
Contents
- African trypanosomiasis
African trypanosomiasis
African trypanosomiasis also known as African sleeping sickness or human African trypanosomiasis, is an infectious disease is caused by the parasites Trypanosoma brucei gambiense or Trypanosoma brucei rhodesiense 1. Without treatment, African sleeping sickness is considered fatal 2. African trypanosomiasis is transmitted by the tsetse fly (Glossina species), which is found only in sub-Saharan Africa and is endemic in 36 sub-Saharan African countries where the tsetse flies that transmit the disease live in rural environments 3. Tsetse flies are found in woodland and savannah areas and they bite during daylight hours. Travelers to urban areas are not at risk. The persons most likely to be exposed to the infection are tourists, hunters, and others working in or visiting game parks. Villagers with infected cattle herds are also at risk. African Trypanosomiasis is different from American Trypanosomiasis or Chagas disease that occurs mainly in Latin America, which is caused by Trypanosoma cruzi and transmitted by the Reduuvid insect vector 4.
There are two types of Human African trypanosomiasis (African sleeping sickness); each named for the region of Africa in which it was found historically and depending on the subspecies of the parasite involved 5:
- Trypanosoma brucei gambiense the parasite that cause West African trypanosomiasis or Gambian sleeping sickness is found in 24 countries in west and central Africa and currently accounts for 95% of reported cases of African sleeping sickness, causes a chronic infection and is endemic across Central and West Africa 6. A person gets West African trypanosomiasis through the bite of an infected tsetse fly. Occasionally a pregnant woman may pass the infection to her baby. In theory, the infection can be transmitted through a blood transfusion, but such cases rarely have been documented. A person can be infected for months or even years without major signs or symptoms of the disease. When more evident symptoms emerge, the patient is often already in an advanced disease stage where the central nervous system (brain and spinal cord) is affected. In recent years, 7,000-10,000 new cases of West African trypanosomiasis have been reported to the World Health Organization (WHO) annually. However, many cases are not recognized or reported and the true number of annual cases is likely to be higher. Cases of West African trypanosomiasis imported into the United States are extremely rare.
- Trypanosoma brucei rhodesiense the parasite that cause East African trypanosomiasis, which is also carried by the tsetse fly, is found in 13 countries in eastern and southern Africa. Nowadays, East African trypanosomiasis represents under 5% of reported African sleeping sickness cases and causes an acute infection. Each year, a few hundred cases of East African trypanosomiasis are reported to the World Health Organization (WHO). However, many cases are not recognized or reported and the true number of new cases is higher. Since 1967, 40 cases of East African trypanosomiasis have been diagnosed or treated in the United States, all among individuals who had traveled to eastern Africa. First signs and symptoms are observed a few months or weeks after infection. The disease develops rapidly and invades the central nervous system. Only Uganda presents both forms of the disease, but in separate zones.
African trypanosomiasis is seen in up to 30 countries across sub-Saharan Africa, and over 7000 cases of the disease were reported in 2012 7. The parasite and subsequent disease are classically broken up into west and east African variants. The west/central African form is caused by Trypanosoma brucei gambiense. It is often chronic and deadly if untreated. The east/southern African infection caused by Trypanosoma brucei rhodesiense and in addition to humans is often found in cattle 1. The vast majority of reported infections are caused by Trypanosoma brucei gambiense. Geographical distribution has demonstrated progressive overlap as Trypanosoma brucei rhodesiense has moved northwest 8.
African trypanosomiasis is mainly seen in rural communities and impoverished areas. This distribution is underreported, and although the World Health Organization (WHO) has attempted to re-institute control programs, not all countries report disease or implement these measures.
The chronic form of African trypanosomiasis from Trypanosoma brucei gambiense is rare in short-term tourists and visitors but is seen in refugees and immigrants. In contrast Trypanosoma brucei rhodesiense has been seen in tourists to East Africa, mainly in Tanzania 8.
Longitudinally, the WHO’s goal is to eliminate African trypanosomiasis by 2020. Multiple screening methods, therapeutic delivery plans, and disease reporting programs are being implemented in an attempt to make this a reality.
Figure 1. Human African trypanosomiasis (Trypanosoma brucei parasite)
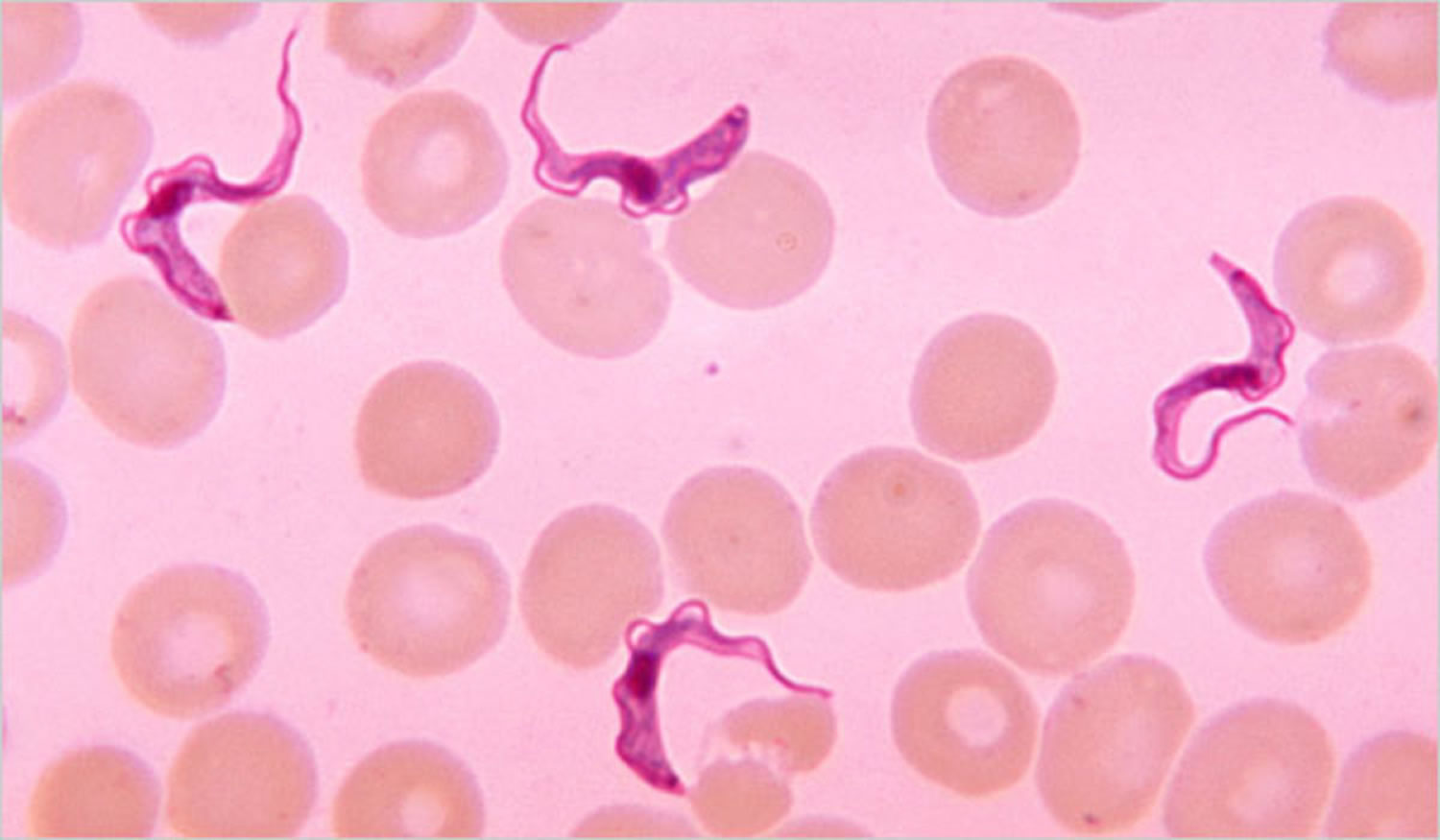
Footnote: Trypanosoma brucei parasites found in a blood smear
East african sleeping sickness
East African trypanosomiasis is caused by the parasite Trypanosoma brucei rhodesiense, which is carried by the tsetse fly. Each year, a few hundred cases of East African trypanosomiasis are reported to the World Health Organization (WHO). However, many cases are not recognized or reported and the true number of new cases is higher. Since 1967, 40 cases of East African trypanosomiasis have been diagnosed or treated in the United States, all among individuals who had traveled to eastern Africa.
A person will get East African trypanosomiasis if he or she is bitten by a tsetse fly infected with the Trypanosoma brucei rhodesiense parasite. The proportion of tsetse flies that are infected with this parasite is low. The tsetse fly is found only in rural Africa. East African trypanosomiasis is found in parts of Eastern and Southeastern Africa. More than 95% of cases are reported from Uganda, Tanzania, Malawi, and Zambia.
East African trypanosomiasis affected individuals typically present after inoculation with a painful eschar and a rapidly progressing illness marked by fevers, rash, fatigue, and myalgias. Within a few weeks to months, the disease progresses to the second stage, with symptoms identical to that of African sleeping sickness Trypanosoma brucei gambiense but with a much-accelerated course that quickly leads to death.
West african sleeping sickness
West African trypanosomiasis also called Gambian sleeping sickness, is caused by a parasite called Trypanosoma brucei gambiense carried by the tsetse fly. In recent years, 7,000-10,000 new cases of West African trypanosomiasis have been reported to the World Health Organization (WHO) annually. However, many cases are not recognized or reported and the true number of annual cases is likely to be higher. Cases of West African trypanosomiasis imported into the United States are extremely rare.
West African trypanosomiasis can be contracted in parts of central Africa and in a few areas of West Africa. Most of the reported cases are found in central Africa (Democratic Republic of Congo, Angola, Sudan, Central African Republic, Republic of Congo, Chad, and northern Uganda). Although the infection is not found in the United States, historically, it has been a serious public health problem in some regions of sub-Saharan Africa. Currently, about 10,000 new cases each year are reported to the World Health organization; however, it is believed that many cases go undiagnosed and unreported. African Trypanosomiasis is considered a neglected tropical disease and is curable with medication, but is fatal if not treated 9.
A person gets West African trypanosomiasis through the bite of an infected tsetse fly. Occasionally a pregnant woman may pass the infection to her baby. In theory, the infection can be transmitted through a blood transfusion, but such cases rarely have been documented.
West African trypanosomiasis initial stage often presents with a painless eschar at the point of infection, though this is often not recalled due to a prolonged asymptomatic course. Patients often have an indolent first stage marked by posterior cervical lymphadenopathy (Winterbottom’s sign), headaches, malaise, and arthralgias. As the disease progresses to the second stage, patients will develop somnolence, fatigue, neurological deficits, tremors, ataxia, seizures, comas, and eventually death.
Who is at risk for contracting West African trypanosomiasis?
The tsetse flies that transmit West African trypanosomiasis are found only in rural areas. Travelers to urban areas are not at risk. The flies bite during daylight hours. They inhabit forests and areas of thick vegetation along rivers and waterholes. Even in areas where the disease is present, most flies are not infected with this parasite, so the risk of infection increases with the number of times a person is bitten by the tsetse fly. Therefore, tourists are not at great risk for contracting West African trypanosomiasis unless they are traveling and spending long periods of time in rural areas of central Africa where the disease is present.
How can I prevent African trypanosomiasis and other insect bites?
- Wear protective clothing, including long-sleeved shirts and pants. The tsetse fly can bite through thin fabrics, so clothing should be made of medium-weight material.
- Wear neutral-colored clothing. The tsetse fly is attracted to bright colors and very dark colors.
- Inspect vehicles for tsetse flies before entering. The tsetse fly is attracted to moving vehicles.
- Avoid bushes. The tsetse fly is less active during the hottest period of the day. It rests in bushes but will bite if disturbed.
- Use insect repellant. Though insect repellants have not proven effective in preventing tsetse fly bites, they are effective in preventing other insects from biting and causing illness.
African sleeping sickness statistics
African trypanosomiasis can occur at any age and can affect all races 10. Human African sleeping sickness threatens millions of people in 36 countries in sub-Saharan Africa 5. Many of the affected populations live in remote rural areas with limited access to adequate health services, which complicates the surveillance and therefore the diagnosis and treatment of cases. In addition, displacement of populations, war and poverty are important factors that facilitate transmission 5.
- In 1998, almost 40 000 cases were reported, but estimates were that 300 000 cases were undiagnosed and therefore untreated.
- During the last epidemic the prevalence reached 50% in several villages in Angola, the Democratic Republic of the Congo, and South Sudan. Sleeping sickness was the first or second greatest cause of mortality in those communities, even ahead of HIV/AIDS.
- In 2009, after continued control efforts, the number of cases reported dropped below 10,000 (9,878) for the first time in 50 years. This decline in number of cases has continued with 992 new cases reported in 2019, the lowest level since the start of systematic global data-collection 80 years ago. The estimated population at risk is 65 million people.
- In the last 10 years, over 70% of reported African sleeping sickness cases occurred in the Democratic Republic of the Congo.
- Angola, Cameroon, Central African Republic, Chad, Congo, Guinea, Malawi, South Sudan and Zambia declared between 10 and 100 new cases in 2019, while Côte d’Ivoire, Equatorial Guinea, Gabon, Uganda, United Republic of Tanzania and Zimbabwe declared between 1 and 10 new cases.
- Countries such as Burkina Faso, Ghana, Kenya and Nigeria, have reported sporadic cases in the last 10 years.
- Countries like Benin, Botswana, Burundi, Ethiopia, Gambia, Guinea Bissau, Liberia, Mali, Mozambique, Namibia, Niger, Rwanda, Senegal, Sierra Leone, Swaziland and Togo have not reported any new cases for over a decade. Transmission of African trypanosomiasis seems to have stopped in some of these countries but there are still some areas where it is difficult to assess the exact situation because the unstable social circumstances and/or difficult accessibility hinder surveillance and diagnostic activities.
Table 1. Cases of human African trypanosomiasis recorded in non-endemic countries from 2011 to 2018
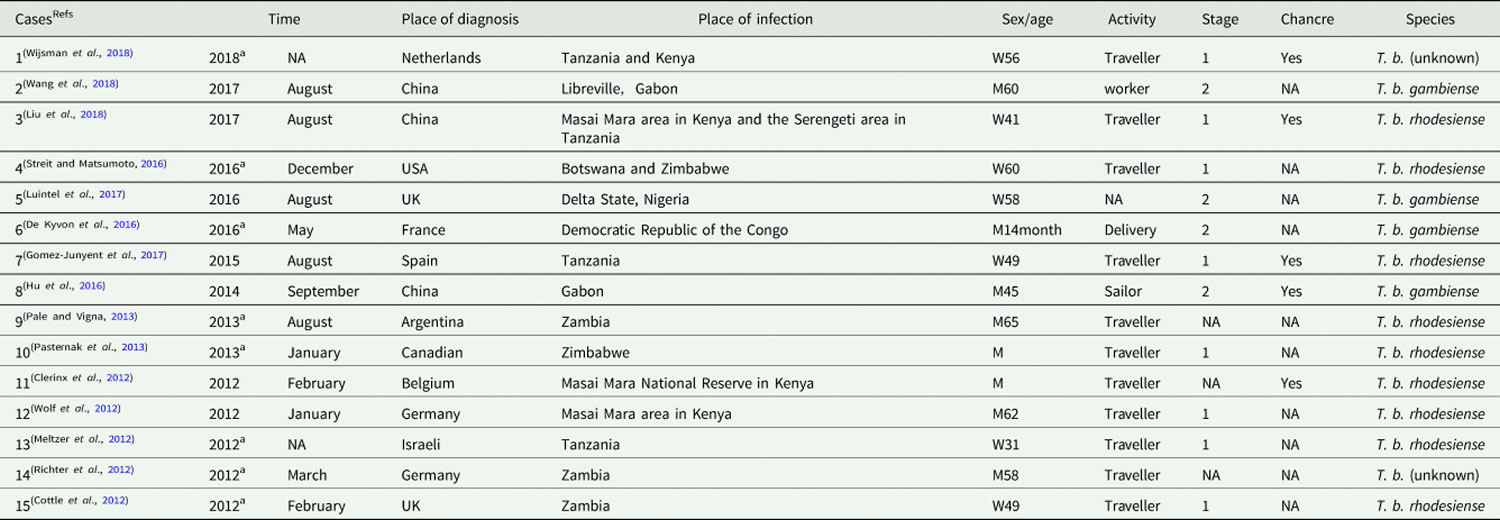
Footnotes: a Time refers to the published date when the diagnostic date is unavailable.
NA = Information is unavailable on the case report.
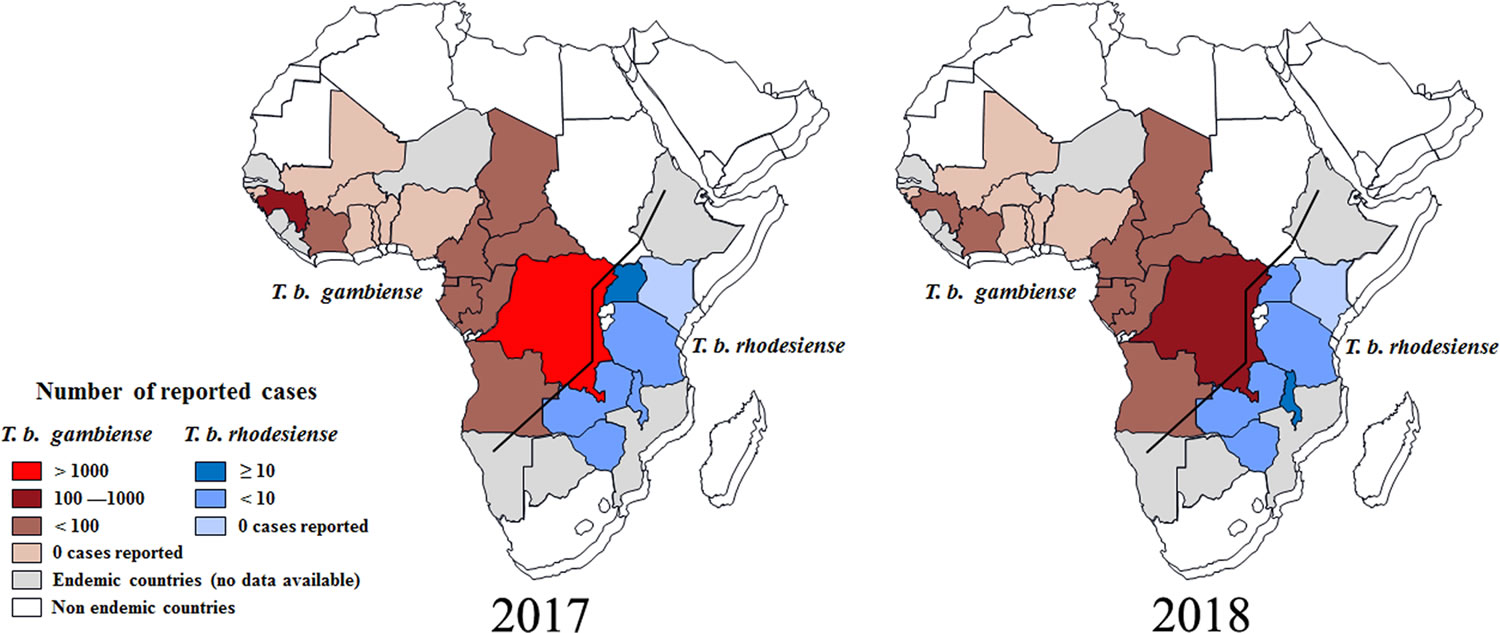
[Source 11 ]Figure 2. Distribution of human African trypanosomiasis in endemic countries (2017 and 2018)
Major human epidemics
There have been several epidemics in Africa over the last century:
- one between 1896 and 1906, mostly in Uganda and the Congo Basin;
- one in 1920 in a number of African countries; and
- the most recent epidemic started in 1970 and lasted until the late 1990s.
The 1920 epidemic was controlled thanks to mobile teams which carried out the screening of millions of people at risk. By the mid-1960s, the disease was under control with less than 5000 cases reported in the whole continent. After this success, surveillance was relaxed, and the disease reappeared, reaching epidemic proportions in several regions by 1970. The efforts of World Health Organization (WHO), national control programmes, bilateral cooperation and nongovernmental organizations (NGOs) during the 1990s and early 21st century reversed the curve.
Since the number of new human African trypanosomiasis cases reported between 2000 and 2012 dropped significantly as a result of international coordinated efforts, the WHO neglected tropical diseases road map targeted its elimination as a public health problem by 2020 and interruption of transmission (zero cases) for 2030.
African sleeping sickness transmission cycle
In both forms of human African trypanosomiasis, infection occurrence depends on the interaction of three elements within a particular environment 6:
- The mammalian reservoirs of parasites (human or animal) that can be also the host suffering from the disease, and which are influenced by their behavioral interactions with the environment;
- Tsetse flies or Glossina species as cyclical vectors for transmission that are fully dependent on environmental factors; and
- The pathogenic parasite, the trypanosome.
Due to reasons that are not always well known, but that are related to the interactions between these three elements, the transmission of African sleeping sickness is confined in areas with quite clear spatial limits, beyond which the disease does not occur. This limited space is called a “focus” 12.
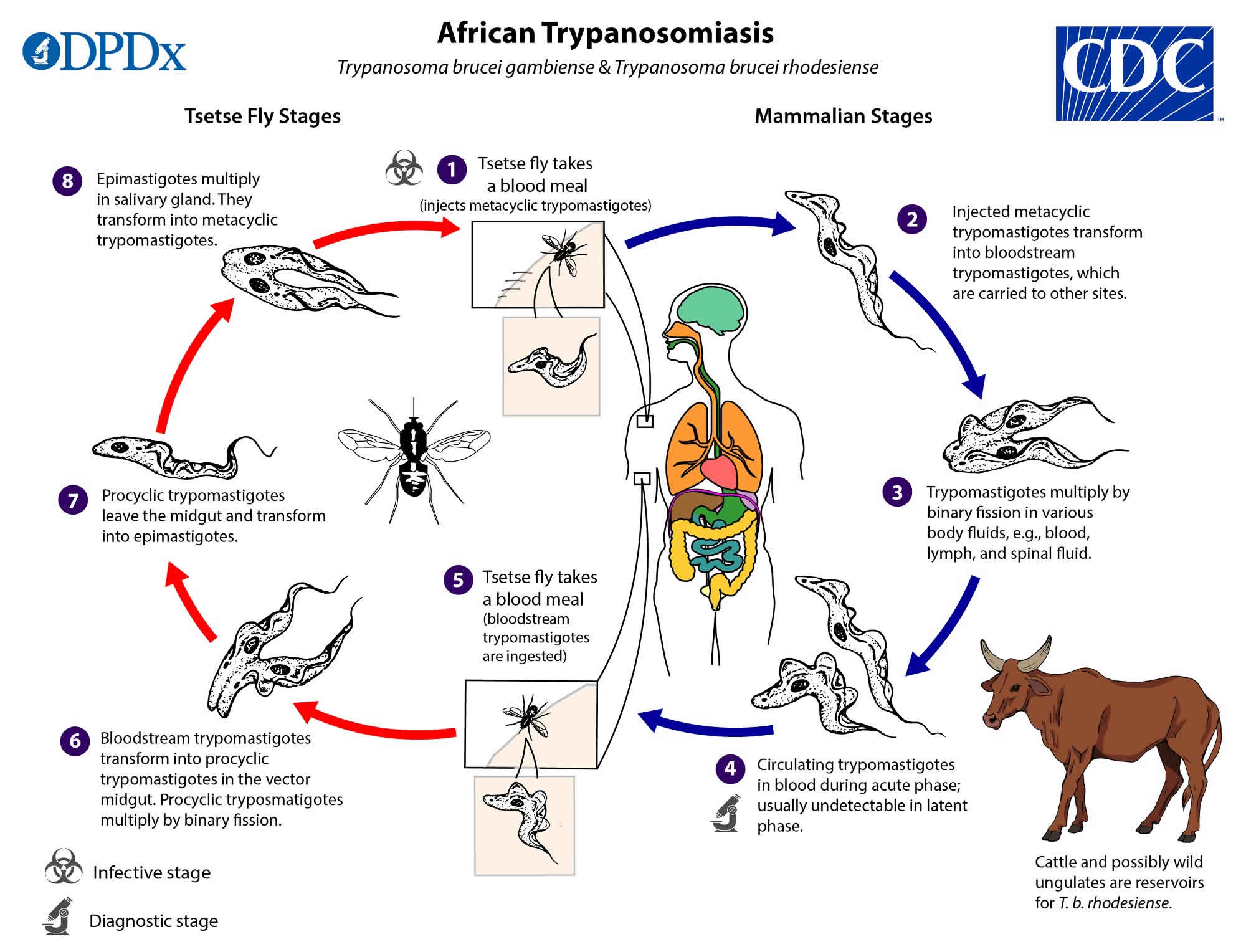
Figure 3. African trypanosomiasis life cycle
The parasite (Trypanosoma brucei)
Human African trypanosomiasis is caused by protozoans hemoflagellates of the genus Trypanosoma. There are many species of trypanosomes, but only two subspecies of the Trypanosoma brucei group are responsible for sleeping sickness: Trypanosoma brucei gambiense and Trypanosoma brucei rhodesiense 7. A third subspecies of the group, Trypanosoma brucei brucei is nonpathogenic for humans, but parasitizes domestic (Bovidae, Suidae, dogs) and wild animals and it is widely used in experimental models of human African trypanosomiasis 13. The Bovidae comprise the biological family of cloven-hoofed, ruminant mammals that includes cattle, bison, buffalo, antelopes, and goat-antelopes. Suidae is a family of artiodactyl mammals which are commonly called pigs, hogs, or boars.
Trypanosomes are extracellular parasites and they can be clearly identified microscopically, but the two subspecies that are pathogenic for human beings are morphologically indistinguishable 7. Since differences between these subspecies cannot be observed by microscopy, other methods have been used; apart from the geographic differences, the specific molecular marker known as the serum resistance-associated (SRA) gene is currently being used to differentiate between the species, as it is always present in all Trypanosoma brucei rhodesiense isolates.14 Trypanosoma brucei gambiense has been characterized by another specific molecular marker, the TgsGP gene, found in type 1 Trypanosoma brucei gambiense 14. This form constitutes the classical form of Trypanosoma brucei gambiense human African trypanosomiasis. A much more uncommon type 2 Trypanosoma brucei gambiense found in western and central Africa does not contain the TgsGP gene16 and it produces a more acute form of gambiense human African trypanosomiasis 6.
Trypanosoma brucei group trypanosomes are transmitted by tsetse flies. They have a complex life cycle with differentiated biological stages in both the insect vector and the mammalian host 15. When entering in the Glossina (tsetse fly), the parasites are ingested as bloodstream trypomastigote forms and they move to the midgut (see Figure 2). Some species of tsetse are refractory to infection by specific species of trypanosome and even when they are susceptible, the population of parasites in the midgut can be reduced making unsustainable the infection at this point 16. However some of the trypomastigote forms in the insect’s midgut may arrive to differentiate into procyclic forms, which replicate in situ and cross the peritrophic membrane to reach the proventriculus, where they becomes mesocyclic trypomastigotes and later epimastigote forms 15. Then, they migrate via the esophagus, proboscis, and hypopharynx to the salivary gland, where they are able to multiply and some of them can transform into infectious metacyclic forms 16. During this migration from the midgut to the salivary glands, the parasite population size experiences a pronounced reduction 16. The metacyclic form is the only stage that is infective to vertebrates, and it is characterized by the presence of the variant surface glycoprotein coat that will protect the parasite in order to survive on the host 17. The whole cycle in the vector takes 18–35 days, and once infected, a tsetse fly remains so for the rest of its lifespan. Nevertheless, in lab conditions it has been observed that the majority of ingested trypanosomes fail to develop, and only 2%–5% of the flies ingesting trypanosomes produce metacyclic forms 18.
Metacyclic forms are injected subdermally in the mammalian host during a tsetse fly’s blood meal. They proliferate at the site of inoculation and they transform in long slender forms, as they are carried by the draining lymph nodes to the bloodstream where they replicate 17. In the blood stream, the parasite can be found in its proliferative, long, slender bloodstream trypomastigote form, which is adapted to maintain the parasite in mammalian blood, or as the nonproliferative, short, stumpy bloodstream trypomastigote form, which is adapted to differentiate into the replicative procyclic form in the tsetse fly, and thus to ensure transmission 17. There are also intermediate forms, which are transitional, as they are between the slender and stumpy forms. The bloodstream forms can enter into different body fluids, including lymph and cerebrospinal fluid, and they can also cross the placenta 19.
Sexual reproduction is not obligatory in trypanosomes, but it can occur in salivary glands, yielding the possibility of genetic exchange and the rapid transmission of important characteristics, such as drug resistance and virulence 20. Genetic exchange is relatively frequent in Trypanosoma brucei rhodesiense, but it is more sporadic in Trypanosoma brucei gambiense 21.
The amount of parasites in the bloodstream (parasitemia) is generally very low in Trypanosoma brucei gambiense infections, often remaining below 100 parasites per mL of blood, while in Trypanosoma brucei rhodesiense, infections typically present with higher parasitemia 22. Nevertheless, in both cases, parasitemia undergoes important fluctuations; low parasitemia levels can make it complicated to see parasites through microscopy.
The fluctuations of parasitemias are related to host immune responses and the antigenic variation presented by the parasites 23. The surface of bloodstream forms of trypanosomes is covered with a glycoprotein coat (variant surface glycoprotein), which keeps antibodies and the complement system away from the parasite membrane 24. The host develops an effective antibody response, mainly through immunoglobulin M (IgM) and immunoglobulin G (IgG), against this highly immunogenic variant surface glycoprotein. Most antibodies of the host are able to disrupt the protective shield and kill the parasites, decreasing the number of parasites present. Nevertheless, the coat protein can turn over very rapidly, leading antibodies that are bound to the surface to be wiped out and permitting the parasites to escape the immune response at a given time. There are a large number of genes with the potential to encode the variant surface glycoprotein, carrying the possibility of producing new variants almost ad infinitum 25. There is also an internal mechanism that regulates shifting to new variants: the parasites release a “stumpy induction factor”, which triggers the production of nonreplicating stumpy forms, adapted to reestablish the life cycle when ingested by the tsetse fly 26.
The vector (tsetse flies)
Human African trypanosomiasis is transmitted through the bite of an infected tsetse fly (genus Glossina). Thirty-one species and subspecies of tsetse flies have been described and classified in three groups or subgenera, which are generally related to different habitats 27:
- Subgenus Nemorhina or the palpalis group, found in western and central Africa, live in vegetation close to a water source, such as forests, gallery forests, riverbanks and lakes, swamps and mangroves, coffee or cocoa plantations, and they can even adapt to environmental changes, surviving in the periurban areas of medium and large towns36 and areas of intensive agriculture. This subgenus includes the main vectors of sleeping sickness: Glossina palpalis palpalis and Glossina palpalis gambiensis for Trypanosoma brucei gambiense and Glossina fuscipes for both Trypanosoma brucei gambiense and Trypanosoma brucei rhodesiense. Glossina palpalis is distributed in the Atlantic coast from Senegal to Angola. Glossina fuscipes is present in central Africa from Cameroon and Congo to the Rift Valley.
- Subgenus Glossina sensu stricto or the morsitans group, occurs in woodland savannah, and is linked to the presence of wild fauna and cattle. Glossina morsitans, Glossina swynnertoni, and Glossina pallidipes, all of which are located mainly in East Africa, might be involved in the transmission of Trypanosoma brucei rhodesiense.
- Subgenus Austenina or the fusca group, lives in primary forest belts (rainforest, savannah, or coastal forests). Increasing human activity in these forests tends to make this subgenus disappear. They have not been reported to be vectors of human African trypanosomiasis.
Figure 4. Tsetse fly (genus Glossina)

Tsetse flies are restricted to sub-Saharan Africa, between the latitudes of 14° North and 29° South, south of the Sahel desert, and north of the Namibian and Kalahari deserts 28. Some pockets of Glossina morsitans and Glossina fuscipes have been described in southwestern Saudi Arabia 29. All the species of tsetse flies are potentially cyclical vectors of trypanosomes, but in nature, the infection is carried almost exclusively by Glossina fuscipes, Glossina palpalis, and Glossina morsitans.
Tsetse flies need a particular temperature (16°C–38°C) and humidity (50%–80% of relative humidity) to survive. Therefore, they are linked to the presence of water that increases the local humidity, allowing for the growth of vegetation that protects them from direct sunlight and wind, and attracts the animals to where tsetse feed 7.
The transmission of human African trypanosomiasis involves the obligatory biological development of trypanosomes in the fly and in only 2%–5% of cases will this development occur, with the presence of metacyclic forms in the salivary glands. Therefore, the mature infection rate among tsetse flies is quite low, with an average of less than 1% of tsetse flies infective for Trypanosoma brucei species 30. This makes the tsetse fly a relatively low competent vector for trypanosomiasis transmission. Nevertheless, even the low ingestion of parasites during the blood meal can result in infection in the fly 31 and a single infected tsetse bite is sufficient for transmitting the infection to another mammalian host 32. However, the probability of the transmission of human African trypanosomiasis is related to the number of trypanosomes inoculated into the mammal. Therefore, the capacity of transmission of the disease by the tsetse fly is influenced by different factors, such as the density of tsetse populations, the tsetse fly’s longevity, the vector’s susceptibility to infection, the tsetse fly’s infestation rates, the availability of other sources of a blood meal, and the factors that determine the frequency and intensity of the host-fly’s contacts 33.
The life cycle of Glossina is peculiar because of the absence of oviposition and the development of a single larva in the uterus of the female fly. Each female produces one single offspring at a time and the third instar larva developed within the uterus is larviposited as a mature larva into humid soil in a shady place. The larva moves actively, and then it quickly buries itself a few centimeters deep in clay or sandy soil to pupate. The pupa lives in the soil on the food reserves, and the adult insect emerges 20–80 days after larviposition, depending on temperature and humidity 7.
The female is mated in the days following her emergence, and only one insemination is generally sufficient for her to remain fertile, but in some cases, young females accept several sexual unions. The female accumulates the spermatozoids in the spermathecae where they can survive for nearly 200 days. The female deposits a larva approximately every 10 days 7.
The lifespan of the fly is highly variable depending on the season: longer in the rainy season (3–5 months) and shorter in the dry season (1–2 months), rarely exceeding 7 months. In general females live longer than males, being more numerous in natural populations 7.
Both female and male tsetse flies feed on blood (hematophagous) and, thus, both are capable of transmitting sleeping sickness disease. Newly emerged flies (teneral flies) need to immediately find a host on which to feed, and they are not very discriminatory about the host. The chance of becoming infected is highest in this first blood meal, but tsetse flies can be infected with trypanosomes at any point in their lives 34. Meals are taken about every 2–4 days (up to 10 days when climatic conditions or the availability of hosts is adverse) 7. Tsetse fly saliva is injected during the blood meal to avoid blood coagulation and to produce a vasodilation. If the saliva contains infective trypanosomes, they are transferred during the meal. If the host is infected by trypanosomes, these are ingested by the fly and it can develop an infection but as already indicated the majority of ingested trypanosomes fail to develop.
The feeding preferences of tsetse flies are not well known; odor stimuli produced by cutaneous secretions, feces, urine, and breath, as well as visual factors related to form, size, movements, contrast, and color are involved in the finding of a suitable host 35. These aspects have led to the development of different vector control tools 36. Suidae (pigs, warthogs, and bush pigs) are regarded as the first preference for Glossina morsitans and Glossina palpalis, and reptiles (monitor lizards, crocodiles, and snakes) are preferred by many species, but in general, tsetse flies are opportunistic and are able to adapt to host availability 7. Some wild animals (kob, zebra, wildebeest, oryx) are rarely ever bitten by tsetse flies, possibly because their colors are less attractive, or because their skin contains repellent substances 37. Humans are occasionally bitten by the morsitans group, although the human odor acts as a repellent 38. Glossina fuscipes and Glossina palpalis seem to be attracted to human odor 39. The palpalis group is more opportunistic, and it feeds more indiscriminately on many species 40.
The blood meal lasts between 20–30 seconds 7. The tsetse flies can be killed by the defensive movements of the host. Thus, they prefer to bite animal hosts on the lower parts of the anterior legs or on the belly, where they are less likely to be killed 7. This behavior is the basis upon which to propose the restrictive application of insecticides to only those parts of cattle to decrease costs and increase effectiveness 41.
The tsetse fly spends most of its time resting, with daily flying activity of 30–50 minutes in males and only a few minutes in females, mainly concentrated in the morning and at the end of the afternoon; the flies do not leave their resting places during the hottest parts of the day or during the rain, or in windy conditions 42. They fly at low levels (~0.5 m) and at high speeds (up to 25 km/hour) in small successive displacements of a few seconds or minutes, usually covering short distances (200–300 m). However, they have a great capacity to disperse, and they can cover up to 20–30 km of distance in successive days 22. The tsetse flyies can be passively transported by cattle herds or on vehicles, boats, or trains, with the risk of reinvading areas free of tsetse 7.
Resting places include the underside of leaves, branches, the holes in tree trunks, roots, or other shady areas, but they also rest in sunny places in the early morning 7. During the dry season, the resting places are below 50 cm during the day, but at night or during the rainy season, they are at around 2.5 m, sometimes reaching up to 10 m. Knowledge about the resting places of tsetse flies determines the use of the selective application of insecticide ground spraying and sequential insecticide aerial treatments 7.
Densities of tsetse populations are determined by climatic, environmental factors and host availability, but in general, the tsetse population growth tends to be low, as this is mainly related to their low reproductive rate 7. Since Glossina is practically the only vector responsible for transmitting the parasite from one mammalian host or reservoir to another, combining its control with the detection and treatment of cases of infection can contribute to the sustainability of human African trypanosomiasis control.
The reservoir and the host – the human being in the cycle of human African trypanosomiasis
The role of animal reservoirs and human beings is very different in the two forms of human African trypanosomiasis. Gambiense human African trypanosomiasis is an anthroponotic disease with a minor role for animal reservoirs, while rhodesiense human African trypanosomiasis is a zoonotic disease, affecting mainly animals (livestock and game); humans are only accidental hosts.
Gambiense human African trypanosomiasis
It is typically regarded that humans constitute the main epidemiological reservoir of Trypanosoma brucei gambiense 7. Despite the fact that the tsetse fly has a relatively low competence as vector for gambiense human African trypanosomiasis, and the presence of parasitemias in human beings is usually low, the long duration of human infection with an extended paucisymptomatic period is considered sufficient to maintain a human–fly–human transmission cycle 43.
Nevertheless, some data suggest the possible role played by asymptomatic human beings or animals as additional reservoirs 44. Asymptomatic individuals infected with trypanosomes who refused treatment and were followed-up for long periods (5–15 years) without showing any clinical manifestation of the disease have been described 45. These individuals can become aparasitemic by microscopy, while keeping a polymerase chain reaction intermittently positive and a trypanolysis test positive 46. They could be asymptomatic chronic carriers of the infection, and a source of vector infection that maintains the transmission of gambiense human African trypanosomiasis in some cases 47. This could be similar to the phenomena of trypanotolerance, which is well described in animals 48.
The other hypothesis relies on the possible role played by a variety of domestic and wild animals (mammals and reptilians) as reservoirs. Trypanosoma brucei gambiense has been described in domestic animals such as pigs, sheep, dogs, goats, and many different wild animals 49. The role played as reservoirs by these animal hosts is not clear; in some gambiense human foci, Trypanosoma brucei gambiense group 1 parasites are found in domestic livestock (mainly pigs), but in other foci where the same infection is present in humans, the parasites have not been found in domestic animals 50. On the other hand, in some foci, the infection rates and trypanosome genotypes circulating in pigs and humans are different, suggesting that the pig may not act as an animal reservoir for human infections in this focus 51. There are also some studies relating the prevalence of gambiense infection in domestic animals and that of gambiense human African trypanosomiasis in humans 52 and which have not found the parasite in wild animals in sites where there are no human cases 53. Some studies show that experimental infections of animals with Trypanosoma brucei gambiense are limited in time, always lasting less than 1 year 54. Therefore, more data are needed to clarify the role played by the animal reservoir in maintaining gambiense transmission when the prevalence of human infection is low and elimination is envisaged.
In addition, human African trypanosomiasis control programs that target only the human reservoir through case detection and the subsequent treatment of cases without including vector control activities have been very successful, as the disease has reached very low prevalence rates after several rounds of active case finding 55 and, in some cases, achieving even elimination 56. However, in other foci, despite active case detection that has been maintained over many years, ongoing transmission continues and the prevalence figures have remained the same 57. In these cases, the persistence of the transmission and maintenance of the prevalence rate may also be linked to incomplete attendance of population to screening activities and to the limited sensitivity of the diagnostic methods used 58.
Rhodesiense human African trypanosomiasis
The transmission cycle of Trypanosoma brucei rhodesiense mainly involves the transmission between nonhuman reservoirs through the tsetse fly, but occasionally humans can be included in the cycle, although their role as reservoirs is limited 6:
- Animal–tsetse fly–animal: this is the usual cycle.
- Animal–tsetse fly–human: this occasionally happens.
- Human–tsetse fly–human: this is very unlikely and probably only happens during epidemics.
The population of Trypanosoma brucei rhodesiense is, therefore, maintained in nonhuman reservoirs. In some cases, the animal just carries the parasite and survives for many years, but in other cases, the animal is affected by the disease. Carriage of Trypanosoma brucei rhodesiense has been confirmed in the following wild animals: bushbuck; duiker; giraffe; hartebeest; hyena; impala; lechwe; lion; oribi; reedbuck; warthog; waterbuck; and zebra, with bushbucks considered as the most frequently described reservoir 7. Domestic animals, such as bovine and porcine species, have also been identified as reservoirs 59. The existence of a reptilian host has been well described, but its contribution to the maintenance of trypanosomes is unlikely, as their body temperature varies considerably, making the survival of parasites improbable 60.
Hence, two situations can be differentiated: there may be areas where the main reservoirs are wild animals; and there may be areas where domestic animals play this role. These two different situations are related to different risks of transmission, and these risks are crucial when considering control measures.
In the case of areas where wild animals can act as the main reservoir, different factors can facilitate the involvement of the human being in the transmission cycle of rhodesiense African sleeping sickness. These factors are related to the increase of contact between human and wild animal reservoirs. Land-use pressures trigger the use of new areas for domestic animal grazing; thus, domestic animals invade areas that are occupied by wildlife, increasing the possibility of the transmission of rhodesiense African sleeping sickness from wildlife to humans, either directly or passing the disease through livestock. Another example is the case described where tourists and rangers visit or work in protected areas 61. In some areas, wild animals formerly acted as the main reservoir for the parasite, but the habitats of these animals were colonized by human activities, and domestic animals substituted wildlife, subsequently becoming the main reservoirs 7.
In the case where cattle serve as the main reservoir, the contact between humans and the reservoir is easier, and outbreaks of the disease in humans can easily occur. This contact can be even peridomestic. Cattle infection with Trypanosoma brucei rhodesiense has accounted for around 1% of livestock infections in areas where human cases are rarely reported 59, but this rate can reach 18% during human outbreaks 62.
African sleeping sickness causes
African sleeping sickness is mostly transmitted through the bite of an infected tsetse fly but there are other ways in which people are infected:
- Mother-to-child infection: the trypanosome can cross the placenta and infect the fetus.
- Mechanical transmission through other blood-sucking insects is possible, however, it is difficult to assess its epidemiological impact.
- Accidental infections have occurred in laboratories due to pricks with contaminated needles.
- Transmission of the parasite through sexual contact has been reported.
Once the tsetse fly ingests the trypanosomes, they multiply and develop into epimastigotes. Humans are infected after the bite from a tsetse fly. The injected parasites then rapidly divide in the bloodstream and lymphatics. Eventually, the parasite enters the central nervous system (brain and spinal cord) and causes neurological and behavioral symptoms.
In the first stage, the trypanosomes multiply in subcutaneous tissues, blood and lymph. This is also called hemo-lymphatic stage, which entails bouts of fever, headaches, enlarged lymph nodes, joint pains and itching
In the second stage the parasites cross the blood-brain barrier to infect the central nervous system (CNS). This is known as the neurological or meningo-encephalic stage. In general this is when more obvious signs and symptoms of the disease appear: changes of behaviour, confusion, sensory disturbances and poor coordination. Disturbance of the sleep cycle, which gives the disease its name, is an important feature. Without treatment, sleeping sickness is considered fatal although cases of healthy carriers have been reported.
The trypanosomes evade the host’s immune system because of extensive antigenic variation of the glycoproteins located on the surface of the parasite. During this time, the parasites invade almost every organ in the body.
Some individuals may develop a severe hypersensitivity reaction to the parasite that leads to itching, swelling, and edema.
In the liver, there may be portal infiltration and fatty degeneration.
When the heart is invaded, arrhythmias may develop leading to death.
When the brain is involved, it may lead to meningoencephalitis, bleeding, edema, and granulomatous lesions.
Risk factors for getting African trypanosomiasis
The risk factors for the transmission of African sleeping sickness are determined by the increases in the possibility with which humans come into contact with a tsetse fly, and these factors are thus related to the site of contact between the tsetse fly and the human, and the intensity and frequency of this contact. In rhodesiense African sleeping sickness, these risk factors are also related to the presence of nonhuman reservoirs.
Gambiense human African trypanosomiasis
The risk factors for gambiense African sleeping sickness vary according to the different environmental settings and the characteristics of the vector in the different biotopes, and the activities carried out by human beings in the biotopes occupied by the tsetse flies.
- In humid forest, the tsetse flies are widely distributed, and human–fly contact is related to activities such as hunting, fetching firewood, timber-related activities, and forest clearing for farming 63.
- In the woodland savanna and riverine forest galleries, the fly is found close to rivers and streams. The risk of transmission has been associated with activities that have been developed along these water bodies, such as fetching water, washing clothing or food (cassava), the artisanal extraction of palm oil, brewing, gold and diamond mining, and fishing 64. The risk of transmission increases when tsetse habitats are restricted, for example, during the dry season.
- In the transitional vegetation between forest and woodland savanna, the islands of vegetation provide a suitable habitat for tsetse, and these locations act as points from which hosts are sighted. These areas are often used for farming, making this activity a risk factor in these areas 65.
- In the mangrove areas, tsetse flies find a favorable habitat where a high risk of transmission is associated with fishing and crustacean collection, but the parasite is also found in cleared areas used for rice cultivation. Pirogue jetties and fishing encampments are areas where human–fly contact can be intense 66.
- Coca, coffee, and also mango and banana plantations, where the original forest has been replaced, are also suitable habitats for tsetse flies, and these areas are related to transmission in plantation workers 67.
- Gambiense African sleeping sickness is considered a rural disease, but transmission has also been occasionally observed in urban settings. Nevertheless, transmission in urban areas is associated with travels to neighboring rural areas for cultivating fields, or it may occur in suburban outskirts closer to transitional vegetation areas, where agricultural activities are possible; these areas constitute a suitable tsetse habitat with few alternative hosts 68.
It has been described that certain protective immunity exists against new infections in humans after suffering from the disease 69.
The risk associated with age- and sex-related factors pertains to the activities and behaviors made by the different age and sex groups. In general, gambiense African sleeping sickness is predominately a disease of adults, mainly affecting young adults, as this is the group that is most involved in productive activities that facilitate contact with the vector 70. Children are usually less affected than adults, but in some areas (such as mangroves), teens present a higher rate of infection related to fishing and leisure activities in water areas.109,110 In areas where at-risk activities include mining, hunting, or fishing, the prevalence is higher in males. In transitional vegetation areas where the risk of infection is associated with agriculture and domestic activities at bodies of water, similar prevalence rates have been found in males and females 71.
The clustering of cases and some familial aggregation has been described, as the risk of gambiense African sleeping sickness for a child significantly increased when the mother also had African sleeping sickness 22. It has been suggested that familial clustering was a consequence of similar exposure to the vector and shared behavioral risk factors, rather than of genetic susceptibility 72.
The risk of gambiense African sleeping sickness infection in short-term travelers from nonendemic areas is very low, as tourists rarely visit the rural areas where gambiense African sleeping sickness is endemic. Gambiense African sleeping sickness cases that are occasionally diagnosed in nonendemic countries are mainly seen in immigrants and expatriate residents living in at risk areas for extended periods 73.
Rhodesiense human African trypanosomiasis
The two different settings described for rhodesiense African sleeping sickness mean different situations of risk for contracting the disease.
- In the areas where wildlife is the main reservoir, the main risk factors are associated with entry into areas that are usually restricted and where wildlife is preserved (national parks and game reserves). A high density of tsetse flies in these areas increases this risk. Exposure is related to the movement of humans, animals, and tsetse flies out of or into reserves, especially during specific seasons of the year. Populations living in the periphery of game reserves and national parks are the ones at the highest risk 74. The exposure to rhodesiense African sleeping sickness in this area involves wildlife conservation activities (for example, rangers and park wardens), hunting and poaching, fishing, honey and firewood collection, and visitors of national parks (tourists) 75. The movement of livestock for grazing in these areas is another risk factor for herdsmen 76; there is also the potential that this risk could be transferred to other areas where the cattle is moving.
- In the areas where livestock is the main reservoir, activities linked to cattle raising increase this risk, but the general population living in these areas is also exposed. Those living on the periphery of villages or near the cattle markets are most at risk 75.
With respect to age, those at highest risk for rhodesiense African sleeping sickness are in the working-age group, which is comprised of individuals venturing into tsetse habitats 75. It is assumed that their contact with flies occurs when the humans enter into the woodland habitat of the flies, but recent studies have found the frequent peridomestic presence of some tsetse species, and even tsetse biting humans inside buildings 77, but transmission still mainly occurs in more tsetse-suitable habitats. As is the case in gambiense African sleeping sickness, the sex-based risks are related to the specific activities and behaviors engaged upon by the members of each sex; in general, rhodesiense African sleeping sickness is predominately a disease of males 75. Familial aggregation also occurs in rhodesiense African sleeping sickness 75, which is most likely related to the common exposure of vectors, especially since there are shared behavioral and spatial risk factors among members of a household.
Given that rhodesiense African sleeping sickness is an acute disease, it is possible to observe a certain seasonal variation in transmission that is linked to the density of Glossina: The peak in the Glossina population densities is usually seen after the rainy season, and the peak in the human cases can be detected in 1–3 months after the rainy season.119
The risk of acquiring rhodesiense African sleeping sickness for short-term travelers from nonendemic areas is low, but this risk factor is more important than that of developing gambiense African sleeping sickness, as tourists commonly visit natural areas where rhodesiense African sleeping sickness is transmitted 73. Cases of rhodesiense African sleeping sickness in travelers can occasionally appear in clusters of people who have visited common areas 78.
African sleeping sickness signs and symptoms
The clinical course of human African trypanosomiasis has two stages. In the first stage, the parasite is found in the peripheral circulation, but it has not yet invaded the central nervous system. Once the parasite crosses the blood-brain barrier and infects the central nervous system, the disease enters the second stage. The subspecies that cause African trypanosomiasis have different rates of disease progression, and the clinical features depend on which form of the parasite (Trypanosoma brucei rhodesiense or Trypanosoma brucei gambiense) is causing the infection. However, infection with either form will eventually lead to coma and death if not treated.
African sleeping sickness general symptoms include:
- Mood changes, anxiety
- Fever, sweating
- Headache
- Weakness
- Insomnia at night
- Sleepiness during the day (may be uncontrollable)
- Swollen lymph nodes all over the body
- Swollen, red, painful nodule at site of the fly bite
Trypanosoma brucei rhodesiense infection (East African sleeping sickness) progresses rapidly. In some patients, a large sore (a chancre) will develop at the site of the tsetse bite. Most patients develop fever, headache, muscle and joint aches, and enlarged lymph nodes within 1-2 weeks of the infective bite. Some people develop a rash. After a few weeks of infection, the parasite invades the central nervous system and eventually causes mental deterioration and other neurologic problems. Death ensues usually within months.
Trypanosoma brucei gambiense infection (West African sleeping sickness) progresses more slowly. At first, there may be only mild symptoms. Infected persons may have intermittent fevers, headaches, muscle and joint aches, and malaise. Itching of the skin, swollen lymph nodes, and weight loss can occur. Usually, after 1-2 years, there is evidence of central nervous system involvement, with personality changes, daytime sleepiness with nighttime sleep disturbance, and progressive confusion. Other neurologic signs, such as partial paralysis or problems with balance or walking may occur, as well as hormonal imbalances. The course of untreated infection rarely lasts longer than 6-7 years and more often kills in about 3 years.
West African trypanosomiasis signs and symptoms
West African trypanosomiasis symptoms may be minimal or intermittent during the first months of infection. They are usually apparent within a few months to a year after getting an infected tsetse fly bite.
Occasionally, within 1 to 3 weeks, the infective bite develops into a red sore, also called a chancre. Several weeks to months later, other symptoms of sleeping sickness occur. These include fever, rash, swelling of the face and hands, headaches, fatigue, aching muscles and joints, itching skin, and swollen lymph nodes. Weight loss occurs as the illness progresses. Progressive confusion, personality changes, daytime sleepiness with nighttime sleep disturbances, and other neurologic problems occur after the infection has invaded the central nervous system. These symptoms become worse as the illness progresses. If left untreated, death will eventually occur after several years of infection.
East African trypanosomiasis signs and symptoms
East African trypanosomiasis symptoms usually develop within 1 to 3 weeks after an infective tsetse fly bite. A bite by the tsetse fly is often painful and can develop into a red sore, also called a chancre. Fever, severe headaches, irritability, extreme fatigue, swollen lymph nodes, and aching muscles and joints are common symptoms of sleeping sickness. Some people develop a skin rash. Progressive confusion, personality changes, and other neurologic problems occur after infection has invaded the central nervous system. If left untreated, infection becomes worse and death will occur within months.
African trypanosomiasis complications
African sleeping sickness complications include:
- Injury related to falling asleep while driving or during other activities
- Gradual damage to the nervous system
- Uncontrollable sleep as the disease gets worse
- Coma
African sleeping sickness diagnosis
If you suspect that you may have West African trypanosomiasis, see your health care provider who will order several tests to look for the parasite. Common tests include examination of blood samples and a spinal tap. Your physician may also take a sample of fluid from swollen lymph nodes.
African sleeping sickness Trypanosoma brucei gambiense can be screened for with the card agglutination trypanosoma test (CATT), a study that examines serum for antigen and that carries a sensitivity of 91% and specificity of 97% 79. Both species can also be identified with Giemsa-stained blood, lymph node aspirates, and cerebrospinal fluid (CSF) fluid. All patients with suspected African sleeping sickness should be screened for central nervous system (CNS) involvement with a lumbar puncture; CSF fluid should be tested for trypanosomes, leukocytosis, and trypanosome IgM. In the second-stage infections, the number of parasites in congestive heart failure (CHF) can be very low. World Health Organization (WHO) diagnostic criteria for suspected second stage trypanosomiasis, therefore, consists of either the presence of trypanosomes in CSF fluid or greater than 5 white blood cells (WBCs) per microliter of fluid in a suspected case.
In many hospitals, in Africa, a blood smear is often done as it will reveal the mobile trypanosomes. Blood smears are often positive in early disease when the number of circulating parasites is very high. The parasite load in Trypanosoma brucei rhodesiense infection is substantially higher than the level in Trypanosoma brucei gambiense infection.
Trypanosoma brucei rhodesiense parasites can easily be found in blood. They can also be found in lymph node fluid or in fluid or biopsy of a chancre. Serologic testing is not widely available and is not used in the diagnosis, since microscopic detection of the parasite is straightforward.
For East African trypanosomiasis, a skin biopsy may be done if you have a chancre.
Lymph node aspiration is sometimes done to identify the parasite and may yield positive results.
CT scan and MRI of the brain frequently reveal massive cerebral edema and enhancement of the white matter.
The classic method for diagnosing Trypanosoma brucei gambiense infection is by microscopic examination of lymph node aspirate, usually from a posterior cervical node. It is often difficult to detect Trypanosoma brucei gambiense in blood. Concentration techniques and serial examinations are frequently needed. Serologic testing is available outside the U.S. for Trypanosoma brucei gambiense; however, it normally is used for screening purposes only and the definitive diagnosis rests on microscopic observation of the parasite.
All patients diagnosed with African trypanosomiasis must have their cerebrospinal fluid examined to determine whether there is involvement of the central nervous system, since the choice of treatment drug(s) will depend on the disease stage. The World Health Organization criteria for central nervous system involvement include increased protein in cerebrospinal fluid and a white cell count of more than 5. Trypanosomes can often be observed in cerebrospinal fluid in persons with second stage infection.
African sleeping sickness treatment
African trypanosomiasis treatment depends on the form of the disease (Trypanosoma brucei gambiense or Trypanosoma brucei rhodesiense) and the disease stage (i.e. whether the central nervous system has been invaded by the parasite). The earlier the African trypanosomiasis is identified, the better the prospect of a cure. The early stage management requires treatment of fever and malaise. Close monitoring of the central nervous system status is necessary. Sometimes, patients may require intubation and mechanical ventilation as they can not maintain a patent airway 80.
There is no test of cure for African trypanosomiasis. Patients should be followed up to 24 months with a lumbar puncture every 6 months (or sooner, if symptoms return) for 2 years after treatment to detect a relapse should it occur, as parasites may remain viable for long periods and reproduce the disease months after treatment.
Treatment success in the second stage depends on drugs that cross the blood-brain barrier to reach the parasite.
New treatment guidelines for gambiense human African trypanosmiasis were issued by World Health Organization (WHO) in 2019. In total six different drugs are used for the treatment of sleeping sickness. These drugs are donated to WHO by manufacturers and distributed free of charge to disease endemic countries.
Pentamidine, which is the recommended drug for first stage Trypanosoma brucei gambiense infection, is widely available in the U.S. The other drugs (suramin, melarsoprol, eflornithine, and nifurtimox) used to treat African trypanosomiasis are available in the U.S. only from the Centers for Disease Control and Prevention (CDC). Physicians can consult with CDC staff for advice on diagnosis and management and to obtain otherwise unavailable treatment drug.
Pentamidine, given by intravenous infusion over 2 hours or by intramuscular injection, is used to treat first stage Trypanosoma brucei gambiense infection. It is generally well tolerated, but adverse reactions of hypoglycemia, injection site pain, diarrhea, nausea and vomiting occur. Suramin is used to treat first stage Trypanosoma brucei rhodesiense. Suramin is also effective against Trypanosoma brucei gambiense, but it is not often used because severe reactions occur in persons who are co-infected with Onchocerca volvulus. Adverse reactions to suramin are frequent, but usually mild and reversible. These include drug rash, nephrotoxicity, and peripheral neuropathy. In rare instances, suramin administration results in a hypersensitivity reaction, and, for this reason, a small test dose is usually given prior to the full first dose.
Second stage Trypanosoma brucei gambiense is treated with eflornithine, which is given in 4 intravenous infusions daily for 14 days. Adverse effects of eflornithine include bone marrow suppression, gastrointestinal symptoms, and seizures. Eflornithine is highly effective, but the difficulty in administering 4 infusions daily in rural African facilities has led to the use of eflornithine (dosed less frequently) in combination with nifurtimox. The efficacy of the combination regimen appears to be at least as high as eflornithine monotherapy. Eflornithine is not effective against Trypanosoma brucei rhodesiense and it is not recommended for treating the East African form of the disease. Melarsoprol, an organoarsenic compound, is the only drug available for treating second stage Trypanosoma brucei rhodesiense. Adverse reactions to melarsoprol can be severe and life-threatening. An encephalopathic reaction occurs in 5-10% of patients with a case-fatality rate of approximately 50% when it occurs. Prednisolone is often given to patients who are being treated with melarsoprol to reduce the risk of encephalopathy. Other adverse reactions observed with melarsoprol include skin reactions, gastrointestinal upset, and peripheral neuropathy. Intravenous injections of melarsoprol are painful and can cause phlebitis. The drug is administered by use of lengthy and complicated dosing schedules, however, an abbreviated 10-day regimen appears promising.
African sleeping sickness antiparasitic
Table 2. African Trypanosomiasis treatment
| Species | Drug of choice | Adult Dosage | Pediatric Dosage |
|---|---|---|---|
| Trypanosoma brucei rhodesiense, hemolymphatic stage | Suramin1 | 1 gm IV on days 1, 3, 7 ,14, and 212 | 20 mg/kg IV on days 1, 3, 7, 14, and 213 |
| Trypanosoma brucei rhodesiense, CNS involvement | Melarsoprol4 | 2-3.6 mg/kg/day IV x 3 days.5 After 7 days, 3.6 mg/kg/day x 3 days. Give a 3rd series of 3.6 mg/kg/d after 7 days. | 2-3.6 mg/kg/day IV x 3 days.5 After 7 days, 3.6 mg/kg/day x 3 days. Give a 3rd series of 3.6 mg/kg/d after 7 days. |
| Trypanosoma brucei gambiense, Hemolymphatic stage | Pentamidine6 | 4 mg/kg/day IM or IV x 7-10 days | 4 mg/kg/day IM or IV x 7-10 days |
| Trypanosoma brucei gambiense, CNS involvement | Eflornithine7 | 400 mg/kg/day in 4 doses x 14 days | 400 mg/kg/day in 4 doses x 14 days |
Footnotes:
- Pentamidine is also effective against Trypanosoma brucei rhodesiense in the hemolymphatic stage, but suramin may have somewhat higher efficacy.
- A test dose of 100 mg should be given prior to the first dose and the patient should be monitored for hemodynamic stability.
- A test dose of 2 mg/kg should be given prior to the first dose and the patient should be monitored for hemodynamic stability.
- Corticosteroids have been used to prevent melarsoprol encephalopathy. The dose of melarsoprol is progressively increased during the first series.
- Suramin is also effective against Trypanosoma brucei gambiense in the hemolymphatic stage.
- Eflornithine (400 mg/kg/d IV in 2 doses x 7 days) given in combination with oral nifurtimox (15 mg/kg/d x 10 days) is also highly effective against
- Trypanosoma brucei gambiense with CNS involvement (Priotto G et al. Lancet 2009:374; 56-64). Nifurtimox is not FDA-approved for this indication.
Drugs used in the treatment of first stage
- Pentamidine: discovered in 1940, used for the treatment of the first stage of Trypanosoma brucei gambiense sleeping sickness. Despite non-negligible undesirable effects, it is in general well tolerated by patients.
- Suramin: discovered in 1920, used for the treatment of the first stage of Trypanosoma brucei rhodesiense. It provokes certain undesirable effects, including nephrotoxicity and allergic reactions.
Pentamidine, which is the recommended drug for first stage Trypanosoma brucei gambiense infection, is widely available in the U.S. The other drugs (suramin, melarsoprol, eflornithine, and nifurtimox) used to treat African trypanosomiasis are available in the U.S. only from the Centers for Disease Control and Prevention (CDC). Physicians can consult with CDC staff for advice on diagnosis and management and to obtain otherwise unavailable treatment drug.
Pentamidine, given by intravenous infusion over 2 hours or by intramuscular injection, is used to treat first stage Trypanosoma brucei gambiense infection. It is generally well tolerated, but adverse reactions of hypoglycemia, injection site pain, diarrhea, nausea and vomiting occur. Suramin is used to treat first stage Trypanosoma brucei rhodesiense. Suramin is also effective against Trypanosoma brucei gambiense, but it is not often used because severe reactions occur in persons who are co-infected with Onchocerca volvulus. Adverse reactions to suramin are frequent, but usually mild and reversible. These include drug rash, nephrotoxicity, and peripheral neuropathy. In rare instances, suramin administration results in a hypersensitivity reaction, and, for this reason, a small test dose is usually given prior to the full first dose.
Drugs used in the treatment of second stage
- Melarsoprol: discovered in 1949, it is used for the treatment of both gambiense and rhodesiense infections. It is derived from arsenic and has many undesirable side effects, the most dramatic of which is reactive encephalopathy (encephalopathic syndrome) which can be fatal (3% to 10%). It is currently recommended as first-line treatment for the rhodesiense form, but rarely used in the gambiense form.
- Eflornithine: much less toxic than melarsoprol, registered in 1990 is only effective against T.b. gambiense. It is generally used in combination with nifurtimox (as part of the Nifurtimox-eflornithine combination therapy, NECT) but can be used also as monotherapy. The regimen is complex and cumbersome to apply.
- Nifurtimox: The Nifurtimox-eflornithine combination therapy, NECT, was introduced in 2009. It simplifies the use of eflornithine by reducing the duration of treatment and the number of IV perfusions, but unfortunately it has not been studied for Trypanosoma brucei rhodesiense. Nifurtimox is registered for the treatment of American trypanosomiasis but not for human African trypanosomiasis. Both drugs are provided free of charge by WHO to endemic countries with a kit containing all the material needed for its administration.
For treatment advice and to obtain suramin, melarsoprol, or eflornithine, physicians should contact the Centers for Disease Control and Prevention (CDC) Division of Parasitic Diseases and Malaria.
Second stage Trypanosoma brucei gambiense is treated with eflornithine. Eflornithine is available for human use in the United States through the Centers for Disease Control and Prevention (CDC). Eflornithine is given in 4 intravenous infusions daily for 14 days. Adverse effects of eflornithine include bone marrow suppression, gastrointestinal symptoms, and seizures. Eflornithine is highly effective, but the difficulty in administering 4 infusions daily in rural African facilities has led to the use of eflornithine (dosed less frequently) in combination with nifurtimox. The efficacy of the combination regimen appears to be at least as high as eflornithine monotherapy. Eflornithine is not effective against Trypanosoma brucei rhodesiense and it is not recommended for treating the East African form of the disease. Melarsoprol, an organoarsenic compound, is the only drug available for treating second stage Trypanosoma brucei rhodesiense. Adverse reactions to melarsoprol can be severe and life-threatening. An encephalopathic reaction occurs in 5-10% of patients with a case-fatality rate of approximately 50% when it occurs. Prednisolone is often given to patients who are being treated with melarsoprol to reduce the risk of encephalopathy. Other adverse reactions observed with melarsoprol include skin reactions, gastrointestinal upset, and peripheral neuropathy. Intravenous injections of melarsoprol are painful and can cause phlebitis. The drug is administered by use of lengthy and complicated dosing schedules, however, an abbreviated 10-day regimen appears promising.
Eflornithine
- Note on treatment in pregnancy: Data on the use of eflornithine in pregnant women are limited, and risk to the embryo-fetus is unknown. Eflornithine should be used during pregnancy only if the potential benefit justifies the potential risk to the fetus.
- Note on treatment in breastfeeding: It is not known whether eflornithine is excreted in breast milk. Eflornithine should be used with caution in breast-feeding women.
- Note on treatment in children: The safety of eflornithine in children has not been established. Eflornithine is not approved by the Food and Drug Administration (FDA) for use in pediatric patients. Eflornithine is listed for the treatment of 1st stage African trypanosomiasis in Trypanosoma brucei gambiense infection on the WHO Model List of Essential Medicines for Children, intended for the use of children up to 12 years of age.
Pentamidine
- Note on treatment in pregnancy: Pentamidine is in pregnancy category C. Data on the use of pentamidine in pregnant women are limited, and risk to the embryo-fetus is unknown. Pentamidine should be used during pregnancy only if the potential benefit justifies the potential risk to the fetus.
- Note on treatment in breastfeeding: It is not known whether pentamidine is excreted in breast milk. The World Health Organization (WHO) classifies pentamidine as compatible with breast-feeding, although data on the use of pentamidine during lactation are limited. Pentamidine should be used during lactation only if the potential benefit of therapy to the mother justifies the potential risk to the infant.
- Note on treatment in children: Intravenous and intramuscular pentamidine have a similar safety profile in children age 4 months and older as in adults. Pentamidine is listed as a medicine for the treatment of 1st stage African trypanosomiasis infection (Trypanosoma brucei gambiense) on the WHO Model List of Essential Medicines for Children, intended for the use of children up to 12 years of age.
Drugs used in the treatment of both stages
Fexinidazole is an oral treatment for gambiense human African trypanosomiasis. It was included in 2019 in the WHO Essential medicines list and WHO human African Trypanosomiasis treatment guidelines. This molecule is indicated as first line for first stage and non-severe second stage. It should be administered within 30 minutes after a solid meal and under supervision of trained medical staff. Currently a clinical trial for its use in rhodesiense African sleeping sickness is ongoing.
African sleeping sickness prognosis
If the African sleeping sickness is treated during the early stage, recovery is possible in most patients 79. However, if the patient presents with stage 2 disease, the central nervous system (brain and spinal cord) involvement usually is fatal. Without treatment, death can occur within 6 months from cardiac failure or from Trypanosoma brucei rhodesiense infection itself.
Today, the cure rate with the drug melarsoprol is more than 90% 2.
- Sutherland CS, Yukich J, Goeree R, Tediosi F. A literature review of economic evaluations for a neglected tropical disease: human African trypanosomiasis (“sleeping sickness”). PLoS Negl Trop Dis. 2015 Feb;9(2):e0003397[↩][↩]
- Kazumba, L. M., Kaka, J. T., Ngoyi, D. M., & Tshala-Katumbay, D. (2018). Mortality trends and risk factors in advanced stage-2 Human African Trypanosomiasis: A critical appraisal of 23 years of experience in the Democratic Republic of Congo. PLoS neglected tropical diseases, 12(6), e0006504. https://doi.org/10.1371/journal.pntd.0006504[↩][↩]
- Simarro, P. P., Cecchi, G., Paone, M., Franco, J. R., Diarra, A., Ruiz, J. A., Fèvre, E. M., Courtin, F., Mattioli, R. C., & Jannin, J. G. (2010). The Atlas of human African trypanosomiasis: a contribution to global mapping of neglected tropical diseases. International journal of health geographics, 9, 57. https://doi.org/10.1186/1476-072X-9-57[↩]
- Hemmige V, Tanowitz H, Sethi A. Trypanosoma cruzi infection: a review with emphasis on cutaneous manifestations. Int. J. Dermatol. 2012 May;51(5):501-8.[↩]
- Trypanosomiasis, human African (sleeping sickness). https://www.who.int/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness[↩][↩][↩]
- Franco, J. R., Simarro, P. P., Diarra, A., & Jannin, J. G. (2014). Epidemiology of human African trypanosomiasis. Clinical epidemiology, 6, 257–275. https://doi.org/10.2147/CLEP.S39728[↩][↩][↩][↩]
- World Health Organization. Control and surveillance of human African trypanosomiasis. World Health Organ Tech Rep Ser. 2013;(984):1-237.[↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩][↩]
- Brun R, Blum J, Chappuis F, Burri C. Human African trypanosomiasis. Lancet. 2010 Jan 09;375(9709):148-59[↩][↩]
- Bentley SJ, Jamabo M, Boshoff A. The Hsp70/J-protein machinery of the African trypanosome, Trypanosoma brucei. Cell Stress Chaperones. 2019 Jan;24(1):125-148[↩]
- Kame-Ngasse, G. I., Njiokou, F., Melachio-Tanekou, T. T., Farikou, O., Simo, G., & Geiger, A. (2018). Prevalence of symbionts and trypanosome infections in tsetse flies of two villages of the “Faro and Déo” division of the Adamawa region of Cameroon. BMC microbiology, 18(Suppl 1), 159. https://doi.org/10.1186/s12866-018-1286-5[↩]
- Gao, J., Qian, Z., Hide, G., Lai, D., Lun, Z., & Wu, Z. (2020). Human African trypanosomiasis: The current situation in endemic regions and the risks for non-endemic regions from imported cases. Parasitology, 147(9), 922-931. doi:10.1017/S0031182020000645[↩][↩]
- Epidemiology and control of African trypanosomiasis. Report of a WHO Expert Committee. World Health Organ Tech Rep Ser. 1986;739:1-127.[↩]
- Uilenberg G. A Field Guide for the Diagnosis, Treatment and Prevention of African animal Trypanosomosis. Rome, Italy: Food and Agriculture Organization of the United Nations; 1998.[↩]
- Berberof M, Pérez-Morga D, Pays E. A receptor-like flagellar pocket glycoprotein specific to Trypanosoma brucei gambiense. Mol Biochem Parasitol. 2001 Mar;113(1):127-38. doi: 10.1016/s0166-6851(01)00208-0[↩]
- Rotureau, B., & Van Den Abbeele, J. (2013). Through the dark continent: African trypanosome development in the tsetse fly. Frontiers in cellular and infection microbiology, 3, 53. https://doi.org/10.3389/fcimb.2013.00053[↩][↩]
- Dyer NA, Rose C, Ejeh NO, Acosta-Serrano A. Flying tryps: survival and maturation of trypanosomes in tsetse flies. Trends Parasitol. 2013 Apr;29(4):188-96. doi: 10.1016/j.pt.2013.02.003[↩][↩][↩]
- MacGregor, P., & Matthews, K. R. (2010). New discoveries in the transmission biology of sleeping sickness parasites: applying the basics. Journal of molecular medicine (Berlin, Germany), 88(9), 865–871. https://doi.org/10.1007/s00109-010-0637-y[↩][↩][↩]
- Vickerman K, Tetley L, Hendry KA, Turner CM. Biology of African trypanosomes in the tsetse fly. Biol Cell. 1988;64(2):109-19. doi: 10.1016/0248-4900(88)90070-6[↩]
- Brun R, Blum J, Chappuis F, Burri C. Human African trypanosomiasis. Lancet. 2010 Jan 9;375(9709):148-59. doi: 10.1016/S0140-6736(09)60829-1[↩]
- Peacock, L., Ferris, V., Sharma, R., Sunter, J., Bailey, M., Carrington, M., & Gibson, W. (2011). Identification of the meiotic life cycle stage of Trypanosoma brucei in the tsetse fly. Proceedings of the National Academy of Sciences of the United States of America, 108(9), 3671–3676. https://doi.org/10.1073/pnas.1019423108[↩]
- Koffi M, Solano P, Barnabé C, de Meeûs T, Bucheton B, Cuny G, Jamonneau V. Genetic characterisation of Trypanosoma brucei s.l. using microsatellite typing: new perspectives for the molecular epidemiology of human African trypanosomiasis. Infect Genet Evol. 2007 Dec;7(6):675-84. doi: 10.1016/j.meegid.2007.07.001[↩]
- Pépin J, Méda HA. The epidemiology and control of human African trypanosomiasis. Adv Parasitol. 2001;49:71-132. doi: 10.1016/s0065-308x(01)49038-5[↩][↩][↩]
- MacGregor, P., Szöőr, B., Savill, N. J., & Matthews, K. R. (2012). Trypanosomal immune evasion, chronicity and transmission: an elegant balancing act. Nature reviews. Microbiology, 10(6), 431–438. https://doi.org/10.1038/nrmicro2779[↩]
- Pays E. The variant surface glycoprotein as a tool for adaptation in African trypanosomes. Microbes Infect. 2006 Mar;8(3):930-7. doi: 10.1016/j.micinf.2005.10.002[↩]
- Berriman M, Ghedin E, Hertz-Fowler C, Blandin G, Renauld H, Bartholomeu DC, Lennard NJ, Caler E, Hamlin NE, Haas B, Böhme U, Hannick L, Aslett MA, Shallom J, Marcello L, Hou L, Wickstead B, Alsmark UC, Arrowsmith C, Atkin RJ, Barron AJ, Bringaud F, Brooks K, Carrington M, Cherevach I, Chillingworth TJ, Churcher C, Clark LN, Corton CH, Cronin A, Davies RM, Doggett J, Djikeng A, Feldblyum T, Field MC, Fraser A, Goodhead I, Hance Z, Harper D, Harris BR, Hauser H, Hostetler J, Ivens A, Jagels K. et al, The genome of the African trypanosome Trypanosoma brucei. Science. 2005 Jul 15;309(5733):416-22. doi: 10.1126/science.1112642[↩]
- Fenn, K., & Matthews, K. R. (2007). The cell biology of Trypanosoma brucei differentiation. Current opinion in microbiology, 10(6), 539–546. https://doi.org/10.1016/j.mib.2007.09.014[↩]
- Control and surveillance of African trypanosomiasis. Report of a WHO Expert Committee. World Health Organ Tech Rep Ser. 1998;881:I-VI, 1-114.[↩]
- Moloo SK. The distribution of Glossina species in Africa and their natural hosts. Insect Science and its Application. 1993;14:511–527.[↩]
- Elsen P, Amoudi MA, Leclercq M. A propos de la découverte, en Arabie Saoudite, de deux espèces de mouches tsé-tsé vectrices de trypanosomiases humaines et animales [The discovery in Saudi Arabia of 2 species of tse-tse flies, vectors of human and animal trypanosomiasis]. Rev Med Liege. 1991 Apr;46(4):225-31. French.[↩]
- Molyneux DH. Host-trypanosomes interactions in Glossina. Insect Science and its Application. 1980;1:39–46.[↩]
- Maudlin I, Welburn SC. A single trypanosome is sufficient to infect a tsetse fly. Ann Trop Med Parasitol. 1989 Aug;83(4):431-3. doi: 10.1080/00034983.1989.11812368[↩]
- Thuita, J. K., Kagira, J. M., Mwangangi, D., Matovu, E., Turner, C. M., & Masiga, D. (2008). Trypanosoma brucei rhodesiense transmitted by a single tsetse fly bite in vervet monkeys as a model of human African trypanosomiasis. PLoS neglected tropical diseases, 2(5), e238. https://doi.org/10.1371/journal.pntd.0000238[↩]
- Aksoy S, Gibson WC, Lehane MJ. Interactions between tsetse and trypanosomes with implications for the control of trypanosomiasis. Adv Parasitol. 2003;53:1-83. doi: 10.1016/s0065-308x(03)53002-0[↩]
- Welburn SC, Maudlin I. The nature of the teneral state in Glossina and its role in the acquisition of trypanosome infection in tsetse. Ann Trop Med Parasitol. 1992 Oct;86(5):529-36. doi: 10.1080/00034983.1992.11812703. Erratum in: Ann Trop Med Parasitol. 2003 Nov;15(9):801.[↩]
- Tirados, I., Esterhuizen, J., Rayaisse, J. B., Diarrassouba, A., Kaba, D., Mpiana, S., Vale, G. A., Solano, P., Lehane, M. J., & Torr, S. J. (2011). How do tsetse recognise their hosts? The role of shape in the responses of tsetse (Glossina fuscipes and G. palpalis) to artificial hosts. PLoS neglected tropical diseases, 5(8), e1226. https://doi.org/10.1371/journal.pntd.0001226[↩]
- Rayaisse, J. B., Esterhuizen, J., Tirados, I., Kaba, D., Salou, E., Diarrassouba, A., Vale, G. A., Lehane, M. J., Torr, S. J., & Solano, P. (2011). Towards an optimal design of target for tsetse control: comparisons of novel targets for the control of Palpalis group tsetse in West Africa. PLoS neglected tropical diseases, 5(9), e1332. https://doi.org/10.1371/journal.pntd.0001332[↩]
- Saini RK, Hassanali A. A 4-alkyl-substituted analogue of guaiacol shows greater repellency to savannah tsetse (Glossina spp.). J Chem Ecol. 2007 May;33(5):985-95. doi: 10.1007/s10886-007-9272-7[↩]
- Torr, S. J., Chamisa, A., Mangwiro, T. N., & Vale, G. A. (2012). Where, when and why do tsetse contact humans? Answers from studies in a national park of Zimbabwe. PLoS neglected tropical diseases, 6(8), e1791. https://doi.org/10.1371/journal.pntd.0001791[↩]
- Rayaisse, J. B., Tirados, I., Kaba, D., Dewhirst, S. Y., Logan, J. G., Diarrassouba, A., Salou, E., Omolo, M. O., Solano, P., Lehane, M. J., Pickett, J. A., Vale, G. A., Torr, S. J., & Esterhuizen, J. (2010). Prospects for the development of odour baits to control the tsetse flies Glossina tachinoides and G. palpalis s.l. PLoS neglected tropical diseases, 4(3), e632. https://doi.org/10.1371/journal.pntd.0000632[↩]
- Sané B, Laveissière C, Meda HA. Diversité du régime alimentaire de Glossina palpalis palpalis en zone forestière de Côte d’Ivoire: relation avec la prévalence de la trypanosomiase humaine africaine [Diversity of feeding behavior of Glossina palpalis palpalis in the forest belt of the Ivory Coast: relation to the prevalence of human African trypanosomiasis]. Trop Med Int Health. 2000 Jan;5(1):73-8. French. doi: 10.1046/j.1365-3156.2000.00488.x[↩]
- Torr SJ, Maudlin I, Vale GA. Less is more: restricted application of insecticide to cattle to improve the cost and efficacy of tsetse control. Med Vet Entomol. 2007 Mar;21(1):53-64. doi: 10.1111/j.1365-2915.2006.00657.x[↩]
- Challier A. The ecology of tsetse (Glossina spp.) (Diptera, Glossinidae): a review (1970–1981) Insect Science and its Application. 1982;3(2–3):97–143.[↩]
- Baker RD. Modelling the probability of a single trypanosome infecting a tsetse fly. Ann Trop Med Parasitol. 1991 Aug;85(4):413-5. doi: 10.1080/00034983.1991.11812585[↩]
- Funk, S., Nishiura, H., Heesterbeek, H., Edmunds, W. J., & Checchi, F. (2013). Identifying transmission cycles at the human-animal interface: the role of animal reservoirs in maintaining gambiense human african trypanosomiasis. PLoS computational biology, 9(1), e1002855. https://doi.org/10.1371/journal.pcbi.1002855[↩]
- Woodruff AW, Evans DA, Owino NO. A healthy carrier of African trypanosomiasis. J Infect. 1982;5:89–92.[↩]
- Jamonneau, V., Bucheton, B., Kaboré, J., Ilboudo, H., Camara, O., Courtin, F., Solano, P., Kaba, D., Kambire, R., Lingue, K., Camara, M., Baelmans, R., Lejon, V., & Büscher, P. (2010). Revisiting the immune trypanolysis test to optimise epidemiological surveillance and control of sleeping sickness in West Africa. PLoS neglected tropical diseases, 4(12), e917. https://doi.org/10.1371/journal.pntd.0000917[↩]
- Checchi, F., Filipe, J. A., Barrett, M. P., & Chandramohan, D. (2008). The natural progression of Gambiense sleeping sickness: what is the evidence?. PLoS neglected tropical diseases, 2(12), e303. https://doi.org/10.1371/journal.pntd.0000303[↩]
- Naessens J. Bovine trypanotolerance: A natural ability to prevent severe anaemia and haemophagocytic syndrome? Int J Parasitol. 2006 May 1;36(5):521-8. doi: 10.1016/j.ijpara.2006.02.012[↩]
- Njiokou F, Nimpaye H, Simo G, Njitchouang GR, Asonganyi T, Cuny G, Herder S. Domestic animals as potential reservoir hosts of Trypanosoma brucei gambiense in sleeping sickness foci in Cameroon. Parasite. 2010 Mar;17(1):61-6. doi: 10.1051/parasite/2010171061[↩]
- Balyeidhusa AS, Kironde FA, Enyaru JC. Apparent lack of a domestic animal reservoir in Gambiense sleeping sickness in northwest Uganda. Vet Parasitol. 2012 Jun 8;187(1-2):157-67. doi: 10.1016/j.vetpar.2011.12.005[↩]
- Jamonneau V, Ravel S, Koffi M, Kaba D, Zeze DG, Ndri L, Sane B, Coulibaly B, Cuny G, Solano P. Mixed infections of trypanosomes in tsetse and pigs and their epidemiological significance in a sleeping sickness focus of Côte d’Ivoire. Parasitology. 2004 Dec;129(Pt 6):693-702. doi: 10.1017/s0031182004005876[↩]
- Cordon-Obras C, Berzosa P, Ndong-Mabale N, Bobuakasi L, Buatiche JN, Ndongo-Asumu P, Benito A, Cano J. Trypanosoma brucei gambiense in domestic livestock of Kogo and Mbini foci (Equatorial Guinea). Trop Med Int Health. 2009 May;14(5):535-41. doi: 10.1111/j.1365-3156.2009.02271.x[↩]
- Njiokou F, Laveissière C, Simo G, Nkinin S, Grébaut P, Cuny G, Herder S. Wild fauna as a probable animal reservoir for Trypanosoma brucei gambiense in Cameroon. Infect Genet Evol. 2006 Mar;6(2):147-53. doi: 10.1016/j.meegid.2005.04.003[↩]
- Schütt ID, Mehlitz D. On the persistence of human serum resistance and isoenzyme patterns of Trypanozoon in experimentally infected pigs. Acta Trop. 1981 Dec;38(4):367-73.[↩]
- Ruiz-Postigo, J. A., Franco, J. R., Lado, M., & Simarro, P. P. (2012). Human African trypanosomiasis in South Sudan: how can we prevent a new epidemic?. PLoS neglected tropical diseases, 6(5), e1541. https://doi.org/10.1371/journal.pntd.0001541[↩]
- Simarro PP, Franco JR, Ndongo P, Nguema E, Louis FJ, Jannin J. The elimination of Trypanosoma brucei gambiense sleeping sickness in the focus of Luba, Bioko Island, Equatorial Guinea. Trop Med Int Health. 2006 May;11(5):636-46. doi: 10.1111/j.1365-3156.2006.01624.x[↩]
- Dje NN, Miezan TW, N’guessan P, Brika P, Doua F, Boa F. Distribution géographique des trypanosomés pris en charge en Côte d’Ivoire de 1993 à 2000 [Geographic distribution of trypanosomiasis treated in Ivory Coast from 1993 to 2000]. Bull Soc Pathol Exot. 2002 Dec;95(5):359-61. French.[↩]
- Checchi, F., Cox, A. P., Chappuis, F., Priotto, G., Chandramohan, D., & Haydon, D. T. (2012). Prevalence and under-detection of gambiense human African trypanosomiasis during mass screening sessions in Uganda and Sudan. Parasites & vectors, 5, 157. https://doi.org/10.1186/1756-3305-5-157[↩]
- von Wissmann, B., Machila, N., Picozzi, K., Fèvre, E. M., deC Bronsvoort, B. M., Handel, I. G., & Welburn, S. C. (2011). Factors associated with acquisition of human infective and animal infective trypanosome infections in domestic livestock in Western Kenya. PLoS neglected tropical diseases, 5(1), e941. https://doi.org/10.1371/journal.pntd.0000941[↩][↩]
- Njagu Z, Mihok S, Kokwaro E, Verloo D. Isolation of Trypanosoma brucei from the monitor lizard (Varanus niloticus) in an endemic focus of Rhodesian sleeping sickness in Kenya. Acta Trop. 1999 Mar 15;72(2):137-48. doi: 10.1016/s0001-706x(98)00092-8[↩]
- Migchelsen SJ, Büscher P, Hoepelman AI, Schallig HD, Adams ER. Human African trypanosomiasis: a review of non-endemic cases in the past 20 years. Int J Infect Dis. 2011 Aug;15(8):e517-24. doi: 10.1016/j.ijid.2011.03.018[↩]
- Welburn SC, Picozzi K, Fèvre EM, Coleman PG, Odiit M, Carrington M, Maudlin I. Identification of human-infective trypanosomes in animal reservoir of sleeping sickness in Uganda by means of serum-resistance-associated (SRA) gene. Lancet. 2001 Dec 15;358(9298):2017-9. doi: 10.1016/s0140-6736(01)07096-9[↩]
- Grebaut P, Bodo JM, Assona A, Foumane Ngane V, Njiokou F, Ollivier G, Soula G, Laveissiere C. Recherche des facteurs de risque de la trypanosomose humaine africaine dans le foyer de Bipindi au Cameroun [Risk factors for human African trypanosomiasis in the Bipindi region of Cameroon]. Med Trop (Mars). 2001;61(4-5):377-83. French.[↩]
- Kohagne, T. L., M’eyi, M. P., Kamkuimo, R. G., Kaba, D., Louis, J. F., & Mimpfoundi, R. (2011). Transmission of human African trypanosomiasis in the Komo-Mondah focus, Gabon. The Pan African medical journal, 8, 36. https://doi.org/10.4314/pamj.v8i1.71151[↩]
- Moore A, Richer M, Enrile M, Losio E, Roberts J, Levy D. Resurgence of sleeping sickness in Tambura County, Sudan. Am J Trop Med Hyg. 1999 Aug;61(2):315-8. doi: 10.4269/ajtmh.1999.61.315[↩]
- Courtin F, Jamonneau V, Camara M, Camara O, Coulibaly B, Diarra A, Solano P, Bucheton B. A geographical approach to identify sleeping sickness risk factors in a mangrove ecosystem. Trop Med Int Health. 2010 Aug;15(8):881-9. doi: 10.1111/j.1365-3156.2010.02559.x[↩]
- Meda AH, Laveissiere C, De Muynck A, Doua F, Diallo PB. Les facteurs de risque de la trypanosomiase humaine africaine dans les foyers endemiques de Cote d’Ivoire [Risk factors for human African trypanosomiasis in the endemic foci of Ivory Coast]. Med Trop (Mars). 1993 Jan-Mar;53(1):83-92. French.[↩]
- Grébaut P, Bena JM, Manzambi EZ, Mansinsa P, Khande V, Ollivier G, Cuny G, Simo G. Characterization of sleeping sickness transmission sites in rural and periurban areas of Kinshasa (République Démocratique du Congo). Vector Borne Zoonotic Dis. 2009 Dec;9(6):631-6. doi: 10.1089/vbz.2008.0118[↩]
- Fischer A, Simarro PP, Franco JR, Becher H, Steverding D. Increased trypanolytic activity in sera of sleeping sickness patients after chemotherapy. Trop Med Int Health. 2001 Dec;6(12):1070-4. doi: 10.1046/j.1365-3156.2001.00805.x[↩]
- Paquet C, Castilla J, Mbulamberi D, Beaulieu MF, Gastellu Etchegorry MG, Moren A. La trypanosomiase a Trypanosoma brucei gambiense dans le foyer du nord-ouest de l’Ouganda. Bilan de 5 années de lutte (1987-1991) [Trypanosomiasis from Trypanosoma brucei gambiense in the center of north-west Uganda. Evaluation of 5 years of control (1987-1991)]. Bull Soc Pathol Exot. 1995;88(1):38-41. French.[↩]
- Abel PM, Kiala G, Lôa V, Behrend M, Musolf J, Fleischmann H, Théophile J, Krishna S, Stich A. Retaking sleeping sickness control in Angola. Trop Med Int Health. 2004 Jan;9(1):141-8. doi: 10.1046/j.1365-3156.2003.01152.x. Erratum in: Trop Med Int Health. 2004 Feb;9(2):314.[↩]
- Khonde N, Pépin J, Niyonsenga T, De Wals P. Familial aggregation of Trypanosoma brucei gambiense trypanosomiasis in a very high incidence community in Zaire. Trans R Soc Trop Med Hyg. 1997 Sep-Oct;91(5):521-4. doi: 10.1016/s0035-9203(97)90008-0[↩]
- Simarro PP, Franco JR, Cecchi G, Paone M, Diarra A, Ruiz Postigo JA, Jannin JG. Human African trypanosomiasis in non-endemic countries (2000-2010). J Travel Med. 2012 Jan-Feb;19(1):44-53. doi: 10.1111/j.1708-8305.2011.00576.x. Epub 2011 Dec 8. Erratum in: J Travel Med. 2012 Mar-Apr;19(2):135.[↩][↩]
- Kinung’hi SM, Malele II, Kibona SN, et al. Knowledge, attitudes and practices on tsetse and sleeping sickness among communities living in and around Serengeti National Park, Tanzania. Tanzan Health Res Bull. 2006;8(3):168–172.[↩]
- Zoller, T., Fèvre, E. M., Welburn, S. C., Odiit, M., & Coleman, P. G. (2008). Analysis of risk factors for T. brucei rhodesiense sleeping sickness within villages in south-east Uganda. BMC infectious diseases, 8, 88. https://doi.org/10.1186/1471-2334-8-88[↩][↩][↩][↩][↩]
- Allsopp R. (1972). The role of game animals in the maintenance of endemic and enzootic trypanosomiases in the Lambwe Valley, South Nyanza District, Kenya. Bulletin of the World Health Organization, 47(6), 735–746.[↩]
- Vale, G. A., Chamisa, A., Mangwiro, C., & Torr, S. J. (2013). A neglected aspect of the epidemiology of sleeping sickness: the propensity of the tsetse fly vector to enter houses. PLoS neglected tropical diseases, 7(2), e2086. https://doi.org/10.1371/journal.pntd.0002086[↩]
- Jelinek, T., Bisoffi, Z., Bonazzi, L., van Thiel, P., Bronner, U., de Frey, A., Gundersen, S. G., McWhinney, P., Ripamonti, D., & European Network on Imported Infectious Disease Surveillance (2002). Cluster of African trypanosomiasis in travelers to Tanzanian national parks. Emerging infectious diseases, 8(6), 634–635. https://doi.org/10.3201/eid0806.010432[↩]
- Dunn N, Wang S, Adigun R. African Trypanosomiasis. [Updated 2021 Aug 11]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2021 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK519580[↩][↩]
- Ferrins, L., Sharma, A., Thomas, S. M., Mehta, N., Erath, J., Tanghe, S., Leed, S. E., Rodriguez, A., Mensa-Wilmot, K., Sciotti, R. J., Gillingwater, K., & Pollastri, M. P. (2018). Anilinoquinoline based inhibitors of trypanosomatid proliferation. PLoS neglected tropical diseases, 12(11), e0006834. https://doi.org/10.1371/journal.pntd.0006834[↩]
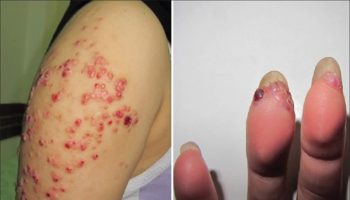

{kind=link}