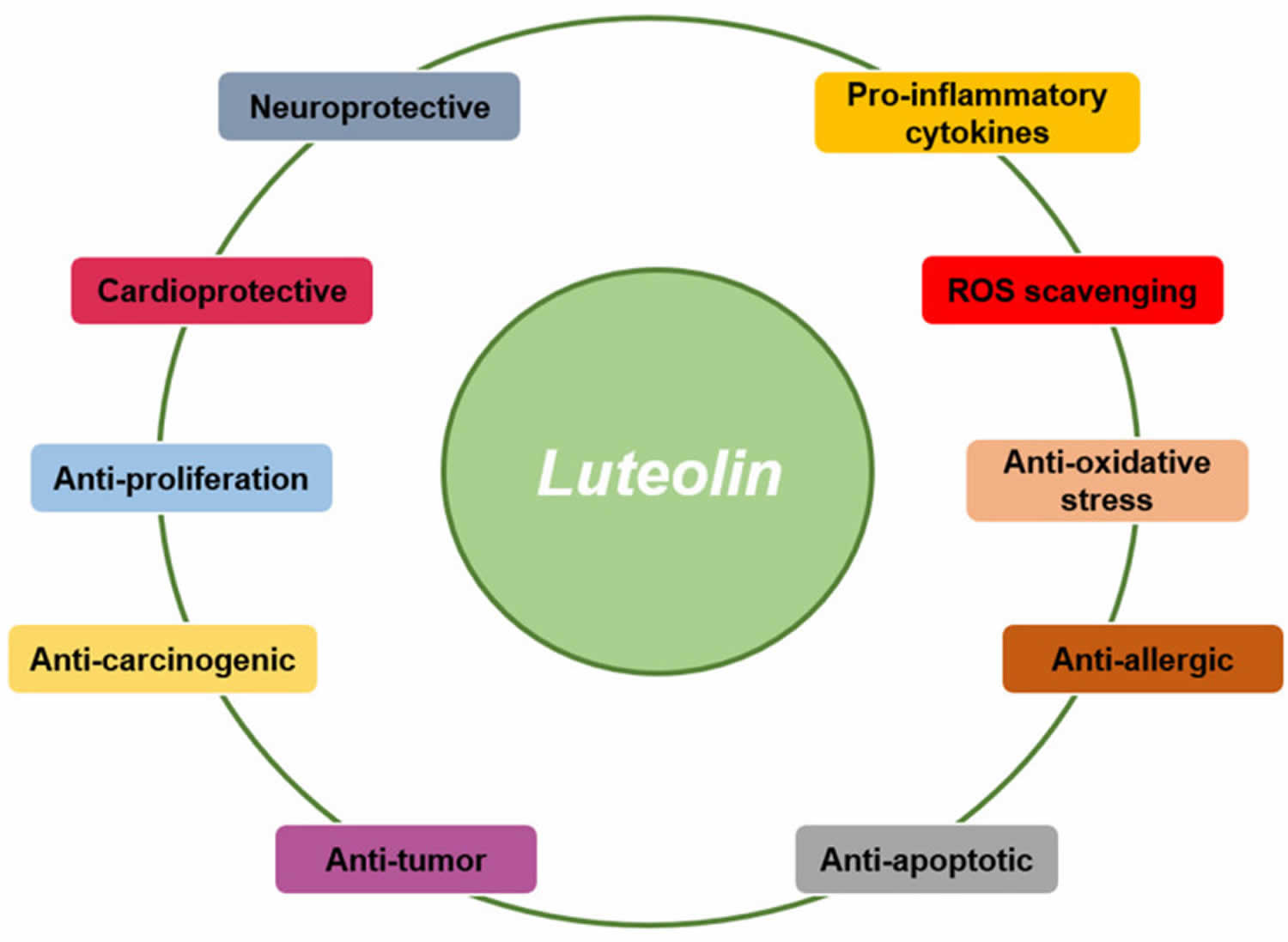
Footnotes: Various kinds of immunopharmacological activities of luteolin such as neuroprotective, cardioprotective, anti-oxidative, anti-allergic, and anti-tumor actions have been reported so far.
[Source 7 ]Figure 4. Luteolin food sources
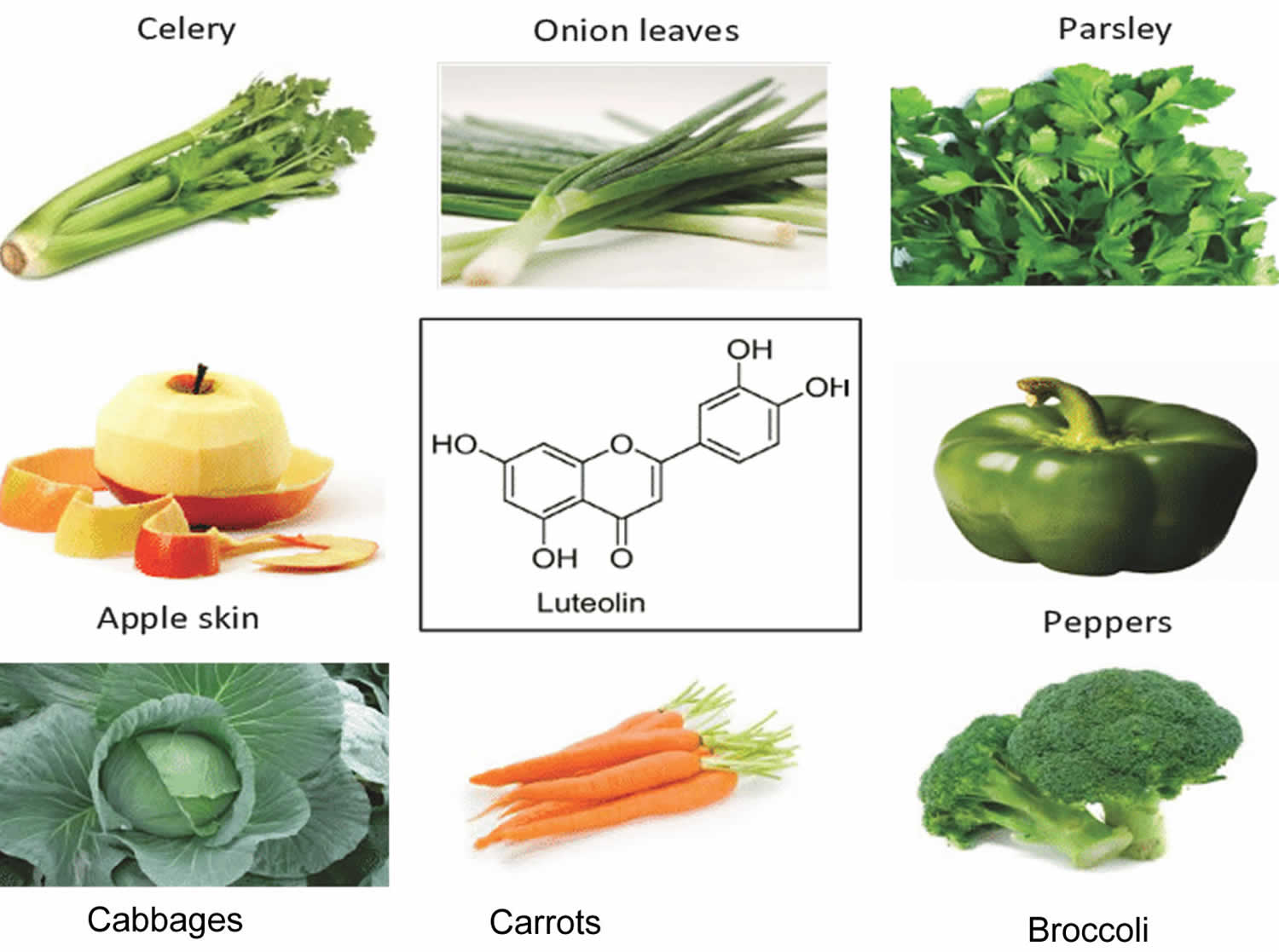
Luteolin food sources
Luteolin is mainly found in various vegetables and fruits, including celery, apple skins, carrots, cabbages, onion leaves, parsley, cabbages, broccoli, peppers and chrysanthemum flowers 27, 28, 29, 30, 5.
Luteolin food sources 5, 28, 31:
- Broccoli (74.5 mg/kg)
- Celery hearts, green (35 mg/kg)
- Celery, raw (10 mg/kg)
- Local celery (80.5 mg/kg)
- Parsley, fresh (11 mg/kg)
- Chili peppers, green, raw (33 mg/kg)
- Green chili (33 mg/kg)
- Bird chili (1035 mg/kg)
- Onion leaves (391 mg/kg)
- Belimbi fruit (202 mg/kg)
- Belimbi leaves (464.5 mg/kg)
- French bean (11 mg/kg)
- Oregano, fresh (10 mg/kg)
- Carrot (37.5 mg/kg)
- White radish (9 mg/kg)
- Peppermint, fresh (127 mg/kg)
- Limau purut leaves (30.5 mg/kg)
- Dried asam gelugur (107.5 mg/kg)
- Thyme, fresh (452 mg/kg).
Luteolin health benefits
Luteolin has shown in several research studies to have anti-inflammatory, antioxidant, anticancer, cytoprotective, anti-microbial, cardioprotective, anti-diabetic, neuroprotective, analgesic and anti-allergic effects 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18. Luteolin, a flavonoid, is mainly found in various vegetables and fruits, including celery, apple skins, carrots, cabbages, onion leaves, parsley, cabbages, broccoli, peppers and chrysanthemum flowers 5.
Anticancer effects
Luteolin has a potential role in cancer management through several mechanisms. Like many other flavonoids, luteolin is able to inhibit the growth of cancer cells derived from nearly all types of cancers, mainly through regulating the cell cycle 32, 33, 34, 35. The anticancer property of luteolin is associated with regulating inflammation, inducing cell-cycle arrest, promoting apoptosis, autophagy, inhibiting angiogenesis, and modulating the PI3/Akt, STAT3 (signal transducer and activator of transcription 3), EGFR (epidermal growth factor receptor) pathways 11, 5. Furthermore, luteolin sensitizes a variety of cancer cells to therapeutically induced cytotoxicity through suppressing cell survival pathways and stimulating apoptosis pathways 11. Notably, luteolin is blood-brain barrier (BBB) permeable, rendering it applicable to the therapy of central nerve system diseases, including brain cancer 36. Furthermore, recent studies have attributed a cancer prevention potential to luteolin 11.
In cancer development, luteolin can reverse epithelial-mesenchymal transition (EMT) by inducing the expression of the epithelial biomarker E-cadherin and downregulating the expression of the mesenchymal biomarkers N-cadherin, snail and vimentin, and also inhibit cell transformation, invasion and metastasis, and angiogenesis during cancer through various pathways such as cell cycle regulation 37. Luteolin can also inhibit cell transformation, invasion and metastasis, and angiogenesis during cancer by regulating the cell cycle, inducing apoptosis, modulating the level of reactive oxygen species (ROS) in tumor cells, and reducing transcription factors 38.
In recent years, a great deal of research has been conducted on the anti-tumor mechanism of luteolin, and luteolin has been shown to induce and enhance apoptosis of cancer cells and inhibit the breakdown of carcinogens that generates active mutagens in liver microsomes, but the specific mechanism of killing cancer cells has not been well explained, and a clear understanding of how luteolin plays a role in affecting the signaling pathways will better promote the role of luteolin in cancer prevention and treatment 39, 40. In earlier studies, it was determined that luteolin potently inhibits human cytochrome P450 (CYP) 1 family enzymes such as CYP1A1, CYP1A2, and CYP1B1, thereby suppressing the mutagenic activation of carcinogens 41. Suppressing the human cytochrome P450 (CYP) 1 enzymes reduces the generation of active mutagens such as benzo[a]pyrene diol epoxide, a metabolite of the tabacco-specific carcinogen benzo[a]pyrene carcinogenesis 42.
Luteolin kills cancer cells by inducing apoptotic cell death in many types of cancer cells, including epidermoid carcinoma, leukemia, pancreatic tumor, and hepatoma 43, 44, 45, 46. Although the mechanisms underlying luteolin-induced apoptosis are complex, they can be generalized as breaking the cell survival and death balance by either enhancing apoptosis or decreasing the survival signaling in cancer cells 11. Luteolin also activates the intrinsic apoptosis pathway through inducing DNA damage and activating p53 47, 48. This is achieved by inhibiting DNA topoisomerases 49, 50. Additionally, luteolin triggers sustained JNK activation that can promote the apoptosis pathway, presumably through modulation of BAD or p53 47, 51, 52. The JNK-mediated p53 activation results in transcriptional expression of Bax that facilitates apoptosis 47, 51, 52. JNK activation causes the mitochondria translocation of Bax and Bak to initiate the intrinsic apoptosis pathway 44, 45.
Luteolin is able to arrest the cell cycle during the G1 phase in human gastric and prostate cancer, and in melanoma cells 53, 54, 55. The G1 cell cycle arrest induced by luteolin is associated with inhibition of the CDK2 activity in melanoma OCM-1 and colorectal cancer HT-29 cells. This arrest is achieved by up-regulation of the CDK inhibitors p27/kip1 and p21/waf1, or direct inhibition on the CDK2 activity 54, 56. Luteolin arrests mouse cancer cell tsFT210 at the G2/M checkpoint 57. DNA damage-activated tumor suppressor protein p53 is involved in both the G1/S and G2/M transition regulation 58, 49. Luteolin can bind and suppress DNA topoisomerases I and II, enzymes essential for repairing damaged DNA, and intercalates directly with the substrate DNA to cause DNA double-strand breaks 49, 59, 50. This action of luteolin induces cell cycle arrest though p53-mediated expression of p21/waf1 54.
The inhibitory effect of luteolin on cancer cell growth is partly achieved through blocking the proliferation signaling pathways induced by these factors. Epidermal growth factor receptor (EGFR) is a typical receptor protein tyrosine kinase (PTK) that mediates cell growth and proliferation. When activated by its ligands, epidermal growth factor receptor (EGFR) is phosphorylated to mediate activation of downstream signaling pathways, including MAPK and PI3K/Akt 60. Luteolin was found to inhibit the proliferation of pancreatic and prostate cancer and human epidermoid cancer cells, which is closely associated with the inhibition of the PTK activity and autophosphorylation of epidermal growth factor receptor (EGFR), transphosphorylation of EGFR downstream effector protein enolase, and activation of MAPK/ERK 46.
Luteolin is able to inhibit IGF-1-induced activation of IGF-1R and Akt, and phosphorylation of the Akt targets p70S6K1, GSK-3β, and FKHR/FKHRL1 11. This inhibition is associated with suppressed expression of cyclin D1, and increased expression of p21/waf1 and proliferation in prostate cancer cells in test tube 61. Luteolin also suppressed prostate tumor growth in animal study through suppressing IGF-1R/Akt signaling 61. Similarly, luteolin inhibits PDGF-induced proliferation by inhibiting PDGF receptor phosphorylation in vascular smooth muscle cells 62. As a consequence, luteolin significantly inhibits PDGF-induced ERK, PI3K/Akt and phospholipase C (PLC)-γ1 activation, and c-fos gene expression. These results suggest that the inhibitory effect of luteolin on the PDGF-induced proliferation may be mediated by blocking phosphorylation of the PDGF receptor 62. As PDGF stimulates cancer cell growth 63, it remains to be determined whether luteolin can block PDGF-induced signaling to suppress cancer cell growth.
In addition to affecting the estrogen receptors (ERs), luteolin may directly target the downstream pathways that are involved in cell growth. For example, protein kinase C, a family of serine-threonine protein kinases that regulates growth factor response and cell proliferation, differentiation and apoptosis 64, 65, can be inhibited in a concentration-dependent manner by luteolin in both cell-free systems and in intact cells 66.
Taken together, the above reports suggest that luteolin suppresses cell growth signaling on distinct components of the growth factor receptor signaling pathways. In addition, carcinogens activate cell survival pathways such as NF-κB and MAPK during the course of cancer formation; these pathways could be additional targets for flavonoids, including luteolin as anti-cancer agents 67, 68.
Figure 5. Luteolin anticancer effects
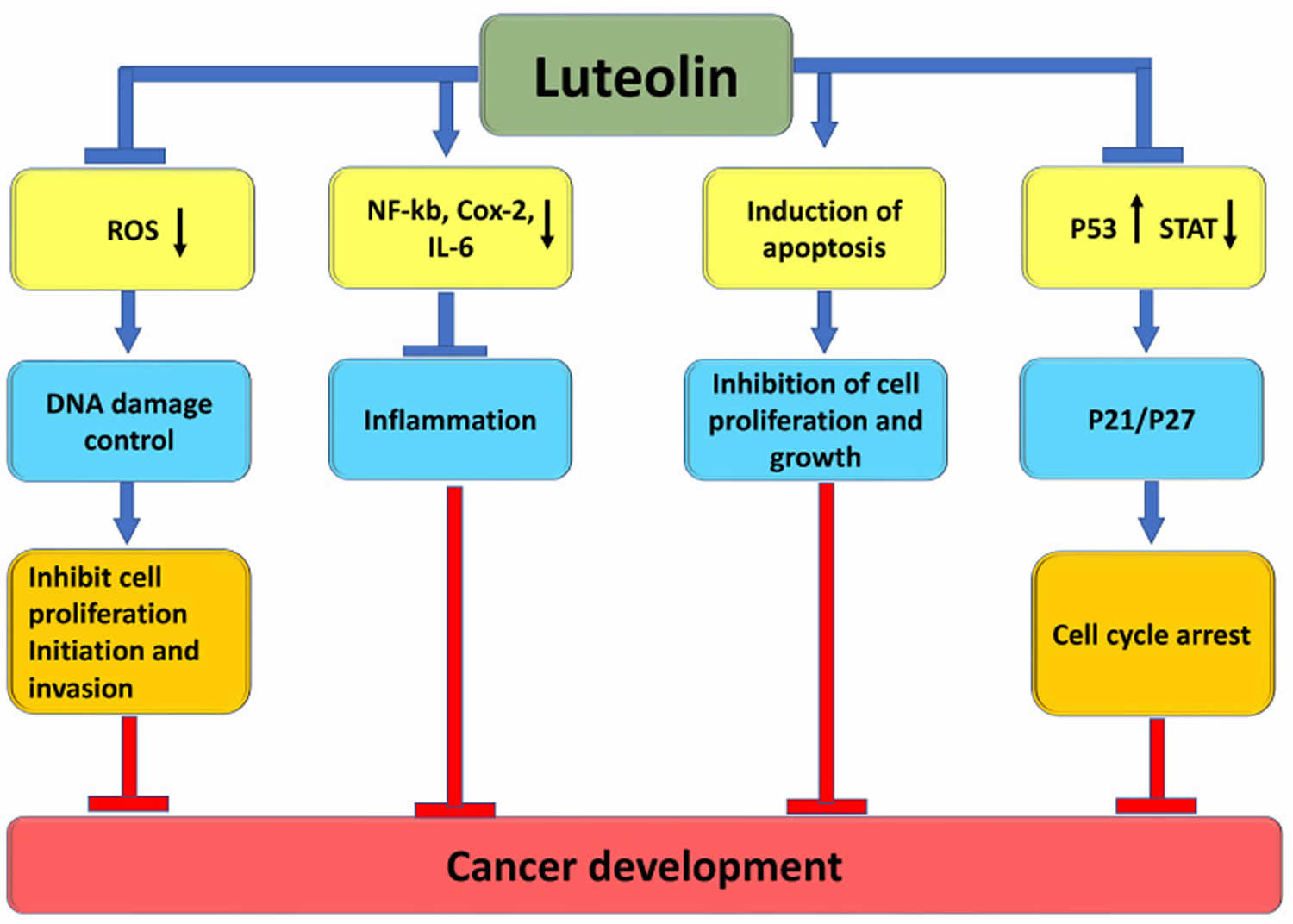
Footnotes: The upward pointing arrow signifies upregulation and the downward pointing arrow signifies downregulation. The block arrow signifies the inhibition.
[Source 5 ]Antioxidant activity
Most flavonoids, including luteolin, are regarded as antioxidants. Reactive oxygen species (ROS) are short-lived unstable molecules that contain oxygen and are highly reactive. Some examples of reactive oxygen species (ROS) include: superoxide (O2•−), hydrogen peroxide (H2O2), hydroxyl radical (•OH), singlet oxygen (1O2), and lipid peroxyl radical (LOO•) 69, 70, 11. Reactive oxygen species (ROS) serve as second messengers for cellular signaling 70. However, excessive production of reactive oxygen species (ROS) results in oxidative stress and damage to DNA, lipids (fatty compounds) and protein that is involved in cancer as well as cardiovascular and neurodegenerative diseases 11. Luteolin was found to inhibit reactive oxygen species (ROS)-induced damage of lipids, DNA, and protein 71, 72. Luteolin has been shown to induce reactive oxygen species (ROS) in untransformed and cancer cells 73, 74. In lung cancer cells, luteolin induced accumulation of superoxide (O2•−) while it reduced hydrogen peroxide (H2O2) concentration 11. However, luteolin could also have pro-oxidant effect depending on the concentration and the source of the free radicals 74, 75. Also, the context and microenvironment of the cell may be important determinants of the outcome of luteolin-induced effects on cellular redox status. For example, the antioxidant activity of luteolin is dependent on copper, vanadium and cadmium ions in the cells 11. Changes in the iron (Fe) ion concentrations dramatically impact the effect of luteolin’s redox-regulating activities. With low iron (Fe) ion concentrations (< 50 μM), luteolin behaves as an antioxidant while high iron (Fe) concentrations (>100 μM) induce luteolin’s pro-oxidative effect 76. Therefore, the anti-and pro-oxidant roles of luteolin in cytotoxicity need to be further investigated 11.
Multiple mechanisms may underlie luteolin’s antioxidant effect. First, luteolin functions as a reactive oxygen species (ROS) scavenger through its own oxidation 77. Luteolin possesses the structures essential to flavonoid’s antioxidant activity: 3′, 4′ hydroxylation, the presence of a double bond between carbons 2 and 3, and a carbonyl group on carbon 4 77. The hydrogen atom from an aromatic hydroxyl group can be donated to free radicals 77. As an aromatic compound, luteolin can support unpaired electrons around the M-electron system 77, 71. Direct evidence showing luteolin as a reactive oxygen species (ROS) scavenger was obtained in cell-free systems 78.
Second, luteolin inhibits reactive oxygen species (ROS)-generating oxidases. For example, luteolin suppresses superoxide (O2•−) formation by inhibiting xanthine oxidase activity 79. However, it is unclear in mammalian cells whether luteolin affects reactive oxygen species (ROS) generation in the mitochondria, the main reactive oxygen species (ROS) generation site, although it interferes with the mitochondrial electron transportation chain in parasite (leishmanial) cells 80.
Third, luteolin may exert its antioxidant effect by protecting or enhancing endogenous antioxidants such as glutathione-S-transferase (GST), glutathione reductase (GR), superoxide dismutase (SOD) and catalase (CAT) 81, 82, 83. Fourth, luteolin may directly inhibit the enzymes that catalyze oxidation of the cellular components. For example, luteolin suppresses lipoxygenase, cyclooxygenase, and ascorbic acid-stimulated malonaldehyde formation in liver lipids 72.
Lastly, luteolin may chelate transition metal ions responsible for the generation of reactive oxygen species (ROS) and therefore inhibit lipooxygenase reaction, or suppress nontransition metal-dependent oxidation 81, 71.
Figure 6. Luteolin antioxidant and pro-oxidant properties
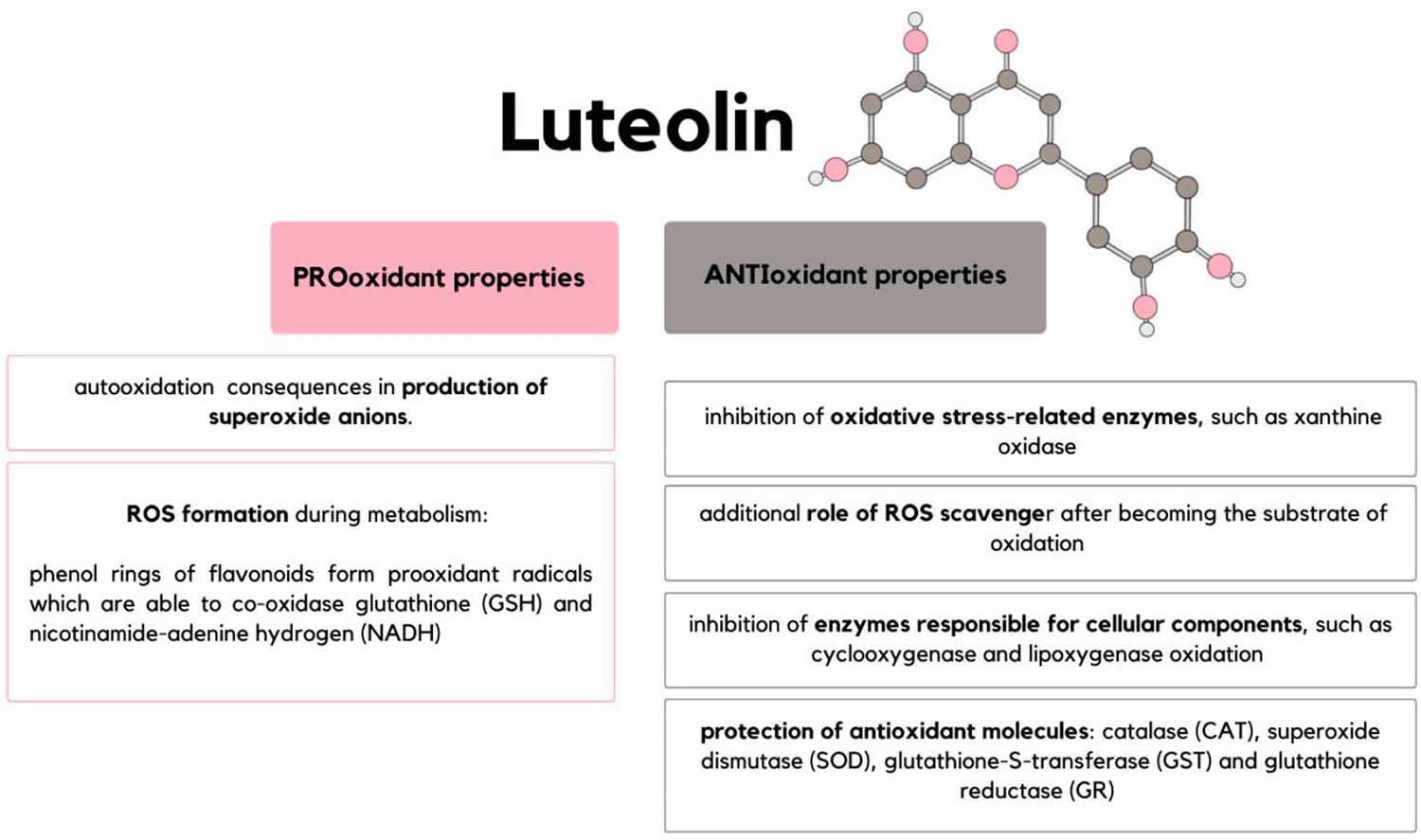
Anti-inflammatory effects
Inflammation is one of the body’s defense mechanisms in response to harmful stimuli, such as damaged cells, pathogens, or irritants. Inflammation guards against infection and helps heal injury. However, chronic (long-term) inflammation may result in harmful diseases such as arthritis, chronic obstructive pulmonary disease, and cancer 84, 85, 86. During inflammation macrophages are activated by various molecules, including cytokines from the host and toxins from the pathogens.
Luteolin is known for its natural anti-inflammatory properties, which can modulate and inhibit the inflammatory response at multiple levels by interfering with several inflammatory pathways and modulators. Luteolin anti-inflammatory effects have also been linked to cancer. Luteolin not only directly affect the production of certain inflammatory signals, but also reduce the release of inflammatory molecules. Luteolin inhibits the synthesis and activity of cyclooxygenase-2 (COX-2), a key enzyme that speeds up the production of prostaglandins. At the same time, luteolin can also inhibit the activation of nuclear factor Kappa-light-chain-enhancer of activated B cells (NF-κB), a key transcription factor in the inflammatory process, thereby reducing the expression of inflammation-related genes 87. The modulation of inflammation-related molecules, pathways and enzymes by luteolin highlights its broad anti-inflammatory capabilities.
Lipopolysaccharide (LPS), an outer membrane component of Gram-negative bacteria, is a common endotoxin and inflammation trigger 11. The activated macrophages produce inflammatory molecules such as tumor necrosis factor alpha (TNFα), interleukins (ILs), and free radicals (reactive oxygen species [ROS] and reactive nitrogen species [RNS]), leading to recruitment of inflammatory cells, such as neutrophils and lymphocytes, to the infection site and clearance of the pathogens 84, 86. Persistent production of these inflammatory molecules during chronic inflammation can result in diseases such as cancer 11. Luteolin exerts its anti-inflammatory effect through suppressing the production of these cytokines and their signal transduction pathways 88, 89, 90. Experiments with animals show that luteolin suppresses lipopolysaccharide (LPS) or bacteria-induced inflammation in animal studies 91, 92. Lipopolysaccharide (LPS)-induced-high mortality was effectively alleviated by luteolin, which is associated with reduction of LPS-stimulated tumor necrosis factor alpha (TNFα) release in serum and intercellular adhesion molecule-1 (ICAM-1) expression in mice liver 91. Luteolin was found to suppress inflammation in mice lung tissue that was caused by Chlamydia pneumoniae 92.
Test tube experiments provided more direct evidence of luteolin’s anti-inflammatory effect. Pretreating mice macrophages (RAW 264.7) with luteolin inhibited LPS-stimulated TNFα and IL-6 release, which was associated with blockage of LPS-induced activation of nuclear factor kappa B (NF-κB) and mitogen-activated protein kinase (MAPK) family members ERK, p38, and JNK 93, 94. Nuclear factor kappa B (NF-κB) and mitogen-activated protein kinase (MAPK) are two major pathways that are involved in macrophage activation and in responses of tissue epithelial and stromal cells to inflammation mediators such as tumor necrosis factor alpha (TNFα) and interleukins (ILs) 95. Suppression of these pathways by luteolin underlies the main mechanism of its inhibitory effect on both acute and chronic inflammation. The suppression of inflammatory cytokine-induced signaling is at least partly on the level of the receptor, because accumulation of lipid rafts, which is the critical step for receptor signaling, was blocked by luteolin 88.
Nuclear factor kappa B (NF-κB) can be activated by both the primary (LPS) and secondary (TNFα and IL-1) inflammatory stimulators 11. Through binding to Toll-like receptor 4 (TLR-4), LPS activates the IκB kinase (IKK), which in turn phosphorylates IκB to trigger its rapid degradation. This allows NF-κB to migrate into the nucleus and activate its targets, including a number of genes with anti-apoptotic properties and cytokines such as tumor necrosis factor alpha (TNFα) and interleukin 1 (IL-1) 96. A positive feedback loop for NF-κB activation is established by these cytokines through binding to their cognate receptors. The NF-κB pathways activated by LPS and the inflammatory cytokines converge at IKK activation 96. Luteolin can effectively block the NF-κB pathway and interfere with the functions of the primary (LPS) and secondary (TNFα and IL-1) inflammatory stimulators through inhibiting IκB kinase (IKK) activation and IκB degradation 93, 90, 89. However, it remains to be determined whether luteolin directly inhibits IκB kinase (IKK) activity or blocks the upstream steps in the IKK activation pathway such as the formation of the receptor signaling complex. On the other hand, the mechanism by which luteolin suppresses MAPK, which is awaiting the dissection of the MAPKKK-MAPKK-MAPK cascade for each MAPK activation, is less well understood. It is unlikely that luteolin suppresses the binding TNFα and IL-1 to their respective receptors because luteolin selectively suppresses each MAPK in macrophages 94.
Based on the observations that some flavonoids with strong antioxidant activities are completely ineffective in suppressing LPS-stimulated TNFα production, it is assumed that the inhibitory action of flavonoids on proinflammatory cytokine production is not directly associated with their antioxidant properties 97. However, because luteolin is able to scavenge reactive oxygen species (ROS) directly and to suppress the LPS-activated nitric oxide production in activated macrophages, the antioxidant activity of luteolin at least in part contributes to luteolin’s anti-inflammatory effect 98, 99. Because inflammation and its involved signaling pathways are strongly associated with formation of a cancer (oncogenesis or tumorigenesis) 100, 101, luteolin’s anti-inflammation role may contribute to cancer prevention 11.
Estrogenic and anti-estrogenic effects
Estrogens are a group of hormones that play an important role in the normal sexual characteristics and reproductive development in women. Estrogens also play a role in other body systems, including the heart, blood vessels, bones, skin, hair, and brain. The woman’s ovaries make most estrogen hormones, although the adrenal glands and fat cells also make small amounts of estrogen.
In response to estrogens, the estrogen receptor (ER) is activated to stimulate DNA synthesis and cell proliferation 102. Flavonoids are naturally occurring phytoestrogens because they can bind to estrogen receptors (ERs) and activate their signaling pathways 103, 32, 104. Because luteolin possesses potent estrogenic activity at low concentrations, it could be a useful agent for hormone replacement therapy 105. However, there are also reports showing anti-estrogenic effects of luteolin 32. The mechanism behind this apparently contradictory estrogenic effect may be attributed to luteolin’s relative low estrogenic activity when it binds to estrogen receptors (ERs) 11. Flavonoids bind and activate estrogen receptors (ERs) when estrogen is deficient. However, due to flavonoids relative weak estrogenic activity, which is 103 to 105 fold lower than 17-β-estradialluteolin, flavonoids may function as anti-estrogenic agents through competition with estrogens for binding to estrogen receptors (ERs) 105, 106. Another mechanism of luteolin’s anti-estrogenic activity is that it inhibits aromatase activity whose function is to aromatize androgens and produce estrogens 107. Additionally, luteolin reduces the estrogen receptor (ER) expression level through inhibiting transcription of the ER gene or potentiating degradation of the ER protein 108, 109. Finally, some alternative signaling mechanisms unrelated to estrogen receptors (ERs) could also be involved 103. Although the interaction of estrogen agonists and antagonists with the estrogen receptor (ER) is a primary event in estrogen action, mammalian cells contain a second binding site (type II [3H] site) for estrogen to control cell growth, which resides in endogenous proteins such as histone 110. Luteolin was found to bind to nuclear type II [3H] sites irreversibly and to compete for estradiol binding to these sites 111.
The cause of breast, prostate, ovarian, and endometrial cancers is associated with estrogen activity. Therefore, consumption of luteolin in diet may reduce risk of these cancers through regulation of estrogen-induced cellular effects 11. Luteolin, as well as other flavonoids, is able to inhibit DNA synthesis and proliferation in mammary epithelial cells and breast cancer cells induced by estrogens, both in test tube and in animal studies 112, 32. Suppressing estrogen-induced cancer cell growth may contribute to luteolin’s therapeutic and preventive activities against estrogen-associated cancer 11.
Luteolin uses
Luteolin has been shown in test tube and animal studies to have a variety of physiological and pharmacological effects, such as antioxidant, anti-aging, analgesic (painkilling), anti-inflammatory, anti-allergy, anti-diabetic, chemotherapeutic, cardioprotective, and neuroprotective properties, attenuation of atherosclerosis and improve of heart ischemia-reperfusion injury 113, 114, 115, 116, 117, 118, 119, 120, 121, 122. In addition, luteolin also has anticancer activities with a large number of studies having shown that luteolin can significantly inhibit the development of cancers such as melanoma, breast cancer, bladder cancer, non-small cell lung cancer, laryngeal cancer, and gastrointestinal malignancies 123, 124, 125, 126, 127, 128, 129, 130. It is worth noting that the anti-inflammatory and antioxidant effects of luteolin may be correlated with their anti-cancer effects. This may be due to the fact that luteolin, as a substance with multiple biological effects, are mostly interrelated in their pharmacological actions and functions 131.
The main reason for the multiple biological activities of luteolin is its antioxidant capacity, which has been shown to have a strong free radical scavenging ability. Luteolin can react with free radicals, providing hydrogen to the free radicals and transforming itself into stable phenoxyl radicals, thus reducing the oxidation of free radicals and preventing them from causing damage to cells 132. It can be seen that the stronger antioxidant property of luteolin can effectively scavenge free radicals during cancer development, prevent cellular DNA damage and mutation, and control the development of cancer.
However, at present time, the studies on the anti-inflammatory, antioxidant and anticancer properties of luteolin in numerous disorders are only in the test tube cells and animal studies, lacking human clinical trials. Therefore, based on test tube experiments and animal model experiments, large-scale randomized controlled trials (RCTs) are needed to confirm the anticancer effect of luteolin and to explore the optimal dosage of luteolin to exert its anticancer effect, in order to provide a more practical basis for the use of luteolin in cancer therapy and to improve the bioavailability and therapeutic effect of luteolin to a greater extent.
Furthermore, due to the poor water solubility and low bioactivity of luteolin, systemic administration of luteolin may not result in the desired therapeutic effect in clinical trials 133, 134, 37. Therefore, the focus of large-scale clinical trials should be on improving the bioavailability of luteolin to maximize its anticancer effect 37. For example, drug delivery systems using materials such as nanolipid particles or hydrogels as carriers or structural modifications of luteolin could be used to alter its limitations 37.
Is Luteolin safe?
Luteolin is considered non-toxic. However, luteolin has not been granted Generally Recognized as Safe (GRAS) status by the U.S. Food and Drug Administration (FDA) 1. Currently, luteolin has been used as a dietary supplement and is also incorporated into cosmetic products, as demonstrated by its nontoxic side effects. Luteolin safety is supported by data showing that the oral median lethal dose (LD50, which is the amount of a substance that kills 50% of a group of test animals) in mice and rats exceeded 2500 and 5000 mg/kg, respectively, roughly translating to 219.8−793.7 mg/kg in humans 20. In animal and test tube cells investigations, luteolin demonstrated a lack of detrimental effects on healthy cells and did not result in noteworthy side effects, even at a relatively high concentration of 30 μM 21, 22. Research intended to examine luteolin’s ability to reverse changes caused by exposure to lead in rats showed no side effects at a dose of 50 mg/kg, oral, daily; therefore, the researchers concluded that luteolin could possibly be administered as a dietary supplement in humans 23. In research designed to examine the effectiveness and tolerability in white children with autism spectrum disorder of a dietary supplement containing two flavonoids (>95% pure), including luteolin (100 mg per 10 kg weight per day), for 26 weeks found transient irritability in subjects as the only side effect of treatment and a significant improvement in adaptive functioning as measured by using the Vineland Adaptive Behavior Scales age-equivalent scores 24.
- Aziz N., Kim M.Y., Cho J.Y. Anti-inflammatory effects of luteolin: A review of in vitro, in vivo, and in silico studies. J. Ethnopharmacol. 2018;225:342–358. doi: 10.1016/j.jep.2018.05.019[↩][↩][↩][↩][↩][↩]
- Chen D., Bi A., Dong X., Jiang Y., Rui B., Liu J., Yin Z., Luo L. Luteolin exhibits anti-inflammatory effects by blocking the activity of heat shock protein 90 in macrophages. Biochem. Biophys. Res. Commun. 2014;443:326–332. doi: 10.1016/j.bbrc.2013.11.122[↩][↩]
- Al-Megrin W.A., Alkhuriji A.F., Yousef A.O.S., Metwally D.M., Habotta O.A., Kassab R.B., Abdel Moneim A.E., El-Khadragy M.F. Antagonistic Efficacy of Luteolin against Lead Acetate Exposure-Associated with Hepatotoxicity is Mediated via Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Activities. Antioxidants. 2019;9:10. doi: 10.3390/antiox9010010[↩][↩]
- Panche A.N., Diwan A.D., Chandra S.R. Flavonoids: An overview. J. Nutr. Sci. 2016;5:e47. doi: 10.1017/jns.2016.41[↩][↩]
- Almatroodi SA, Almatroudi A, Alharbi HOA, Khan AA, Rahmani AH. Effects and Mechanisms of Luteolin, a Plant-Based Flavonoid, in the Prevention of Cancers via Modulation of Inflammation and Cell Signaling Molecules. Molecules. 2024 Feb 29;29(5):1093. doi: 10.3390/molecules29051093[↩][↩][↩][↩][↩][↩][↩][↩][↩]
- Caporali S, De Stefano A, Calabrese C, Giovannelli A, Pieri M, Savini I, Tesauro M, Bernardini S, Minieri M, Terrinoni A. Anti-Inflammatory and Active Biological Properties of the Plant-Derived Bioactive Compounds Luteolin and Luteolin 7-Glucoside. Nutrients. 2022 Mar 9;14(6):1155. doi: 10.3390/nu14061155[↩][↩]
- Huang L, Kim MY, Cho JY. Immunopharmacological Activities of Luteolin in Chronic Diseases. Int J Mol Sci. 2023 Jan 21;24(3):2136. doi: 10.3390/ijms24032136[↩][↩][↩]
- Cárdenas-Castro AP, Rochín-Medina JJ, Ramírez K, Tovar J, Sáyago-Ayerdi SG. In vitro intestinal bioaccessibility and colonic biotransformation of polyphenols from mini bell peppers (Capsicum annuum L.) Plant Foods Hum. Nutr. 2022 doi: 10.1007/s11130-022-00948-5[↩][↩]
- Jeon IH, et al. Anti-inflammatory and antipruritic effects of luteolin from perilla (P. frutescens L.) leaves. Molecules. 2014;19:6941–6951. doi: 10.3390/molecules19066941[↩][↩]
- Jeon IH, Kim HS, Kang HJ, Lee HS, Jeong SI, Kim SJ, Jang SI. Anti-inflammatory and antipruritic effects of luteolin from Perilla (P. frutescens L.) leaves. Molecules. 2014 May 27;19(6):6941-51. doi: 10.3390/molecules19066941[↩][↩]
- Lin Y, Shi R, Wang X, Shen HM. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr Cancer Drug Targets. 2008 Nov;8(7):634-46. doi: 10.2174/156800908786241050[↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩]
- De Stefano A, Caporali S, Di Daniele N, Rovella V, Cardillo C, Schinzari F, Minieri M, Pieri M, Candi E, Bernardini S, Tesauro M, Terrinoni A. Anti-Inflammatory and Proliferative Properties of Luteolin-7-O-Glucoside. Int J Mol Sci. 2021 Jan 28;22(3):1321. doi: 10.3390/ijms22031321[↩][↩]
- Lin LZ, Lu S, Harnly JM. Detection and quantification of glycosylated flavonoid malonates in celery, chinese celery, and celery seed by LC-DAD-ESI/MS. J. Agric. Food Chem. 2007;55:1321–1326. doi: 10.1021/jf0624796[↩][↩]
- Harris GK, Qian Y, Leonard SS, Sbarra DC, Shi X. Luteolin and chrysin differentially inhibit cyclooxygenase-2 expression and scavenge reactive oxygen species but similarly inhibit prostaglandin-E 2 formation in RAW 264.7 cells. J. Nutr. 2006;136:1517–1521. doi: 10.1093/jn/136.6.1517[↩][↩]
- Igile GO, et al. Flavonoids from Vernonia amygdalina and Their Antioxidant Activities. J. Agric. Food Chem. 1994;42:2445–2448. doi: 10.1021/jf00047a015[↩][↩]
- Yan J, et al. Luteolin enhances TNF-related apoptosis-inducing ligand’s anticancer activity in a lung cancer xenograft mouse model. Biochem. Biophys. Res. Commun. 2012;417:842–846. doi: 10.1016/j.bbrc.2011.12.055[↩][↩]
- Liang KL, Yu SJ, Huang WC, Yen HR. Luteolin attenuates allergic nasal inflammation via inhibition of interleukin-4 in an allergic rhinitis mouse model and peripheral blood from human subjects with allergic rhinitis. Front. Pharmacol. 2020 doi: 10.3389/fphar.2020.00291[↩][↩]
- Kempuraj D, et al. Neuroprotective effects of flavone luteolin in neuroinflammation and neurotrauma. BioFactors. 2021;47:190–197. doi: 10.1002/biof.1687[↩][↩]
- Harborne JB, Williams CA. Advances in flavonoid research since 1992. Phytochemistry. 2000;55:481–504. doi: 10.1016/s0031-9422(00)00235-1[↩]
- Rakoczy K, Kaczor J, Sołtyk A, Szymańska N, Stecko J, Sleziak J, Kulbacka J, Baczyńska D. Application of Luteolin in Neoplasms and Nonneoplastic Diseases. Int J Mol Sci. 2023 Nov 6;24(21):15995. doi: 10.3390/ijms242115995[↩][↩][↩]
- Lin Y., Shi R., Wang X., Shen H.-M. Luteolin, a flavonoid with potentials for cancer prevention and therapy. Curr. Cancer Drug Targets. 2008;8:634. doi: 10.2174/156800908786241050[↩][↩]
- Yi C., Li G., Ivanov D.N., Wang Z., Velasco M.X., Hernández G., Kaundal S., Villarreal J., Gupta Y.K., Qiao M., et al. Luteolin inhibits Musashi1 binding to RNA and disrupts cancer phenotypes in glioblastoma cells. RNA Biol. 2018;15:1420–1432. doi: 10.1080/15476286.2018.1539607[↩][↩]
- Al-Megrin W.A., Alkhuriji A.F., Yousef A.O.S., Metwally D.M., Habotta O.A., Kassab R.B., Abdel Moneim A.E., El-Khadragy M.F. Antagonistic Efficacy of Luteolin against Lead Acetate Exposure-Associated with Hepatotoxicity is Mediated via Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Activities. Antioxidants. 2020;9:10. doi: 10.3390/antiox9010010[↩][↩]
- Taliou A., Zintzaras E., Lykouras L., Francis K. An open-label pilot study of a formulation containing the anti-inflammatory flavonoid luteolin and its effects on behavior in children with autism spectrum disorders. Clin. Ther. 2013;35:592–602. doi: 10.1016/j.clinthera.2013.04.006[↩][↩]
- Andersen O., Markham R. Flavonoids, Chemistry, Biochemistry and Applications. Volume 1. CRC Press; Boca Raton, FL, USA: 2005. p. 1256.[↩]
- Pandurangan AK, Esa NM. Luteolin, a bioflavonoid inhibits colorectal cancer through modulation of multiple signaling pathways: a review. Asian Pac J Cancer Prev. 2014;15(14):5501-8. doi: 10.7314/apjcp.2014.15.14.5501[↩]
- Neuhouser M.L. Dietary flavonoids and cancer risk: Evidence from human population studies. Nutr. Cancer. 2004;50:1–7. doi: 10.1207/s15327914nc5001_1[↩]
- Miean K.H., Mohamed S. Flavonoid (myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J. Agric. Food Chem. 2001;49:3106–3112. doi: 10.1021/jf000892m[↩][↩]
- Sun T., Xu Z., Wu C.T., Janes M., Prinyawiwatkul W., No H.K. Antioxidant activities of different colored sweet bell peppers (Capsicum annuum L.) J. Food Sci. 2007;72:S98–S102. doi: 10.1111/j.1750-3841.2006.00245.x[↩]
- Mencherini T., Picerno P., Scesa C., Aquino R. Triterpene, Antioxidant, and Antimicrobial Compounds from Melissa officinalis. J. Nat. Prod. 2007;70:1889–1894. doi: 10.1021/np070351s[↩]
- Flavonoids. https://lpi.oregonstate.edu/mic/dietary-factors/phytochemicals/flavonoids[↩]
- Han DH, Denison MS, Tachibana H, Yamada K. Relationship between estrogen receptor-binding and estrogenic activities of environmental estrogens and suppression by flavonoids. Biosci. Biotechnol. Biochem. 2002;66:1479–1487. doi: 10.1271/bbb.66.1479[↩][↩][↩][↩]
- Ko WG, Kang TH, Lee SJ, Kim YC, Lee BH. Effects of luteolin on the inhibition of proliferation and induction of apoptosis in human myeloid leukaemia cells. Phytother Res. 2002;16:295–298. doi: 10.1002/ptr.871[↩]
- Knowles LM, Zigrossi DA, Tauber RA, Hightower C, Milner JA. Flavonoids suppress androgen-independent human prostate tumor proliferation. Nutr. Cancer. 2000;38:116–120. doi: 10.1207/S15327914NC381_16[↩]
- Fotsis T, Pepper MS, Aktas E, Breit S, Rasku S, Adlercreutz H, Wähälä K, Montesano R, Schweigerer L. Flavonoids, dietary-derived inhibitors of cell proliferation and in vitro angiogenesis. Cancer Res. 1997 Jul 15;57(14):2916-21.[↩]
- Wruck CJ, Claussen M, Fuhrmann G, Romer L, Schulz A, Pufe T, Waetzig V, Peipp M, Herdegen T, Gotz ME. Luteolin protects rat PC12 and C6 cells against MPP+ induced toxicity via an ERK dependent Keap1-Nrf2-ARE pathway. J. Neural Transm. Suppl. 2007;72:57–67. doi: 10.1007/978-3-211-73574-9_9[↩]
- Wang R, Li X, Xu Y, Li Y, Zhang W, Guo R, Song J. Progress, pharmacokinetics and future perspectives of luteolin modulating signaling pathways to exert anticancer effects: A review. Medicine (Baltimore). 2024 Aug 23;103(34):e39398. doi: 10.1097/MD.0000000000039398[↩][↩][↩][↩]
- Birt DF, Hendrich S, Wang W. Dietary agents in cancer prevention: flavonoids and isoflavonoids. Pharmacol Ther. 2001 May-Jun;90(2-3):157-77. doi: 10.1016/s0163-7258(01)00137-1[↩]
- Huang MT, Wood AW, Newmark HL, Sayer JM, Yagi H, Jerina DM, Conney AH. Inhibition of the mutagenicity of bay-region diolepoxides of polycyclic aromatic hydrocarbons by phenolic plant flavonoids. Carcinogenesis. 1983;4:1631–1637. doi: 10.1093/carcin/4.12.1631[↩]
- Buening MK, Chang RL, Huang MT, Fortner JG, Wood AW, Conney AH. Activation and inhibition of benzo(a)pyrene and aflatoxin B1 metabolism in human liver microsomes by naturally occurring flavonoids. Cancer Res. 1981 Jan;41(1):67-72.[↩]
- Kim HJ, Lee SB, Park SK, Kim HM, Park YI, Dong MS. Effects of hydroxyl group numbers on the B-ring of 5,7-dihydroxyflavones on the differential inhibition of human CYP 1A and CYP1B1 enzymes. Arch. Pharm. Res. 2005;28:1114–1121. doi: 10.1007/BF02972971[↩]
- Miller KP, Ramos KS. Impact of cellular metabolism on the biological effects of benzo[a]pyrene and related hydrocarbons. Drug Metab. Rev. 2001;33:1–35. doi: 10.1081/dmr-100000138[↩]
- Huang YT, Hwang JJ, Lee PP, Ke FC, Huang JH, Huang CJ, Kandaswami C, Middleton E, Jr., Lee MT. Effects of luteolin and quercetin, inhibitors of tyrosine kinase, on cell growth and metastasis-associated properties in A431 cells overexpressing epidermal growth factor receptor. Br. J. Pharmacol. 1999;128:999–1010. doi: 10.1038/sj.bjp.0702879[↩]
- Cheng A-C, Huang T-C, Lai C-S, Pan M-H. Induction of apoptosis by luteolin through cleavage of Bcl-2 family in human leukemia HL-60 cells. Eur. J. Pharmacol. 2005;509:1–10. doi: 10.1016/j.ejphar.2004.12.026[↩][↩]
- Lee HJ, Wang CJ, Kuo HC, Chou FP, Jean LF, Tseng TH. Induction apoptosis of luteolin in human hepatoma HepG2 cells involving mitochondria translocation of Bax/Bak and activation of JNK. Toxicol. Appl. Pharmacol. 2005;203:124–131. doi: 10.1016/j.taap.2004.08.004[↩][↩]
- Lee LT, Huang YT, Hwang JJ, Lee PP, Ke FC, Nair MP, Kanadaswam C, Lee MT. Blockade of the epidermal growth factor receptor tyrosine kinase activity by quercetin and luteolin leads to growth inhibition and apoptosis of pancreatic tumor cells. Anticancer Res. 2002 May-Jun;22(3):1615-27.[↩][↩]
- Shi R, Huang Q, Zhu X, Ong YB, Zhao B, Lu J, Ong CN, Shen HM. Luteolin sensitizes the anticancer effect of cisplatin via c-Jun NH2-terminal kinase-mediated p53 phosphorylation and stabilization. Mol. Cancer Ther. 2007;6:1338–1347. doi: 10.1158/1535-7163.MCT-06-0638[↩][↩][↩]
- Plaumann B, Fritsche M, Rimpler H, Brandner G, Hess RD. Flavonoids activate wild-type p53. Oncogene. 1996 Oct 17;13(8):1605-14.[↩]
- Helton ES, Chen X. p53 modulation of the DNA damage response. J. Cell. Biochem. 2007;100:883–896. doi: 10.1002/jcb.21091[↩][↩][↩]
- Chowdhury AR, Sharma S, Mandal S, Goswami A, Mukhopadhyay S, Majumder HK. Luteolin, an emerging anti-cancer flavonoid, poisons eukaryotic DNA topoisomerase I. Biochem. J. 2002;366:653–661. doi: 10.1042/BJ20020098[↩][↩]
- Shi RX, Ong CN, Shen HM. Luteolin sensitizes tumor necrosis factor-alpha-induced apoptosis in human tumor cells. Oncogene. 2004;23:7712–7721. doi: 10.1038/sj.onc.1208046[↩][↩]
- Yu C, Minemoto Y, Zhang J, Liu J, Tang F, Bui TN, Xiang J, Lin A. JNK suppresses apoptosis via phosphorylation of the proapoptotic Bcl-2 family protein BAD. Mol. Cell. 2004;13:329–340. doi: 10.1016/s1097-2765(04)00028-0[↩][↩]
- Kobayashi T, Nakata T, Kuzumaki T. Effect of flavonoids on cell cycle progression in prostate cancer cells. Cancer Lett. 2002;176:17–23. doi: 10.1016/s0304-3835(01)00738-8[↩]
- Casagrande F, Darbon JM. Effects of structurally related flavonoids on cell cycle progression of human melanoma cells: regulation of cyclin-dependent kinases CDK2 and CDK1. Biochem. Pharmacol. 2001;61:1205–1215. doi: 10.1016/s0006-2952(01)00583-4[↩][↩][↩]
- Matsukawa Y, Marui N, Sakai T, Satomi Y, Yoshida M, Matsumoto K, Nishino H, Aoike A. Genistein arrests cell cycle progression at G2-M. Cancer Res. 1993 Mar 15;53(6):1328-31.[↩]
- Lim do Y, Jeong Y, Tyner AL, Park JH. Induction of cell cycle arrest and apoptosis in HT-29 human colon cancer cells by the dietary compound luteolin. Am J Physiol Gastrointest Liver Physiol. 2007;292:G66–75. doi: 10.1152/ajpgi.00248.2006[↩]
- Li WX, Cui CB, Cai B, Wang HY, Yao XS. Flavonoids from Vitex trifolia L. inhibit cell cycle progression at G2/M phase and induce apoptosis in mammalian cancer cells. J. Asian Nat. Prod. Res. 2005;7:615–626. doi: 10.1080/10286020310001625085[↩]
- Caino MC, Oliva JL, Jiang H, Penning TM, Kazanietz MG. Benzo[a]pyrene-7,8-dihydrodiol promotes checkpoint activation and G2/M arrest in human bronchoalveolar carcinoma H358 cells. Mol. Pharmacol. 2007;71:744–750. doi: 10.1124/mol.106.032078[↩]
- Yamashita N, Kawanishi S. Distinct mechanisms of DNA damage in apoptosis induced by quercetin and luteolin. Free Radic. Res. 2000;33:623–633. doi: 10.1080/10715760000301141[↩]
- Zhang L, Lau YK, Xi L, Hong RL, Kim DS, Chen CF, Hortobagyi GN, Chang C, Hung MC. Tyrosine kinase inhibitors, emodin and its derivative repress HER-2/neu-induced cellular transformation and metastasis-associated properties. Oncogene. 1998;16:2855–2863. doi: 10.1038/sj.onc.1201813[↩]
- Fang J, Zhou Q, Shi XL, Jiang BH. Luteolin inhibits insulin-like growth factor 1 receptor signaling in prostate cancer cells. Carcinogenesis. 2007;28:713–723. doi: 10.1093/carcin/bgl189[↩][↩]
- Kim JH, Jin YR, Park BS, Kim TJ, Kim SY, Lim Y, Hong JT, Yoo HS, Yun YP. Luteolin prevents PDGF-BB-induced proliferation of vascular smooth muscle cells by inhibition of PDGF beta-receptor phosphorylation. Biochem. Pharmacol. 2005;69:1715–1721. doi: 10.1016/j.bcp.2005.04.002[↩][↩]
- Matei D, Emerson RE, Lai YC, Baldridge LA, Rao J, Yiannoutsos C, Donner DD. Autocrine activation of PDGFRalpha promotes the progression of ovarian cancer. Oncogene. 2006;25:2060–2069. doi: 10.1038/sj.onc.1209232[↩]
- Lucas M, Sanchez-Margalet V. Protein kinase C involvement in apoptosis. Gen. Pharmacol. 1995;26:881–887. doi: 10.1016/0306-3623(94)00295-x[↩]
- Weinstein IB, Kahn SM, O’Driscoll K, Borner C, Bang D, Jiang W, Blackwood A, Nomoto K. The role of protein kinase C in signal transduction, growth control and lipid metabolism. Adv. Exp. Med. Biol. 1997;400A:313–321. doi: 10.1007/978-1-4615-5325-0_44[↩]
- Ferriola PC, Cody V, Middleton E., Jr. Protein kinase C inhibition by plant flavonoids. Kinetic mechanisms and structure-activity relationships. Biochem. Pharmacol. 1989;38:1617. doi: 10.1016/0006-2952(89)90309-2[↩]
- Stathopoulos GT, Sherrill TP, Cheng DS, Scoggins RM, Han W, Polosukhin VV, Connelly L, Yull FE, Fingleton B, Blackwell TS. Epithelial NF-kappaB activation promotes urethane-induced lung carcinogenesis. Proc. Natl. Acad Sci. U.S.A. 2007;104:18514–18519. doi: 10.1073/pnas.0705316104[↩]
- Lee SH, Lee SJ, Kim JH, Park BJ. Chemical carcinogen, N-methyl-N′-nitro-N-nitrosoguanidine, is a specific activator of oncogenic Ras. Cell Cycle. 2007;6:1257–1264. doi: 10.4161/cc.6.10.4243[↩]
- Reactive oxygen species. https://www.cancer.gov/publications/dictionaries/cancer-terms/def/reactive-oxygen-species[↩]
- Nimnual AS, Taylor LJ, Bar-Sagi D. Redox-dependent downregulation of Rho by Rac. Nat. Cell. Biol. 2003;5:236–241. doi: 10.1038/ncb938[↩][↩]
- Brown JE, Rice-Evans CA. Luteolin-rich artichoke extract protects low density lipoprotein from oxidation in vitro. Free Radic. Res. 1998;29:247–255. doi: 10.1080/10715769800300281[↩][↩][↩]
- Robak J, Shridi F, Wolbís M, Królikowska M. Screening of the influence of flavonoids on lipoxygenase and cyclooxygenase activity, as well as on nonenzymic lipid oxidation. Pol J Pharmacol Pharm. 1988 Sep-Oct;40(5):451-8.[↩][↩]
- Matsuo M, Sasaki N, Saga K, Kaneko T. Cytotoxicity of flavonoids toward cultured normal human cells. Biol. Pharm. Bull. 2005;28:253–259. doi: 10.1248/bpb.28.253[↩]
- Ju W, Wang X, Shi H, Chen W, Belinsky SA, Lin Y. A critical role of luteolin-induced reactive oxygen species in blockage of tumor necrosis factor-activated nuclear factor-kappaB pathway and sensitization of apoptosis in lung cancer cells. Mol. Pharmacol. 2007;71:1381–1388. doi: 10.1124/mol.106.032185[↩][↩]
- Cao G, Sofic E, Prior RL. Antioxidant and Prooxidant Behavior of Flavonoids: Structure-Activity Relationships. Free Radic. Biol. Med. 1997;22:749–760. doi: 10.1016/s0891-5849(96)00351-6[↩]
- Sugihara N, Arakawa T, Ohnishi M, Furuno K. Anti- and pro-oxidative effects of flavonoids on metal-induced lipid hydroperoxide-dependent lipid peroxidation in cultured hepatocytes loaded with alpha-linolenic acid. Free Radic. Biol. Med. 1999;27:1313–1323. doi: 10.1016/s0891-5849(99)00167-7[↩]
- Lien EJ, Ren S, Bui HH, Wang R. Quantitative structure-activity relationship analysis of phenolic antioxidants. Free Radic. Biol. Med. 1999;26:285–294. doi: 10.1016/s0891-5849(98)00190-7[↩][↩][↩][↩]
- Shimoi K, Masuda S, Furugori M, Esaki S, Kinae N. Radioprotective effect of antioxidative flavonoids in gamma-ray irradiated mice. Carcinogenesis. 1994;15:2669–2672. doi: 10.1093/carcin/15.11.2669[↩]
- Nagao A, Seki M, Kobayashi H. Inhibition of xanthine oxidase by flavonoids. Biosci. Biotechnol. Biochem. 1999;63:1787–1790. doi: 10.1271/bbb.63.1787[↩]
- Sen N, Das BB, Ganguly A, Banerjee B, Sen T, Majumder HK. Leishmania donovani: intracellular ATP level regulates apoptosis-like death in luteolin induced dyskinetoplastid cells. Exp. Parasitol. 2006;114:204–214. doi: 10.1016/j.exppara.2006.03.013[↩]
- Ross JA, Kasum CM. Dietary flavonoids: bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002;22:19–34. doi: 10.1146/annurev.nutr.22.111401.144957[↩][↩]
- Leung HW, Kuo CL, Yang WH, Lin CH, Lee HZ. Antioxidant enzymes activity involvement in luteolin-induced human lung squamous carcinoma CH27 cell apoptosis. Eur. J. Pharmacol. 2006;534:12–18. doi: 10.1016/j.ejphar.2006.01.021[↩]
- Manju V, Nalini N. Chemopreventive potential of luteolin during colon carcinogenesis induced by 1,2-dimethylhydrazine. Ital J Biochem. 2005 Sep-Dec;54(3-4):268-75.[↩]
- Brody JS, Spira A. State of the art. Chronic obstructive pulmonary disease, inflammation, and lung cancer. Proc. Am. Thorac. Soc. 2006;3:535–537. doi: 10.1513/pats.200603-089MS[↩][↩]
- Perwez Hussain S, Harris CC. Inflammation and cancer: an ancient link with novel potentials. Int. J. Cancer. 2007;121:2373–2380. doi: 10.1002/ijc.23173[↩]
- Karin M, Lawrence T, Nizet V. Innate immunity gone awry: linking microbial infections to chronic inflammation and cancer. Cell. 2006;124:823–835. doi: 10.1016/j.cell.2006.02.016[↩][↩]
- Wang T, Yin Y, Jiang X, Ruan Y, Xu J, Hu X, Li T, Chu L, Li L. Exploring the mechanism of luteolin by regulating microglia polarization based on network pharmacology and in vitro experiments. Sci Rep. 2023 Aug 23;13(1):13767. doi: 10.1038/s41598-023-41101-9[↩]
- Kumazawa Y, Kawaguchi K, Takimoto H. Immunomodulating effects of flavonoids on acute and chronic inflammatory responses caused by tumor necrosis factor alpha. Curr. Pharm. Des. 2006;12:4271–4279. doi: 10.2174/138161206778743565[↩][↩]
- Xagorari A, Papapetropoulos A, Mauromatis A, Economou M, Fotsis T, Roussos C. Luteolin inhibits an endotoxin-stimulated phosphorylation cascade and proinflammatory cytokine production in macrophages. J Pharmacol Exp Ther. 2001 Jan;296(1):181-7. https://jpet.aspetjournals.org/content/296/1/181.long[↩][↩]
- Chen CC, Chow MP, Huang WC, Lin YC, Chang YJ. Flavonoids inhibit tumor necrosis factor-alpha-induced up-regulation of intercellular adhesion molecule-1 (ICAM-1) in respiratory epithelial cells through activator protein-1 and nuclear factor-kappaB: structure-activity relationships. Mol Pharmacol. 2004 Sep;66(3):683-93. https://molpharm.aspetjournals.org/content/66/3/683.long[↩][↩]
- Kotanidou A, Xagorari A, Bagli E, Kitsanta P, Fotsis T, Papapetropoulos A, Roussos C. Luteolin Reduces Lipopolysaccharide-induced Lethal Toxicity and Expression of Proinflammatory Molecules in Mice. Am. J. Respir. Crit. Care Med. 2002;165:818–823. doi: 10.1164/ajrccm.165.6.2101049[↩][↩]
- Tormakangas L, Vuorela P, Saario E, Leinonen M, Saikku P, Vuorela H. In vivo treatment of acute Chlamydia pneumoniae infection with the flavonoids quercetin and luteolin and an alkyl gallate, octyl gallate, in a mouse model. Biochem. Pharmacol. 2005;70:1222–1230. doi: 10.1016/j.bcp.2005.07.012[↩][↩]
- Chen CY, Peng WH, Tsai KD, Hsu SL. Luteolin suppresses inflammation-associated gene expression by blocking NF-kappaB and AP-1 activation pathway in mouse alveolar macrophages. Life Sci. 2007;81:1602–1614. doi: 10.1016/j.lfs.2007.09.028[↩][↩]
- Xagorari A, Roussos C, Papapetropoulos A. Inhibition of LPS-stimulated pathways in macrophages by the flavonoid luteolin. Br. J. Pharmacol. 2002;136:1058–1064. doi: 10.1038/sj.bjp.0704803[↩][↩]
- Karin M. Mitogen activated protein kinases as targets for development of novel anti-inflammatory drugs. Ann. Rheum. Dis. 2004;63(Suppl 2):ii62–ii64. doi: 10.1136/ard.2004.028274[↩]
- Hayden MS, Ghosh S. Signaling to NF-kappaB. Genes Dev. 2004;18:2195–2224. doi: 10.1101/gad.1228704[↩][↩]
- Devasagayam TP, Subramanian M, Singh BB, Ramanathan R, Das NP. Protection of plasmid pBR322 DNA by flavonoids against single-stranded breaks induced by singlet molecular oxygen. J. Photochem. Photobiol. B. 1995;30:97–103. doi: 10.1016/1011-1344(95)07159-y[↩]
- Kim HK, Cheon BS, Kim YH, Kim SY, Kim HP. Effects of naturally occurring flavonoids on nitric oxide production in the macrophage cell line RAW 264.7 and their structure-activity relationships. Biochem. Pharmacol. 1999;58:759–765. doi: 10.1016/s0006-2952(99)00160-4[↩]
- Hu C, Kitts DD. Luteolin and luteolin-7-O-glucoside from dandelion flower suppress iNOS and COX-2 in RAW264.7 cells. Mol. Cell. Biochem. 2004;265:107–113. doi: 10.1023/b:mcbi.0000044364.73144.fe[↩]
- Kamata H, Honda S, Maeda S, Chang L, Hirata H, Karin M. Reactive oxygen species promote TNFalpha-induced death and sustained JNK activation by inhibiting MAP kinase phosphatases. Cell. 2005;120:649–661. doi: 10.1016/j.cell.2004.12.041[↩]
- Karin M, Greten FR. NF-kappaB: linking inflammation and immunity to cancer development and progression. Nat. Rev. Immunol. 2005;5:749–759. doi: 10.1038/nri1703[↩]
- Colditz GA. Estrogen, estrogen plus progestin therapy, and risk of breast cancer. Clin Cancer Res. 2005 Jan 15;11(2 Pt 2):909s-17s.[↩]
- Collins-Burow BM, Burow ME, Duong BN, McLachlan JA. Estrogenic and antiestrogenic activities of flavonoid phytochemicals through estrogen receptor binding-dependent and -independent mechanisms. Nutr. Cancer. 2000;38:229–244. doi: 10.1207/S15327914NC382_13[↩][↩]
- Ise R, Han D, Takahashi Y, Terasaka S, Inoue A, Tanji M, Kiyama R. Expression profiling of the estrogen responsive genes in response to phytoestrogens using a customized DNA microarray. FEBS Lett. 2005;579:1732–1740. doi: 10.1016/j.febslet.2005.02.033[↩]
- Zand RS, Jenkins DJ, Diamandis EP. Steroid hormone activity of flavonoids and related compounds. Breast Cancer Res. Treat. 2000;62:35–49. doi: 10.1023/a:1006422302173[↩][↩]
- Murkies AL, Wilcox G, Davis SR. Phytoestrogens. J. Clin. Endocrinol. Metab. 1998;83:297–303. doi: 10.1210/jcem.83.2.4577[↩]
- Wang C, Makela T, Hase T, Adlercreutz H, Kurzer MS. Lignans and flavonoids inhibit aromatase enzyme in human preadipocytes. J. Steroid. Biochem. Mol. Biol. 1994;50:205–212. doi: 10.1016/0960-0760(94)90030-2[↩]
- Wang TT, Milner MJ, Milner JA, Kim YS. Estrogen receptor alpha as a target for indole-3-carbinol. J. Nutr. Biochem. 2006;17:659–664. doi: 10.1016/j.jnutbio.2005.10.012[↩]
- Chiu FL, Lin JK. Downregulation of androgen receptor expression by luteolin causes inhibition of cell proliferation and induction of apoptosis in human prostate cancer cells and xenografts. Prostate. 2008;68:61–71. doi: 10.1002/pros.20690[↩]
- Shoulars K, Rodriguez MA, Crowley J, Turk J, Thompson T, Markaverich BM. Reconstitution of the type II [3H]estradiol binding site with recombinant histone H4. J. Steroid. Biochem. Mol. Biol. 2006;99:1–8. doi: 10.1016/j.jsbmb.2005.11.009[↩]
- Markaverich BM, Roberts RR, Alejandro MA, Johnson GA, Middleditch BS, Clark JH. Bioflavonoid interaction with rat uterine type II binding sites and cell growth inhibition. J. Steroid. Biochem. 1988;30:71–78. doi: 10.1016/0022-4731(88)90078-7[↩]
- Holland MB, Roy D. Estrone-induced cell proliferation and differentiation in the mammary gland of the female Noble rat. Carcinogenesis. 1995;16:1955–1961. doi: 10.1093/carcin/16.8.1955[↩]
- Lopez-Lazaro M. Distribution and Biological Activities of the Flavonoid Luteolin. Mini-Rev. Med. Chem. 2008;9:31–59. doi: 10.2174/138955709787001712[↩]
- Xiao X, Wang X, Gui X, Chen L, Huang B. Natural flavonoids as promising analgesic candidates: a systematic review. Chem Biodivers. (2016) 13(11):1427–40. 10.1002/cbdv.201600060[↩]
- Yu Y, Wang Y, Yue X, et al. Determination of luteolin and antioxidant capacity of luteolin in Tiger’s Eye Wannabe by RP-HPLC. J Changchun Normal Univ. 2021;40:70–5.[↩]
- Gendrisch F, Esser PR, Schempp CM, Wölfle U. Luteolin as a modulator of skin aging and inflammation. Biofactors. 2021 Mar;47(2):170-180. doi: 10.1002/biof.1699[↩]
- Dar AA, Fehaid A, Alkhatani S, Alarifi S, Alqahtani WS, Albasher G, Almeer R, Alfarraj S, Moneim AA. The protective role of luteolin against the methotrexate-induced hepato-renal toxicity via its antioxidative, anti-inflammatory, and anti-apoptotic effects in rats. Hum Exp Toxicol. 2021 Jul;40(7):1194-1207. doi: 10.1177/0960327121991905[↩]
- Conti P, Caraffa A, Gallenga CE, Ross R, Kritas SK, Frydas I, Younes A, Di Emidio P, Ronconi G, Pandolfi F. Powerful anti-inflammatory action of luteolin: Potential increase with IL-38. Biofactors. 2021 Mar;47(2):165-169. doi: 10.1002/biof.1718[↩]
- Sun WL, Yang JW, Dou HY, Li GQ, Li XY, Shen L, Ji HF. Anti-inflammatory effect of luteolin is related to the changes in the gut microbiota and contributes to preventing the progression from simple steatosis to nonalcoholic steatohepatitis. Bioorg Chem. 2021 Jul;112:104966. doi: 10.1016/j.bioorg.2021.104966[↩]
- Ding X, Zheng L, Yang B, Wang X, Ying Y. Luteolin Attenuates Atherosclerosis Via Modulating Signal Transducer And Activator Of Transcription 3-Mediated Inflammatory Response. Drug Des Devel Ther. 2019 Nov 18;13:3899-3911. doi: 10.2147/DDDT.S207185[↩]
- Li J, Dong JZ, Ren YL, Zhu JJ, Cao JN, Zhang J, Pan LL. Luteolin decreases atherosclerosis in LDL receptor-deficient mice via a mechanism including decreasing AMPK-SIRT1 signaling in macrophages. Exp Ther Med. 2018 Sep;16(3):2593-2599. doi: 10.3892/etm.2018.6499[↩]
- Wei B, Lin Q, Ji YG, Zhao YC, Ding LN, Zhou WJ, Zhang LH, Gao CY, Zhao W. Luteolin ameliorates rat myocardial ischaemia-reperfusion injury through activation of peroxiredoxin II. Br J Pharmacol. 2018 Aug;175(16):3315-3332. doi: 10.1111/bph.14367[↩]
- Wei J, Zeng J. Effects of luteolin on proliferation and migration of melanoma A375 cells and its mechanism of action. J Guangxi Med Univ. 2019;36:1894–7.[↩]
- Lan H. Functional Study on the Inhibition of Breast Cancer Cell Growth by Luteolin. Zhejiang University; 2020.[↩]
- Zhang X, Li L, Shi X, et al. Effect of luteolin on apoptosis of laryngeal cancer Hep2 cells. J Zhengzhou Univ (Med Ed). 2019;54:883–6.[↩]
- Iida K, Naiki T, Naiki-Ito A, Suzuki S, Kato H, Nozaki S, Nagai T, Etani T, Nagayasu Y, Ando R, Kawai N, Yasui T, Takahashi S. Luteolin suppresses bladder cancer growth via regulation of mechanistic target of rapamycin pathway. Cancer Sci. 2020 Apr;111(4):1165-1179. doi: 10.1111/cas.14334[↩]
- Masraksa W, Tanasawet S, Hutamekalin P, Wongtawatchai T, Sukketsiri W. Luteolin attenuates migration and invasion of lung cancer cells via suppressing focal adhesion kinase and non-receptor tyrosine kinase signaling pathway. Nutr Res Pract. 2020 Apr;14(2):127-133. doi: 10.4162/nrp.2020.14.2.127[↩]
- Sagawa H, Naiki-Ito A, Kato H, Naiki T, Yamashita Y, Suzuki S, Sato S, Shiomi K, Kato A, Kuno T, Matsuo Y, Kimura M, Takeyama H, Takahashi S. Connexin 32 and luteolin play protective roles in non-alcoholic steatohepatitis development and its related hepatocarcinogenesis in rats. Carcinogenesis. 2015 Dec;36(12):1539-49. doi: 10.1093/carcin/bgv143[↩]
- Pu Y, Zhang T, Wang J, Mao Z, Duan B, Long Y, Xue F, Liu D, Liu S, Gao Z. Luteolin exerts an anticancer effect on gastric cancer cells through multiple signaling pathways and regulating miRNAs. J Cancer. 2018 Sep 8;9(20):3669-3675. doi: 10.7150/jca.27183[↩]
- Yao Y, Rao C, Zheng G, Wang S. Luteolin suppresses colorectal cancer cell metastasis via regulation of the miR‑384/pleiotrophin axis. Oncol Rep. 2019 Jul;42(1):131-141. doi: 10.3892/or.2019.7136[↩]
- Franza L, Carusi V, Nucera E, Pandolfi F. Luteolin, inflammation and cancer: Special emphasis on gut microbiota. Biofactors. 2021 Mar;47(2):181-189. doi: 10.1002/biof.1710[↩]
- Chen F, Su X, Gao J, Liu Y, Zhang Q, Luo D. A modified strategy to improve the dissolution of flavonoids from Artemisiae Argyi Folium using ultrasonic-assisted enzyme-deep eutectic solvent system. J Chromatogr A. 2023 Sep 27;1707:464282. doi: 10.1016/j.chroma.2023.464282[↩]
- Zhu P., Chen L., Zhao Y., Gao C., Yang J., Liao X., Liu D., Yang B. A Novel Host-Guest Complex Based on Biotin Functionalized Polyamine-β-Cyclodextrin for Tumor Targeted Delivery of Luteolin. J. Mol. Struct. 2021;1237:130339. doi: 10.1016/j.molstruc.2021.130339[↩]
- Wang Y., Wang Q., Feng W., Yuan Q., Qi X., Chen S., Yao P., Dai Q., Xia P., Zhang D. Folic Acid-Modified ROS-Responsive Nanoparticles Encapsulating Luteolin for Targeted Breast Cancer Treatment. Drug Deliv. 2021;28:1695–1708. doi: 10.1080/10717544.2021.1963351[↩]



{kind=link}