
Contents
- Best Supplements for Muscle Building
- Protein sources
- Animal Protein
- Adjunctive Nutritional Strategies to Augment Muscle Protein Synthesis
- Protein Recommended intake
- Protein timing
- Pre-sleep protein intake
- Timing and consumption of protein and/or carbohydrate during workouts
- Micronutrients
- Caffeine
- Beta-alanine
- Beta-hydroxy-beta-methylbutyrate
- Creatine
- Creatine supplementation Responders vs. Non-responders
- Effects of creatine supplementation on Muscle Mass
- Effects of creatine ingestion to improve recovery from injury, muscle damage and oxidative stress induced by exercise
- Effects of creatine supplementation on glycogen stores
- Effects of creatine supplementation on predominantly aerobic exercise
- Documented effects of creatine supplementation on physical performance
- Effects of creatine supplementation on predominantly anaerobic exercise
- Effects of creatine supplementation on predominantly aerobic exercise
- Effects of creatine supplementation on range of motion
- Creatine in combination with other supplements
Best Supplements for Muscle Building
The International Society of Sports Nutrition 1 provides an objective and critical review related to the intake of protein for healthy, exercising individuals. Based on the current available literature, the position of the The International Society of Sports Nutrition is as follows:
- An acute exercise stimulus, particularly resistance exercise, and protein ingestion both stimulate muscle protein synthesis and are synergistic when protein consumption occurs before or after resistance exercise.
- For building muscle mass and for maintaining muscle mass through a positive muscle protein balance, an overall daily protein intake in the range of 1.4–2.0 g protein/kg body weight/day (g/kg/d) is sufficient for most exercising individuals, a value that falls in line within the Acceptable Macronutrient Distribution Range published by the Institute of Medicine for protein.
- Higher protein intakes (2.3–3.1 g/kg/d) may be needed to maximize the retention of lean body mass in resistance-trained subjects during hypocaloric periods.
- There is novel evidence that suggests higher protein intakes (>3.0 g/kg/d) may have positive effects on body composition in resistance-trained individuals (i.e., promote loss of fat mass).
- Recommendations regarding the optimal protein intake per serving for athletes to maximize muscle protein synthesis are mixed and are dependent upon age and recent resistance exercise stimuli. General recommendations are 0.25 g of a high-quality protein per kg of body weight, or an absolute dose of 20–40 g.
- Acute protein doses should strive to contain 700–3000 mg of leucine and/or a higher relative leucine content, in addition to a balanced array of the essential amino acids (EAAs).
- These protein doses should ideally be evenly distributed, every 3–4 hours, across the day.
- The optimal time period during which to ingest protein is likely a matter of individual tolerance, since benefits are derived from pre- or post-workout ingestion; however, the anabolic effect of exercise is long-lasting (at least 24 h), but likely diminishes with increasing time post-exercise.
- While it is possible for physically active individuals to obtain their daily protein requirements through the consumption of whole foods, supplementation is a practical way of ensuring intake of adequate protein quality and quantity, while minimizing caloric intake, particularly for athletes who typically complete high volumes of training.
- Rapidly digested proteins that contain high proportions of essential amino acids (EAAs) and adequate leucine, are most effective in stimulating muscle protein synthesis.
- Different types and quality of protein can affect amino acid bioavailability following protein supplementation.
- Athletes should consider focusing on whole food sources of protein that contain all of the EAAs (i.e., it is the EAAs that are required to stimulate muscle protein synthesis).
- Endurance athletes should focus on achieving adequate carbohydrate intake to promote optimal performance; the addition of protein may help to offset muscle damage and promote recovery.
- Pre-sleep casein protein intake (30–40 g) provides increases in overnight muscle protein synthesis and metabolic rate without influencing lipolysis.
Body composition
Improving your body composition through the loss of fat mass and increasing fat-free mass (lean mass) is often associated with improvements in physical performance. In this respect, many published investigations report that protein supplementation results in significant improvements in lean body weight/cross-sectional areas as compared to placebo treatments 2, 3, 4, 5, 6. Andersen et al. 2 examined 22 healthy men that completed a 14-week resistance-training program (3 days/week consisting of 3–4 sets of lower body exercises) while supplementing with either 25 g of a high-quality protein blend or 25 g of carbohydrate. When the blend of milk proteins was provided, significantly greater increases in lean mass, muscle cross-sectional area in both the Type I and Type II muscle fibers occurred when compared to changes seen with carbohydrate consumption. Collectively, a meta-analysis by Cermak and colleagues 7 reported a mean increase in fat-free mass of 0.69 kg when protein supplementation was provided versus a placebo during a resistance-training program. Other reviews by Tipton 8, Phillips 9 and Pasiakos 10, respectively, provide further support that protein supplementation (15–25 g over 4–14 weeks) augments lean mass accretion when combined with completion of a resistance training program.
Beyond gaining of fat-free mass, increasing daily protein intake through a combination of food and supplementation to levels above the recommended daily allowance (RDA) (RDA 0.8 g/kg/day, increasing to 1.2–2.4 g/kg/day for the endurance and strength/power athletes) while restricting energy intake (30–40% reduction in energy intake) has been demonstrated to maximize the loss of fat tissue while also promoting the maintenance of fat-free mass 11, 12. The majority of this work has been conducted using overweight and obese individuals who were prescribed an energy-restricted diet that delivered a greater ratio of protein relative to carbohydrate. As a classic example, Layman and investigators 11 randomized obese women to consume one of two restricted energy diets (1600–1700 kcals/day) that were either higher in carbohydrates (>3.5: carbohydrate-to-protein ratio) or protein (<1.5: carbohydrate-to-protein ratio). Groups were further divided into those that followed a five-day per week exercise program (walking + resistance training, 20–50 min/workout) and a control group that performed light walking of less than 100 min per week. Greater amounts of fat were lost when higher amounts of protein were ingested, but even greater amounts of fat loss occurred when the exercise program was added to the high-protein diet group, resulting in significant decreases in body fat. Using an active population that ranged from normal weight to overweight (BMI: 22–29 kg/m2), Pasiakos and colleagues 13 examined the impact of progressively increasing dietary protein over a 21-day study period. An aggressive energy reduction model was employed that resulted in each participant reducing their caloric intake by 30% and increasing their energy expenditure by 10%. Each person was randomly assigned to consume a diet that contained either 1× (0.8 g/kg), 2× (1.6 g/kg) or 3× (2.4 g/kg) the recommended daily allowance (RDA) for protein. Participants were measured for changes in body weight and body composition. While the greatest body weight loss occurred in the 1× RDA group, this group also lost the highest percentage of lean mass and lowest percentage of fat mass. The 2× and 3× RDA groups lost significant amounts of body weight that consisted of 70% and 64% fat mass, respectively.
Collectively, these results indicate that increasing dietary protein can promote favorable adaptations in body composition through the promotion of fat-free mass accretion when combined with a high calorie diet and a heavy resistance training program and can also promote the loss of fat mass when higher intakes of daily protein (2-3× the RDA) are combined with an exercise program and a low calorie diet.
Key points:
- When combined with a high calorie diet and a heavy resistance-training program, protein supplementation may promote increases in skeletal muscle cross-sectional area and lean body mass.
- When combined with a resistance-training program and a low calorie diet, an elevated daily intake of protein (2 – 3× the RDA) can promote greater losses of fat mass and greater overall improvements in body composition.
Resistance exercise performance
The extent to which protein supplementation, in conjunction with resistance training, enhances maximal strength is contingent upon many factors, including:
- Resistance-training program variables (such as intensity, volume, and progression)
- Length of the resistance-training program/intervention
- Training status of the participants engaging in the resistance-training program
- Energy intake in the diet
- Quality and quantity of protein intake (with an emphasis on leucine content of the protein)
- Co-ingestion of additional dietary ingredients that may favorably impact strength (e.g. creatine, Beta-hydroxy-beta-methylbutyrate (HMB))
Taking each of these variables into consideration, the effects of supplemental protein consumption has on maximal strength enhancement are varied, with a majority of the investigations reporting no benefit 14, 15, 16 and a few reporting improvements in maximal strength 17, 18. With limited exceptions, most of the studies utilized young, healthy, untrained males as participants. In one investigation examining college football athletes supplementing with a proprietary milk protein supplement (two servings of 42 g per day) for 12 weeks, a 14.5% increase in maximal squat strength was observed compared to a 6.9% increase in the placebo group 19. These differences were statistically significant. When females were the only sex investigated, the outcomes consistently indicated that supplemental protein does not appear to enhance maximal strength at magnitudes that reach statistical significance. Hida et al. 20 reported that females supplementing with 15 g of egg white protein (which raised daily protein intake to 1.23 g of protein/kg body weight/day) experienced no improvements in maximal upper and lower body strength as compared to a carbohydrate placebo (ingesting one gram of protein/kg body weight/day) over an 8-week period. An important note for this study is that 15 g of egg protein is considered by many to be a sub-optimal dose 21. However, others have advocated that the total daily intake of protein might be as important or more important 22. In another study, Josse et al. 23 reported that non-resistance trained females supplementing with one liter of skimmed bovine milk (providing 36 g of protein) after resistance exercise improved maximal strength in seven of nine measures as compared to a carbohydrate placebo group, but only the improvements to maximal bench press strength attained statistical significance compared to the placebo. In contrast, Taylor and colleagues 24 reported that pre- and post-exercise whey protein ingestion significantly increased maximal upper-body strength (+4.9 kg bench press one repetition maximum) in comparison to changes seen when a maltodextrin placebo (+2.3 kg) was ingested in a group of female collegiate basketball players over an 8-week period.
In summary, while research investigating the addition of supplemental protein to a diet with adequate energy and nutrient intakes is inconclusive in regards to stimulating strength gains in conjunction with a resistance-training program to a statistically significant degree, greater protein intakes that are achieved from both dietary and supplemental sources do appear to have some advantage. Hoffman and colleagues 18 reported that in athletes consuming daily protein intakes above 2.0 g/kg/d which included protein intakes from both diet and supplements, a 22% and 42% increase in strength was noted in both the squat and bench press exercises during off-season conditioning in college football players compared to athletes that consumed only the recommended levels (1.6–1.8 g/kg/d) for strength/power athletes. Further, it is important to highlight that in most studies cited, protein intervention resulted in greater but non-statistically significant strength improvements as compared to the placebo/control condition. Cermak and colleagues 25 pooled the outcomes from 22 separate clinical trials to yield 680 subjects in their statistical analysis and found that protein supplementation with resistance training resulted in a 13.5 kg increase in lower-body strength when compared to changes seen when a placebo was provided. A similar conclusion was also drawn by Pasiakos et al. 26 in a meta-analysis where they reported that in untrained participants, protein supplementation might exert very little benefit on strength during the initial weeks of a resistance training program, but as duration, frequency and volume of resistance training increased, protein supplementation may favorably impact skeletal muscle hypertrophy and strength.
Key points:
- Results from many single investigations indicate that in both men and women protein supplementation exerts a small to modest impact on strength development.
- Pooled results of multiple studies using meta-analytic and other systematic approaches consistently indicate that protein supplementation (15 to 25 g over 4 to 21 weeks) exerts a positive impact on performance.
Regulation of Muscle Protein Turnover
The process of skeletal muscle protein turnover is constant and ongoing. Protein turnover within muscle is the sum of the processes of both muscle protein synthesis (MPS) and muscle protein breakdown (MPB). Beyond childhood growth, chronic imbalances between the processes of MPS and MPB lead to a net gain in protein pool size (hypertrophy: MPS > MPB) or a net loss (atrophy: MPB > MPS). With regular practice, resistance exercise can lead to gains in skeletal muscle mass by means of hypertrophy. The process of skeletal muscle fiber hypertrophy comes about as a result of the confluence of positive muscle protein balance and satellite cell addition to muscle fibers. Positive muscle protein balance is achieved when the rate of new muscle protein synthesis (MPS) exceeds that of muscle protein breakdown (MPB). While resistance exercise and postprandial hyperaminoacidemia both stimulate muscle protein synthesis, it is through the synergistic effects of these two stimuli that a net gain in muscle proteins occurs and muscle fiber hypertrophy takes place 27.
Resistance exercise provides a loading stimulus to skeletal muscle that results in increases in skeletal muscle protein synthesis and, if performed in the fasted state, an increase in muscle protein breakdown 28. The increase in fasted-stated muscle protein synthesis with resistance exercise is long-lasting and persists for at least 48 h 28, and maybe longer with a higher volume of focused contractions 29. Provision of amino acids intravenously 30, as isolated proteins 31, or in foods such as beef and milk that promote hyperaminoacidemia and hyperinsulinemia are all effective in stimulating muscle protein synthesis. In addition, post-exercise hyperaminoacidemia suppresses the rise in muscle protein breakdown 30 that occurs following resistance exercise in the fasted state 32. Post-exercise hyperinsulinemia is not overtly stimulatory for muscle protein synthesis 33, but will also simultaneously suppress muscle protein breakdown 33. It thus appears that rather than being strictly anabolic, the hyperinsulinemia that accompanies post-exercise protein consumption is not stimulatory but probably merely permissive for muscle protein synthesis 34 and suppressive for muscle protein breakdown 35. Therefore, when protein is ingested after resistance exercise it is the amino acids themselves that are driving the rise in post-exercise muscle protein synthesis 36. It is also now quite clear that it is really only the essential amino acids (EAA) that drive the process of muscle protein synthesis 37. However, perhaps more important is that the key essential amino acid is leucine, as it alone appears to be the metabolic trigger for muscle protein synthesis 38, 39.
Dose–Response of Dietary Protein and Muscle Protein Synthesis
To date only three true dose–response studies in which muscle protein synthesis has been measured have been published 36, 40, 41. In those studies, the main message was that muscle protein synthesis is a saturable process in young people at protein ingestion doses of approximately 20–25 g (~8.5–10 g of essential amino acids) regardless of whether the subjects exercised 36 or not 41. Moore et al. 36 also noted that, in parallel with the rise in muscle protein synthesis, the albumin protein synthetic rate showed a strikingly similar saturable dose–response curve, demonstrating that at least one other body protein had similar synthetic kinetics. In an attempt to standardize this protein dose to body mass (BM), and using the subjects’ mass from the study by Moore et al. 36, the dose of protein that was maximally effective (20 g) post-exercise equated to approximately 0.25 g protein/kg BM. While egg was the protein source used in that study 36, the rationale being that it is the internationally recognized standard protein, similar data would be expected with other high-quality proteins. However, the dose of protein that is maximally stimulatory in older adults is closer to 40 g following resistance exercise and 20 g at rest 41. Beyond the levels at which muscle protein synthesis is maximally stimulated, it has been noted that the oxidation of an indicator amino acid, leucine, rises quite sharply 41, indicating that amino acids are not being used for protein synthesis and instead are oxidized, probably leading to urea production. While oxidative amino acid loss has been used as an indication of protein excess, it may well be that oxidative losses would still occur despite the fact that protein synthesis is not maximally stimulated as a result of lower Km (Michaelis–Menten kinetics—the substrate concentration at which the reaction rate is half of maximal) values of enzymes involved in amino acid degradation. Metabolic products of amino acids (urea, creatinine, uric acid, and other nitrogenous products) are excreted in the urine; nitrogen is also lost in feces, sweat, and other body secretions and in sloughed skin, hair, and nails. A continuous supply of dietary amino acids is required to replace these losses, even after growth has ceased 42. The traditional interpretation of amino acid oxidation as being ‘wasteful’ may not be a true sentiment where optimal stimulation of muscle protein synthesis is concerned.
Protein deficiency rarely occurs as an isolated condition. It usually accompanies a deficiency of dietary energy and other nutrients resulting from insufficient food intake. The symptoms are most commonly seen in deprived children in poor countries. Where protein intake is exceptionally low, there are physical signs—stunting, poor musculature, edema, thin and fragile hair, skin lesions—and biochemical changes that include low serum albumin and hormonal imbalances. Edema and loss of muscle mass and hair are the prominent signs in adults. Deficiency of this severity is very rare in the United States, except as a consequence of pathologic conditions and poor medical management of the acutely ill.
Protein quality
There are 20 total amino acids, comprised of 9 EAAs (essential amino acids) and 11 non-essential amino acids (NEAAs). Nine essential amino acids (EAAs)—histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, and valine — cannot be produced in the body and therefore must be consumed in the diet 1. Several methods exist to determine protein quality such as Chemical Score, Protein Efficiency Ratio, Biological Value, Protein Digestibility-Corrected Amino Acid Score (PDCAAS) and most recently, the Indicator Amino Acid Oxidation (IAAO) technique. Ultimately, in vivo protein quality is typically defined as how effective a protein is at stimulating muscle protein synthesis (MPS) and promoting muscle hypertrophy 43. Overall, research has shown that products containing animal and dairy-based proteins contain the highest percentage of EAAs and result in greater hypertrophy and protein synthesis following resistance training when compared to a vegetarian protein-matched control, which typically lacks one or more EAAs 44.
Several studies, but not all, 45 have indicated that EAAs alone stimulate protein synthesis in the same magnitude as a whole protein with the same EAA content 46. For example, Borsheim et al. 47 found that 6 g of EAAs stimulated protein synthesis twice as much as a mixture of 3 g of NEAAs combined with 3 g of EAAs. Moreover, Paddon-Jones and colleagues 48 found that a 180-cal supplement containing 15 g of EAAs stimulated greater rates of protein synthesis than an 850-cal meal with the same EAA content from a whole protein source. While important, the impact of a larger meal on changes in circulation and the subsequent delivery of the relevant amino acids to the muscle might operate as important considerations when interpreting this data. In contrast, Katsanos and colleagues 45 had 15 elderly subjects consume either 15 g of whey protein or individual doses of the essential and nonessential amino acids that were identical to what is found in a 15-g whey protein dose on separate occasions. Whey protein ingestion significantly increased leg phenylalanine balance, an index of muscle protein accrual, while EAA and NEAA ingestion exerted no significant impact on leg phenylalanine balance. This study, and the results reported by others 49 have led to the suggestion that an approximate 10 g dose of EAAs might serve as an optimal dose to maximally stimulate MPS and that intact protein feedings of appropriate amounts (as opposed to free amino acids) to elderly individuals may stimulate greater improvements in leg muscle protein accrual.
Based on this research, scientists have also attempted to determine which of the EAAs are primarily responsible for modulating protein balance. The three branched-chain amino acids (BCAAs), leucine, isoleucine, and valine are unique among the EAAs for their roles in protein metabolism 50, neural function 51, and blood glucose and insulin regulation 52. Additionally, enzymes responsible for the degradation of BCAAs operate in a rate-limiting fashion and are found in low levels in splanchnic tissues. Thus, orally ingested BCAAs appear rapidly in the bloodstream and expose muscle to high concentrations ultimately making them key components of skeletal MPS 53. Furthermore, Wilson and colleagues 54 have recently demonstrated, in an animal model, that leucine ingestion (alone and with carbohydrate) consumed between meals (135 min post-consumption) extends protein synthesis by increasing the energy status of the muscle fiber. Multiple human studies have supported the contention that leucine drives protein synthesis. Moreover, this response may occur in a dose-dependent fashion, plateauing at approximately 2 g at rest 54, and increasing up to 3.5 g when ingestion occurs after completion of a 60-min bout of moderate intensity cycling 55. However, it is important to realize that the duration of protein synthesis after resistance exercise appears to be limited by both the signal (leucine concentrations), ATP status, as well as the availability of substrate (i.e., additional EAAs found in a whole protein source) 56. As such, increasing leucine concentration may stimulate increases in muscle protein, but a higher total dose of all EAAs (as free form amino acids or intact protein sources) seems to be most suited for sustaining the increased rates of MPS 56.
Table 1. Protein quality rankings
| Protein Type | Protein Efficiency Ratio | Biological Value | Net Protein Utilization | Protein Digestibility Corrected Amino Acid Score (PDCAAS) |
|---|---|---|---|---|
| Beef | 2.9 | 80 | 73 | 0.92 |
| Black Beans | 0 | 0 | 0.75 | |
| Casein | 2.5 | 77 | 76 | 1.00 |
| Egg | 3.9 | 100 | 94 | 1.00 |
| Milk | 2.5 | 91 | 82 | 1.00 |
| Peanuts | 1.8 | 0.52 | ||
| Soy protein | 2.2 | 74 | 61 | 1.00 |
| Wheat gluten | 0.8 | 64 | 67 | 0.25 |
| Whey protein | 3.2 | 104 | 92 | 1.00 |
It is well known that exercise improves net muscle protein balance and in the absence of protein feeding, this balance becomes more negative. When combined with protein feeding, net muscle protein balance after exercise becomes positive 59. Norton and Layman 50 proposed that consumption of leucine, could turn a negative protein balance to a positive balance following an intense exercise bout by prolonging the MPS response to feeding. In support, the ingestion of a protein or essential amino acid complex that contains sufficient amounts of leucine has been shown to shift protein balance to a net positive state after intense exercise training 50. Even though leucine has been demonstrated to independently stimulate protein synthesis, it is important to recognize that supplementation should not be with just leucine alone. For instance, Wilson et al. 60 demonstrated in an animal model that leucine consumption resulted in a lower duration of protein synthesis compared to a whole meal. In summary, athletes should focus on consuming adequate leucine content in each of their meals through selection of high-quality protein sources 60.
Key points:
- Protein sources containing higher levels of the EAAs are considered to be higher quality sources of protein.
- The body uses 20 amino acids to make proteins, seven of which are essential (nine conditionally), requiring their ingestion to meet daily needs.
- EAAs appear to be uniquely responsible for increasing MPS with doses ranging from 6 to 15 g all exerting stimulatory effects. In addition, doses of approximately one to three g of leucine per meal appear to be needed to stimulate protein translation machinery.
- The BCAAs (i.e., isoleucine, leucine, and valine) appear to exhibit individual and collective abilities to stimulate protein translation. However, the extent to which these changes are aligned with changes in MPS remains to be fully explored.
- While greater doses of leucine have been shown to independently stimulate increases in protein synthesis, a balanced consumption of the EAAs promotes the greatest increases.
- The prioritization of feedings of protein with adequate levels of leucine/BCAAs will best promote increases in MPS.
Criteria for comparing protein sources
Some valid criteria exist to compare protein sources and provide an objective method of how to include them in a diet. As previously mentioned, common means of assessing protein quality include Biological Value, Protein Efficiency Ratio, PDCAAS and IAAO. The derivation of each technique is different with all having distinct advantages and disadvantages. For nearly all populations, ideal methods should be linked to the capacity of the protein to positively affect protein balance in the short term, and facilitate increases and decreases in lean and fat-mass, respectively, over the long term. In addition, the protein’s ability to enhance immune function and promote an anti-oxidative environment should also be considered. To this point, dairy, egg, meat, and plant-based proteins have been discussed.
Two critical variables exist that determine a protein’s impact on overall protein accretion and protein turnover: a) the protein’s leucine content and b) the rate at which the protein is digested.
In general, the proteins with the greatest leucine content include dairy (9–11%), egg (8.6%), and meat (8%), while sources low in leucine include plant-based proteins. Faster digesting sources of protein include whey and egg whites, soy, and very lean cuts of meat (>95% lean). In contrast, casein and fatty cuts of meat (<80% lean) act as slowly digested sources of protein. As mentioned previously, initial research by Boirie and Dangin has highlighted the impact of protein digestion rate on net protein balance with the two milk proteins: whey and casein 61. Subsequent follow-up work has used this premise as a reference point for the digestion rates of other protein sources.
Using the criteria of leucine content, Norton and Wilson et al. 62 used animal models to compare the potential to activate initiation factors and MPS between four different protein sources: wheat (supplemented with leucine), soy, egg, and whey, (containing 6.8, 8.0, 8.8, and 11% leucine, respectively) using a diet consisting of three meals per day. Macronutrient intake was 16/54/30% for protein, carbohydrates and fat, respectively. Wheat and soy did not stimulate MPS above fasted levels, whereas egg and whey proteins significantly increased MPS rates, with MPS for whey protein being greater than egg protein. MPS responses were closely related to changes in plasma leucine and phosphorylation of 4E–BP1 and S6 K protein signaling molecules. More importantly, following 2- and 11-weeks of ingestion, it was demonstrated that the leucine content of the meals increased muscle mass and was inversely correlated with body fat.
Tang et al. 63 compared high leucine/fast-digesting (hydrolyzed whey isolate), lower leucine/intermediate digesting (soy isolate) and high leucine/slow-digesting (micellar casein) protein sources on MPS at rest and following exercise. The researchers demonstrated that MPS at rest was higher after ingestion of faster digesting proteins compared to slower digesting proteins (whey and soy > casein). Specifically, MPS after consumption of whey was approximately 93% and 18% greater than casein and soy, respectively. A similar pattern of results was observed after resistance exercise (whey > soy > casein) whereby protein synthesis following whey consumption was approximately 122% and 31% greater than casein and soy, respectively. MPS was also greater after soy consumption at rest (64%) and following resistance exercise (69%) compared with casein. These findings lead us to conclude that athletes should seek protein sources that are both fast-digesting and high in leucine content to maximally stimulate rates of MPS at rest and following training. Moreover, in consideration of the various additional attributes that high-quality protein sources deliver, it may be advantageous to consume a combination of higher quality protein sources (dairy, egg, and meat sources).
Key points:
- Multiple protein sources are available for an athlete to consider, and each has their own advantages and disadvantages.
- Protein sources are commonly evaluated based upon the content of amino acids, particularly the EAAs, they provide. Beyond amino acid content, the fat, calorie, and micronutrient content, and presence of various bioactive peptides all contribute to a protein’s quality.
- Leucine content and rate of digestion have also been demonstrated in multiple scientific studies to play an important role in an athlete’s ability to train, compete, and recover.
- Blends of protein sources might afford a favorable combination of key nutrients such as leucine, EAAs, bioactive peptides, and antioxidants, but more research is needed to determine their ideal composition.
Preparation methods of various proteins
Nutrient density is defined as the amount of a particular nutrient (carbohydrate, protein, fat, etc.) per unit of energy in a given food. In many situations, the commercial preparation method of foods can affect the actual nutrient density of the resulting food. Using protein as an example, full-fat milk is approximately 150 cal a serving, and of this 8 g, or about 21% is from protein. Skim milk on the other hand contains approximately 9 g of protein in a 90-cal eight-ounce serving, making it approximately 40% protein. When producing milk protein supplements, special preparations must be made to separate the protein sources from the lactose and fat calories in milk. For example, the addition of acid to milk causes the casein to coagulate or collect at the bottom, while the whey is left on the top 64. These proteins are then filtered to increase their purity. A concentrate is commonly defined as any protein product that is 29–80% protein by dry weight. Sport nutrition products generally use concentrates that are 70–80% protein [200]. As extra filtering steps are added, the purity of the final product increases and when a final protein product yields greater than 90% protein, it is considered an isolated protein 64.
Filtration processes
Filtration methods differ, and there are both benefits and disadvantages to each. The two most popular methods of filtration of a given protein are the use of ion exchange and micro/ultrafiltration methods. Ion exchange exposes a given protein source, such as whey, to hydrochloric acid and sodium hydroxide, thereby producing an electric charge on the proteins that can be used to separate them from lactose and fat 64. The advantage of this method is that it is relatively cheap and produces the highest protein concentration 64. The disadvantage is that ion exchange filtration typically denatures some of the valuable immune-boosting, anti-carcinogenic peptides found in whey 64. Cross-flow microfiltration, and ultra-micro filtration are based on the premise that the molecular weight of whey protein is greater than lactose, and use 1 and 0.25-μm ceramic membranes, respectively, to separate the two. As a result, whey protein is trapped in the membranes but the lactose and other components pass through. The advantage is that these processes do not denature valuable proteins and peptides found in whey, so the protein itself is deemed to be of higher quality 64. The main disadvantage is that this filtration process is typically costlier than the ion exchange method.
Hydrolyzed proteins
When consumed whole, proteins are digested through a series of steps beginning with homogenization by chewing, followed by partial digestion by pepsin in the stomach 65. Following this, a combination of peptides, proteins, and negligible amounts of single amino acids are released into the small intestine and from there are either partially hydrolyzed into oligopeptides, 2–8 amino acids in length or are fully hydrolyzed into individual amino acids 65. Absorption of individual amino acids and various small peptides (di, tri, and tetra) into the blood occurs inside the small intestine through separate transport mechanisms 65. Oftentimes, products contain proteins that have been pre-exposed to specific digestive enzymes causing hydrolysis of the proteins into di, tri, and tetrapeptides. A plethora of studies have investigated the effects of the degree of protein fractionation (or degree of hydrolysis) on the absorption of amino acids and the subsequent hormonal response 66. Research indicates that amino acids are absorbed more rapidly when they are consumed as di and/or tri peptides compared to free form amino acids or complete proteins 67. Further, the rate of absorption may lead to a more favorable anabolic hormonal environment. Calbet et al. 68 examined both amino acid appearance and insulin responses following consumption of protein solutions containing the same amount of protein, or pure carbohydrates. The treatments consisted of a pure glucose solution, whey peptide hydrolysates, and cow’s milk containing milk proteins, lactose and fat. Each of the nitrogen containing solutions contained 15 g of glucose and 30 g of protein. Results indicated that peptide hydrolysates produced a faster increase in venous plasma amino acids compared to milk proteins. Further, the peptide hydrolysates produced peak plasma insulin levels that were two- and four-times greater than that evoked by the milk and glucose solutions, respectively, with a correlation of 0.8 between plasma amino acids and the insulin response in the peptide hydrolysates. One of the inherent shortcomings of this study is that milk proteins are 80% casein and, therefore, are not ideal candidates to compare with hydrolyzed whey.
In a more appropriate comparison, Morifuji et al. 67 investigated the effects of 12.5 g of either hydrolyzed or non-hydrolyzed soy and whey proteins on changes in plasma levels of the EAAs, BCAAs, and insulin. Results indicated that protein hydrolysates produced greater responses than their non-hydrolyzed counterpart in plasma for each of the variables (Hydrolyzed whey > Non-hydrolyzed whey > hydrolyzed soy > Non-hydrolyzed soy). However, Calbet et al. 69 found that 36 g of hydrolyzed or non-hydrolyzed whey and casein led to no differences in the plasma amino acid/BCAA responses in the whey groups. The hydrolyzed casein, however, did result in a greater amino acid response than the nonhydrolyzed casein. Finally, both hydrolyzed groups resulted in greater gastric secretions, as well as greater plasma increases, in glucose-dependent insulinotropic polypeptides 70.
Buckley and colleagues 66 found that a ~ 30 g dose of a hydrolyzed whey protein isolate resulted in a more rapid recovery of muscle force-generating capacity following eccentric exercise, compared with a flavored water placebo or a non-hydrolyzed form of the same whey protein isolate. Indeed, the effect of this hydrolysate was such that complete recovery of muscle force-generating capacity had been achieved by six hours post supplementation, while the normal whey and placebo groups’ strength remained depressed 24 h later. In agreement with these findings, Cooke et al. 71 had 17 untrained men complete an eccentric-based resistance training bout to invoke muscle damage and supplemented with either carbohydrate or a hydrolyzed whey protein isolate. Three and seven days after completing the damaging exercise bout, maximal strength levels were higher in the hydrolyzed whey protein group compared to carbohydrate supplementation. Additionally, blood concentrations of muscle damage markers tended to be lower when four ~30-g doses of a hydrolyzed whey protein isolate were ingested for two weeks following the damaging bout. Beyond influencing strength recovery after damaging exercise, other benefits of hydrolyzed proteins have been suggested. For example, Morifuji et al. 72 using an animal model reported that the ability of whey hydrolysates to increase skeletal muscle glycogen replenishment after exercise was greater when compared to BCAA ingestion. Furthermore, Lockwood et al. 73 investigated the effects of ingesting either 30 g of hydrolyzed whey or two varying forms of whey protein concentrates during a linear resistance-training protocol over 8 weeks. Results indicated that strength and lean body mass (LBM) increased equally in all groups. However, fat mass decreased only in the hydrolyzed whey protein group. While more work needs to be completed to fully determine the potential impact of hydrolyzed proteins on strength and body composition changes, this initial study suggests that hydrolyzed whey may be efficacious for decreasing body fat. Finally, Saunders et al. 74 had thirteen trained male cyclists complete a simulated 60-km time trial where they ingested either carbohydrate or carbohydrate and protein hydrolysate at equal intervals throughout the race as well as at the conclusion of the race. The authors reported that co-ingestion of a carbohydrate and protein hydrolysate improved time-trial performance late in the exercise protocol and significantly reduced soreness and markers of muscle damage. Two excellent reviews on the topic of hydrolyzed proteins and their impact on performance and recovery have been published by Van Loon et al. 75 and Saunders 76.
Digestive enzymes in proteins
Digestion is the physiological process of rendering the food we eat into smaller components that allow key nutrients to be assimilated into our body’s tissues. The prevalence of digestive enzymes in sports nutrition products has increased during recent years with many products now containing a combination of proteases and lipases, with the addition of carbohydrates in plant proteins. Proteases can hydrolyze proteins into various peptide configurations and potentially single amino acids. It appears that digestive enzyme capabilities and production decrease with age 77, thus increasing the difficulty with which the body can break down and digest large meals. Digestive enzymes could potentially work to promote optimal digestion by allowing up-regulation of various metabolic enzymes that may be needed to allow for efficient bodily operation. Further, digestive enzymes have been shown to minimize quality differences between varying protein sources 78. Individuals looking to increase plasma peak amino acid concentrations may benefit from hydrolyzed protein sources or protein supplemented with digestive enzymes. However, more work is needed before definitive conclusions can be drawn regarding the efficacy of digestive enzymes.
Protein sources
Animal Protein
Proteins from animal sources (i.e. eggs, milk, meat, fish and poultry) provide the highest quality rating of food sources. This is primarily due to the ‘completeness’ of proteins from these sources. Although protein from these sources are also associated with high intakes of saturated fats and cholesterol, there have been a number of studies that have demonstrated positive benefits of animal proteins in various population groups 79.
Protein from animal sources during late pregnancy is believed to have an important role in infants born with normal body weights. Godfrey et al. 80 examined the nutrition behavior of more than 500 pregnant women to determine the effect of nutritional intake on placental and fetal growth. They reported that a low intake of protein from dairy and meat sources during late pregnancy was associated with low birth weights.
In addition to the benefits from total protein consumption, elderly subjects have also benefited from consuming animal sources of protein. Diets consisting of meat resulted in greater gains in lean body mass compared to subjects on a lactoovovegetarian diet 79. High animal protein diets have also been shown to cause a significantly greater net protein synthesis than a high vegetable protein diet 81. This was suggested to be a function of reduced protein breakdown occurring during the high animal protein diet.
There have been a number of health concerns raised concerning the risks associated with protein emanating primarily from animal sources. Primarily, these health risks have focused on cardiovascular disease (due to the high saturated fat and cholesterol consumption), bone health (from bone resorption due to sulfur-containing amino acids associated with animal protein) and other physiological system disease.
Milk proteins
Milk proteins have undergone extensive research related to their potential roles in augmenting adaptations from exercise training 82. For example, consuming milk following exercise has been demonstrated to accelerate recovery from muscle damaging exercise 83, increase glycogen replenishment 84, improve hydration status 85, and improve protein balance to favor synthesis 82, ultimately resulting in increased gains in both neuromuscular strength and skeletal muscle hypertrophy 82. Moreover, milk protein contains the highest score on the PDCAAS rating system, and in general contains the greatest density of leucine 53. Milk can be fractionated into two protein classes, casein and whey.
Comparison of the quality of whey and casein reveal that these two proteins routinely contain the highest leucine content of all other protein sources at 11% and 9.3%, respectively. While both are high in quality, the two differ in the rate at which they digest as well as the impact they have on protein metabolism 86. Whey protein is water soluble, mixes easily, and is rapidly digested. In contrast, casein is water insoluble, coagulates in the gut and is digested more slowly than whey protein 87. Casein also has intrinsic properties such as opioid peptides, which effectively slow gastric motility 87. Original research investigating the effects of digestion rate was conducted by Boirie, Dangin and colleagues 88. These researchers gave a 30 g bolus of whey protein and a 43 g bolus of casein protein to subjects on separate occasions and measured amino acid levels for several hours after ingestion. They reported that the whey protein condition displayed robust hyperaminoacidemia 100 min after administration. However, by 300 min, amino acid concentrations had returned to baseline. In contrast, the casein condition resulted in a slow increase in amino acid concentrations, which remained elevated above baseline after 300 min. Over the study duration, casein produced a greater whole body leucine balance than the whey protein condition, leading the researcher to suggest that prolonged, moderate hyperaminoacidemia is more effective at stimulating increases in whole body protein anabolism than a robust, short lasting hyperaminoacidemia.
While this research appears to support the efficacy of slower digesting proteins, subsequent work has questioned its validity in athletes. The first major criticism is that Boire and colleagues investigated whole body (non-muscle and muscle) protein balance instead of skeletal (myofibrillar) MPS. This is important considering that skeletal muscle protein turnover occurs at a much slower rate than protein turnover of both plasma and gut proteins; as a result, MPS has been suggested to contribute anywhere from 25 to 50% of total whole body protein synthesis 89. These findings suggest that changes in whole body protein turnover may poorly reflect the level of skeletal muscle protein metabolism that may be taking place. Trommelen and investigators 90 examined 24 young men ingesting 30 g of casein protein with or without completion of a single bout of resistance exercise, and concluded that rates of MPS were increased, but whole-body protein synthesis rates were not impacted.
More recently, Tang and colleagues 63 investigated the effects of administering 22 g of hydrolyzed whey isolate and micellar casein (10 g of EAAs) at both rest and following a single bout of resistance training in young males. The area under the curve calculations demonstrated a 200% greater increase in leucine concentrations in the blood following whey versus casein ingestion. Moreover, these researchers reported that whey protein ingestion stimulated greater MPS at both rest and following exercise when compared to casein. Tipton et al. 91 used an acute study design involving a single bout of lower body resistance exercise and 20-g doses of casein or whey after completing the exercise session. In comparison to the control group, both whey and casein significantly increased leucine balance, but no differences were found between the two protein sources for amino acid uptake and muscle protein balance. Additional research has also demonstrated that 10 weeks of whey protein supplementation in trained bodybuilders resulted in greater gains in lean mass (5.0 vs. 0.8 kg) and strength compared to casein 92. These findings suggest that the faster-digesting whey proteins may be more beneficial for skeletal muscle adaptations than the slower digesting casein.
Effects of milk proteins on glycogen replenishment and skeletal muscle damage
Skeletal muscle glycogen stores are a critical element to both prolonged and high-intensity exercise. In skeletal muscle, glycogen synthase activity is considered one of the key regulatory factors for glycogen synthesis. Research has demonstrated that the addition of protein in the form of milk and whey protein isolate (0.4 g/kg) to a moderate (0.8 g/kg), but not high (1.2 g/kg) carbohydrate-containing (dextrose-maltodextrin) beverage promotes increased rates of muscle glycogen replenishment following hard training 93. Further, the addition of protein facilitates repair and recovery of the exercised muscle 94. These effects are thought to be related to a greater insulin response following the exercise bout. Intriguingly, it has also been demonstrated that whey protein enhances glycogen synthesis in the liver and skeletal muscle more than casein in an insulin-independent fashion that appears to be due to its capacity to upregulate glycogen synthase activity 95. Therefore, the addition of milk protein to a post-workout meal may augment recovery, improve protein balance, and speed glycogen replenishment.
Health benefits of milk-based proteins
While athletes tend to view whey as the ideal protein for skeletal muscle repair and function it also has several health benefits. In particular, whey protein contains an array of biologically active peptides whose amino acids sequences give them specific signaling effects when liberated in the gut. Not only is whey protein high in β-Lactoglobulin and α-lactalbumin (75% of total bovine whey proteins), but it is also rich in EAAs (approximately 50% by weight). Furthermore, whey protein appears to play a role in enhancing lymphatic and immune system responses 96. In addition, α-lactalbumin contains an ample supply of tryptophan which increases cognitive performance under stress 97, improves the quality of sleep 97 and may also speed wound healing 97, properties which could be vital for recovery from combat and contact sporting events. In addition, lactoferrin is also found in both milk and in whey protein, and has been demonstrated to have antibacterial, antiviral, and antioxidant properties 98. Moreover, there is some evidence that whey protein can bind iron and therefore increase its absorption and retention 99.
Bovine Colostrum
Bovine colostrum is the “pre” milk liquid secreted by female mammals the first few days following birth. This nutrient-dense fluid is important for the newborn for its ability to provide immunities and assist in the growth of developing tissues in the initial stages of life. Evidence exists that bovine colostrum contains growth factors that stimulate cellular growth and DNA synthesis 100, and as might be expected with such properties, it makes for interesting choice as a potential sports supplement.
Although bovine colostrum is not typically thought of as a food supplement, the use by strength/power athletes of this protein supplement as an ergogenic aid has become common. Oral supplementation of bovine colostrum has been demonstrated to significantly elevate insulin-like-growth factor 1 (IGF-1) and enhance lean tissue accruement. However, the results on athletic performance improvement are less conclusive. Mero and colleagues (1997) reported no changes in vertical jump performance following 2-weeks of supplementation, and Brinkworth and colleagues 101 saw no significant differences in strength following 8-weeks of training and supplementation in both trained and untrained subjects. In contrast, following 8-weeks of supplementation significant improvements in sprint performance were seen in elite hockey players 102. Further research concerning bovine colostrum supplementation is still warranted.
Whey Protein
Whey is a general term that typically denotes the translucent liquid part of milk that remains following the process (coagulation and curd removal) of cheese manufacturing. From this liquid, whey proteins are separated and purified using various techniques yielding different concentrations of whey proteins. Whey is one of the two major protein groups of bovine milk, accounting for 20% of the milk while casein accounts for the remainder. All of the constituents of whey protein provide high levels of the essential and branched chain amino acids. The bioactivities of these proteins possess many beneficial properties as well. Additionally, whey is also rich in vitamins and minerals. Whey protein is most recognized for its applicability in sports nutrition. Additionally, whey products are also evident in baked goods, salad dressings, emulsifiers, infant formulas, and medical nutritional formulas.
Varieties of Whey Protein
There are three main forms of whey protein that result from various processing techniques used to separate whey protein. They are whey powder, whey concentrate, and whey isolate. Table 2 provides the composition of Whey Proteins.
Table 2. Composition (%) of whey protein forms
| Component | Whey Powder | Whey Concentrate | Whey Isolate |
|---|---|---|---|
| Protein | 11 – 14.5 | 25 – 89 | 90 + |
| Lactose | 63 – 75 | 10 – 55 | 0.5 |
| Milk Fat | 1 – 1.5 | 2 – 10 | 0.5 |
Whey Protein Powder
Whey protein powder has many applications throughout the food industry. As an additive it is seen in food products for beef, dairy, bakery, confectionery, and snack products. Whey powder itself has several different varieties including sweet whey, acid whey (seen in salad dressings), demineralized (seen primarily as a food additive including infant formulas), and reduced forms. The demineralized and reduced forms are used in products other than sports supplements.
Whey Protein Concentrate
The processing of whey concentrate removes the water, lactose, ash, and some minerals. In addition, compared to whey isolates whey concentrate typically contains more biologically active components and proteins that make them a very attractive supplement for the athlete.
Whey Protein Isolate (WPI)
Isolates are the purest protein source available. Whey protein isolates contain protein concentrations of 90% or higher. During the processing of whey protein isolate there is a significant removal of fat and lactose. As a result, individuals who are lactose-intolerant can often safely take these products 103. Although the concentration of protein in this form of whey protein is the highest, it often contain proteins that have become denatured due to the manufacturing process. The denaturation of proteins involves breaking down their structure and losing peptide bonds and reducing the effectiveness of the protein.
Whey is a complete protein whose biologically active components provide additional benefits to enhance human function. Whey protein contains an ample supply of the amino acid cysteine. Cysteine appears to enhance glutathione levels, which has been shown to have strong antioxidant properties that can assist the body in combating various diseases 104. In addition, whey protein contains a number of other proteins that positively effect immune function such as antimicrobial activity 105. Whey protein also contains a high concentration of branched chain amino acids (BCAA) that are important for their role in the maintenance of tissue and prevention of catabolic actions during exercise 106.
Casein
Casein is the major component of protein found in bovine milk accounting for nearly 70-80% of its total protein and is responsible for the white color of milk. It is the most commonly used milk protein in the industry today. Milk proteins are of significant physiological importance to the body for functions relating to the uptake of nutrients and vitamins and they are a source of biologically active peptides. Similar to whey, casein is a complete protein and also contains the minerals calcium and phosphorous. Casein has a PDCAAS rating of 1.23 (generally reported as a truncated value of 1.0) 107.
Casein exists in milk in the form of a micelle, which is a large colloidal particle. An attractive property of the casein micelle is its ability to form a gel or clot in the stomach. The ability to form this clot makes it very efficient in nutrient supply. The clot is able to provide a sustained slow release of amino acids into the blood stream, sometimes lasting for several hours 108. This provides better nitrogen retention and utilization by the body.
Egg proteins
Egg protein is often thought of as an ideal protein because its amino acid profile has been used as the standard for comparing other dietary proteins 87. Due to their excellent digestibility and amino acid content, eggs are an excellent source of protein for athletes. While the consumption of eggs has been criticized due to their cholesterol content, a growing body of evidence demonstrates the lack of a relationship between egg consumption and coronary heart disease, making egg-based products more appealing 109. One large egg has 75 kcal and 6 g of protein, but only 1.5 g of saturated fat while one large egg white has 16 kcal with 3.5 g of protein and is fat-free. Research using eggs as the protein source for athletic performance and body composition is lacking, perhaps due to less funding opportunities relative to funding for dairy. Egg protein may be particularly important for athletes, as this protein source has been demonstrated to significantly increase protein synthesis of both skeletal muscle and plasma proteins after resistance exercise at both 20 and 40 g doses. Leucine oxidation rates were found to increase following the 40 g dose, suggesting that this amount exceeds an optimal dose. In addition to providing a cost effective, high-quality source of protein rich in leucine (0.5 g of leucine per serving), eggs have also been identified as a functional food 110. Functional foods are defined as foods that, by the presence of physiologically active components, provide a health benefit beyond basic nutrition 111. According to the Academy of Nutrition and Dietetics, functional foods should be consumed as part of a varied diet on a regular basis, at effective levels. Thus, it is essential that athletes select foods that meet protein requirements and also optimize health and prevent decrements in immune function following intense training. Important nutrients provided by eggs include riboflavin (15% RDA), selenium (17% RDA) and vitamin K (31% RDA) 110. Eggs are also rich in choline, a nutrient which may have positive effects on cognitive function 112. Moreover, eggs provide an excellent source of the carotenoid-based antioxidants lutein and zeaxanthin 113. Also, eggs can be prepared with most meal choices, whether at breakfast, lunch, or dinner. Such positive properties increase the probability of the athletes adhering to a diet rich in egg protein.
Beef and other flesh proteins
Meat proteins are a major staple in the American diet and, depending on the cut of meat, contain varying amounts of fat and cholesterol. Meat proteins are well known to be rich sources of the EAAs 114. Beef is a common source of dietary protein and is considered to be of high biological value because it contains the full balance of EAAs in a fraction similar to that found in human skeletal muscle 114. A standard serving of 113.4 g lean beef provides 10 g of the EAAs (3.5 g of leucine) and 30 g of total amino acids. Moreover, this 30 g dose of beef protein has been shown to stimulate protein synthesis in both young and elderly subjects 114. In addition to its rich content of amino acids, beef and other flesh proteins can serve as important sources of micronutrients such as iron, selenium, vitamins A, B12 and folic acid. For the most part, these quality minerals and micronutrients cannot be as easily obtained through plant-based proteins and/or the bioavailability of these macronutrients from plants is limited. This is a particularly important consideration for pregnant and breastfeeding women. Ultimately, as an essential part of a mixed diet, meat helps to ensure adequate distribution of essential micronutrients and amino acids to the body.
Research has shown that significant differences in skeletal muscle mass and body composition between older men who resistance train and either consume meat-based or lactoovovegetarian diet 44. Over a 12-week period, whole-body density, fat-free mass, and whole-body muscle mass (as measured by urinary creatinine excretion) increased in the meat-sourced diet group but decreased in the lactoovovegetarian diet group. These results indicate that not only do meat-based diets increase fat-free mass, but also they may specifically increase muscle mass, thus supporting the many benefits of meat-based diets. A diet high in meat protein in older adults may provide an important resource in reducing the risk of sarcopenia.
Positive results have also been seen in elite athletes that consume meat-based proteins, as opposed to vegetarian diets 115. For example, carnitine is a molecule that transports long-chain fatty acids into mitochondria for oxidation and is found in high amounts in meat. While evidence is lacking to support an increase in fat oxidation with increased carnitine availability, carnitine has been linked to the sparing of muscle glycogen, and decreases in exercise-induced muscle damage 116. Certainly, more research is needed to support these assertions. Creatine is a naturally occurring compound found mainly in muscle. The concentration of creatine in uncooked chicken and beef is approximately 30 mmol/kg (4–5 g/kg), meaning that one serving of beef contains approximately 0.4 g of creatine 117. Vegetarians have lower total body creatine stores than omnivores, which demonstrates that regular meat eating has a significant effect on human creatine status 118. Moreover, creatine supplementation studies with vegetarians indicate that increased creatine uptake levels do exist in people who practice various forms of vegetarianism. Sharp and investigators 119 published the only study known to compare different supplemental (powdered) forms of animal proteins on adaptations to resistance training such as increases in strength and improvements in body composition. Forty-one men and women performed a standardized resistance-training program over eight weeks and consumed a daily 46 g dose of either hydrolyzed chicken protein, beef protein isolate, or whey protein concentrate in comparison to a control group. All groups experienced similar increases in upper and lower-body strength, but all protein-supplemented groups reported significant increases in lean mass and decreases in fat mass.
Meat-based diets have been shown to include additional overall health benefits. Some studies have found that meat, as a protein source, is associated with higher serum levels of IGF-1 120, which in turn is related to increased bone mineralization and fewer fractures 121.
Meat vs. plant based proteins: Is one better than the other ?
A highly debated topic in nutrition and epidemiology is whether vegetarian diets are a healthier choice than omnivorous diets. One key difference is the fact that vegetarian diets often lack equivalent amounts of protein when compared to omnivorous diets 44. However, with proper supplementation and careful nutritional choices, it is possible to have complete proteins in a vegetarian diet. Generally by consuming high-quality, animal-based products (meat, milk, eggs, and cheese) an individual will achieve optimal growth as compared to ingesting only plant proteins 44. Research has shown that soy is considered a lower quality complete protein. Hartman et al. 82 had participants consume a mixture of sucrose and either 30 g of milk or soy proteins during 12-weeks of resistance training. They found that the participants that consumed the milk protein increased lean mass and decreased fat mass more than the control and soy groups. Moreover, the soy group was not significantly different from the control group. Similarly, a study by Tang and colleagues 63 directly compared the abilities of hydrolyzed whey isolate, soy isolate, and micellar casein to stimulate rates of MPS both at rest and in response to a single bout of lower body resistance training. These authors reported that the ability of soy to stimulate MPS was greater than casein, but less than whey, at rest and in response to an acute resistance exercise stimulus. While soy is considered a complete protein, it contains lower amounts of BCAAs than bovine milk 87. Additionally, research has found that dietary soy phytoestrogens inhibit mTOR expression in skeletal muscle through activation of AMPK 122. Thus, not only does soy contain lower amounts of the EAAs and leucine, but soy protein may also be responsible for inhibiting growth factors and protein synthesis via its negative regulation of mTOR. When considering the multitude of plant sources of protein, soy overwhelmingly has the most research. Limited evidence using wheat protein in older men has suggested that wheat protein stimulates significantly lower levels of MPS when compared to an identical dose (35 g) of casein protein, but when this dose is increased nearly two fold (60 g) this protein source is able to significantly increase rates of myofibrillar protein synthesis 123. Rice protein is a medium to slow absorbing protein, which is in line with other non-meat/non-dairy proteins, however, leucine from rice protein shows unique absorption kinetics, peaking faster than leucine from whey protein 124. As mentioned earlier, a study by Joy and colleagues 125 in which participants participated in resistance training program for eight weeks while taking identical, high doses of either rice or whey protein, demonstrated that rice protein stimulated similar increases in body composition adaptations to whey protein.
Protein blends
The majority of available science has explored the efficacy of ingesting single protein sources, but evidence continues to mount that combining protein sources may afford additional benefits 126. For example, a 10-week resistance training study by Kerksick and colleagues 127 demonstrated that a combination of whey (40 g) and casein (8 g) yielded the greatest increase in fat-free mass (determined by DEXA) when compared to both a combination of 40 g of whey, 5 g of glutamine, and 3 g of BCAAs and a placebo consisting of 48 g of a maltodextrin carbohydrate. Later, Kerksick et al. 128 demonstrated various combinations of whey, casein, and colostrum proteins with and without creatine can also yield positive improvements in strength and body composition over a 12-week resistance training and supplementation regimen. Similarly, Hartman and investigators 82 had 56 healthy young men train for 12 weeks while either ingesting isocaloric and isonitrogenous doses of fat-free milk (a blend of whey and casein), soy protein or a carbohydrate placebo and concluded that fat-free milk stimulated the greatest increases in Type I and II muscle fiber area as well as fat-free mass; however, strength outcomes were not affected. Moreover, Wilkinson and colleagues 129 demonstrated that ingestion of fat-free milk (vs. soy or carbohydrate) led to a greater area under the curve for net balance of protein and that the fractional synthesis rate of muscle protein was greatest after milk ingestion. In 2013, Reidy et al. 130 indicated that a mixture of whey and soy protein over a four-hour measurement window similarly increased MPS rates during the early (0–2 h) time-period versus whey protein, but only the protein blend was able to stimulate significantly increased MPS rates during the later (2–4 h) measurement window. However, when the entire four-hour measurement period was considered, no difference in MPS rates were found. A follow-up publication from the same clinical trial also reported that ingestion of the protein blend resulted in a positive and prolonged amino acid balance when compared to ingestion of whey protein alone, while post-exercise rates of myofibrillar protein synthesis were similar between the two conditions 131. Reidy et al. 132 reported that in 68 healthy young men who were participating in a supervised resistance-training program over 12 weeks, there were increases in whole body lean mass with either whey protein or a whey protein and soy protein blend compared to a maltodextrin placebo. No differences were found between whey and the whey and soy blend.
Protein safety
Despite a plethora of studies demonstrating safety, much concern still exists surrounding the clinical implications of consuming increased amounts of protein, particularly on renal and hepatic health. The majority of these concerns stem from renal failure patients and educational dogma that has not been rewritten as evidence mounts to the contrary. Certainly, it is clear that people in renal failure benefit from protein-restricted diets 133, but extending this pathophysiology to otherwise healthy exercise-trained individuals who are not clinically compromised is inappropriate. Published reviews on this topic consistently report that an increased intake of protein by competitive athletes and active individuals provides no indication of hepato-renal harm or damage 134. This is supported by a recent commentary which referenced recent reports from the World Health Organization 135 where they indicated a lack of evidence linking a high protein diet to renal disease. Likewise, the panel charged with establishing reference nutrient values for Australia and New Zealand also stated there was no published evidence that elevated intakes of protein exerted any negative impact on kidney function in athletes or in general 136.
Recently, Antonio and colleagues published a series of original investigations that prescribed extremely high amounts of protein (~3.4–4.4 g/kg/day) and have consistently reported no harmful effects 137. The first study in 2014 had resistance-trained individuals consume an extremely high protein diet (4.4 g/kg/day) for eight weeks and reported no change in adverse outcomes (Antonio J, Peacock CA, Ellerbroek A, Fromhoff B, Silver T. The effects of consuming a high protein diet (4.4 g/kg/d) on body composition in resistance-trained individuals. J Int Soc Sports Nutr. 2014;11:19. doi: 10.1186/1550-2783-11-19. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4022420/)). A follow-up investigation 138 required participants to ingest up to 3.4 g/kg/day of protein for eight weeks while following a prescribed resistance training program and reported no changes in any of the blood parameters commonly used to assess clinical health (e.g., there was no effect on kidney or liver function). Their next study employed a crossover study design in twelve healthy resistance-trained men in which each participant was tested before and after for body composition as well as blood-markers of health and performance 139. In one eight-week block, participants followed their normal (habitual) diet (2.6 g/kg/day) and in the other eight-week block, participants were prescribed to ingest greater than 3.0 g/kg/day resulting in an average protein intake of 2.9 g/kg/day over the entire 16-week study. No changes in body composition were reported, and importantly, no clinical side effects were observed throughout the study. Finally, the same group of authors published a one-year crossover study 140 in fourteen healthy resistance-trained men. When prescribed to a high protein diet, the participants were instructed to ingest 3 g/kg/day and achieved an average intake of 3.3 g/kg/day and when following their normal diet they consumed 2.5 g/kg/day. This investigation showed that the chronic consumption of a high protein diet (i.e., for 1 year) had no harmful effects on kidney or liver function. Furthermore, there were no alterations in clinical markers of metabolism and blood lipids.
Key points
- Multiple review articles indicate that no controlled scientific evidence exists indicating that increased intakes of protein pose any health risks in healthy, exercising individuals.
- Series of controlled investigations spanning up to one year in duration utilizing protein intakes of up to 2.5–3.3 g/kg/day in healthy resistance-trained individuals consistently indicate that increased intakes of protein exert no harmful effect on blood lipids or markers of kidney and liver function.
Adjunctive Nutritional Strategies to Augment Muscle Protein Synthesis
While it is clear that aminoacidemia following protein ingestion drives the rise in muscle protein synthesis, other nutrients have been added to protein in an attempt to augment its impact on muscle protein synthesis. Carbohydrates have been a primary focus in this area, with the rationale that their energy may serve to reverse an exercise-induced suppression of protein synthesis, either by activation of adenosine monophosphate kinase 141 or through a calcium–calmodulin-dependent mechanism 142. Alternatively, insulin as a result of carbohydrate ingestion could either promote protein synthesis, suppress proteolysis, or both 143. However, to date several studies combining protein and carbohydrate have shown no augmentation of protein synthesis when protein is provided in adequate amounts 144, 35, 145. However, these data do not preclude the hypothesis that carbohydrate is not stimulatory with lower-than-optimal protein doses. In addition, the restoration of muscle glycogen by means of carbohydrate ingestion is also obviously important for athletes and should not be neglected.
Only a few amino acids have been tested in their capacity to augment muscle protein synthesis, but none has proved beneficial in young men. Glutamine (0.3 g/kg BM) was given to young men following 90 min of cycling at 65 % of peak oxygen uptake in addition to carbohydrate and balanced EAA, and there was no difference in post-exercise muscle protein synthesis compared with the placebo trial 146. The lack of an effect of glutamine on muscle protein synthesis following endurance exercise is at odds with data showing that even endurance exercise is anabolic for mitochondrial and myofibrillar protein synthesis 147. Congruent with the absence of any benefit of glutamine on muscle protein synthesis after endurance exercise are data from young men performing resistance training who received glutamine throughout 6 weeks of training (0.9 g/kg lean tissue/day) 148. Glutamine supplementation has been shown to be useful in certain clinical populations, in whom there is a relative lack of intracellular glutamine 149. However, it is perhaps not overly surprising that glutamine is ineffective in populations who have adequate levels of the amino acid, because it is hard for even a high dose of glutamine to increase intramuscular glutamine 148, and conclusions of recent reviews have been that glutamine appears to be far from useful for athletes 150.
As a precursor for nitric oxide biosynthesis, the amino acid arginine has received some attention for its potential role to promote blood flow and enhance nutrient or hormonal delivery to muscles allowing enhanced anabolism 151. The one study in which muscle protein synthesis has been measured in humans following exercise with arginine supplementation showed no effect of a bolus dose (10 g) of arginine on nitrate or nitrite concentration, femoral artery flow, or muscle protein synthesis 152. An interesting observation was that growth hormone concentrations were enhanced by arginine supplementation 152 but, similar to other studies 153, the transiently increased growth hormone concentration did not enhance muscle protein synthesis. Other attempts to enhance blood flow after resistance exercise by means of arginine or other nitric oxide-enhancing compounds have proved unsuccessful, at least in healthy young men 154.
Protein Recommended intake
Proteins provide the building blocks of all tissues via their constituent amino acids. Athletes consume dietary protein to repair and rebuild skeletal muscle and connective tissues following intense training bouts or athletic events. During in the 1980s and early 1990’s Tarnopolsky 155, Phillips 156, and Lemon 157 first demonstrated that total protein needs were 50 to 175% greater in athletes than sedentary subjects. Adequate protein consumption during a body building contest preparation is required to support maintenance of lean body mass. Athletes require higher protein intakes to support increased activity and strength athletes benefit from higher intakes to support growth of LBM 158. Some researchers suggest these requirements increase further when athletes undergo energy restriction 159. Furthermore, there is evidence that protein requirements are higher for leaner individuals in comparison to those with higher body fat percentages 160, 161.
The collective agreement among reviewers is that a protein intake of 1.2-2.2 g/kg is sufficient to allow adaptation to training for athletes whom are at or above their energy needs. However, bodybuilders during their contest preparation period typically perform resistance and cardiovascular training, restrict calories and achieve very lean conditions. Each of these factors increases protein requirements and when compounded may further increase protein needs. Therefore, optimal protein intakes for bodybuilders during contest preparation may be significantly higher than existing recommendations.
In support of this notion, Butterfield et al. 162 found that male athletes running five to 10 miles per day during a slight caloric deficit were in a significant negative nitrogen balance despite consuming 2 g/kg of protein daily. Celejowa et al. 163 showed that five out of 10 competitive weight lifters achieved a negative nitrogen balance over the course of a training camp while consuming an average protein intake of 2 g/kg. Out of these five, as many as three were in a caloric deficit. The authors concluded that a protein intake of 2–2.2 g/kg under these conditions only allows for a small margin of error before nitrogen losses occur.
Walberg et al. 164 examined the effects of two energy restricted isocaloric diets of differing protein intakes in 19 lean (9.1-16.7% body fat), male, non-competitive body builders. One group consumed a protein intake of 0.8 g/kg and higher carbohydrates, while the other consumed 1.6 g/kg of protein with lower carbohydrates. The length of the intervention was only one week, but nonetheless nitrogen losses occurred only in the lower protein group and LBM decreased by a mean of 2.7 kg in the 0.8 g/kg protein group and by a mean of 1.4 kg in the 1.6 g/kg protein group. While the high protein group mitigated LBM losses compared to the low protein group, they were not eliminated.
A recent study by Mettler et al. 165 employed the same basic methodology as Walberg et al. 164. However, one group consumed a protein intake of 1 g/kg, while the other consumed 2.3 g/kg. The high-protein group lost significantly less LBM (0.3 kg) over the course of the two week intervention compared to the low-protein group (1.6 kg). Unlike Walberg et al. 164 calorie balance between diets was maintained by reducing dietary fat as opposed to carbohydrate to allow for the increase in protein.
While it appears that the 2.3 g/kg protein intervention in Mettler et al. 165 was superior for maintaining LBM compared to 1.6 g/kg in Walberg et al. 164 a recent study by Pasiakos et al. 166 found a trend towards the opposite. In this study, a non-significant trend of greater LBM retention occurred when subjects consumed 1.6 g/kg of protein compared to 2.4 g/kg of protein. However, the participants were intentionally prescribed low volume, low intensity resistance training “to minimize the potential of an unaccustomed, anabolic stimulus influencing study outcome measures”. Thus, the non-anabolic nature of the training may not have increased the participants’ protein requirements to the same degree as the participants in Mettler et al. 165 or to what would be expected among competitive bodybuilders.
Maestu et al. 167 did not observe a significant loss of LBM in a group of drug free bodybuilders consuming 2.5-2.6 g/kg of protein during the 11 weeks prior to competition. These results when considered alongside the works by Walberg et al. 164 and Mettler et al. 165 imply that the higher the protein intake, the lower the chance for LBM loss. However, it should be noted that this study did not include a low protein control and not all studies show a linear increase in LBM preservation with increases in protein 166. Furthermore, two subjects did lose significant amounts of LBM (1.5 kg and 1.8 kg), and the authors noted that these specific bodybuilders were among the leanest of the subjects. These two subjects lost the majority of their LBM (approximately 1 kg) during the latter half of the intervention as their percentage of calories from protein increased from 28% to 32-33% by the end of the study. The group as a whole progressively decreased their calories by reducing all three macronutrients throughout the investigation. Thus, the two subjects uniquely increased their proportion of protein, possibly reducing fat and carbohydrate to the point of detriment. That said it is also plausible that the lost LBM seen by these two subjects was necessary in order to achieve their low levels of body fat. It is unknown whether or not the lost LBM influenced their competitive outcome and it is possible that had the competitors not been as lean, they may have retained more LBM but also not have placed as well.
In a review by Phillips and Van Loon 158, it is suggested that a protein intake of 1.8-2.7 g/kg for athletes training in hypocaloric conditions may be optimal. While this is one of the only recommendations existing that targets athletes during caloric restriction, this recommendation is not given with consideration to bodybuilders performing concurrent endurance and resistance training at very low levels of body fat. However, the recently published systematic review by Helms et al. 159 on protein intakes in resistance-trained, lean athletes during caloric restriction suggests a range of 2.3-3.1 g/kg of LBM, which may be more appropriate for bodybuilding. Moreover, the authors suggest that the lower the body fat of the individual, the greater the imposed caloric deficit and when the primary goal is to retain LBM, the higher the protein intake (within the range of 2.3-3.1 g/kg of LBM) should be.
For example, Moore 21 found that muscle and albumin protein synthesis was optimized at approximately 20 g of egg protein at rest. Witard et al. 168 provided incremental doses of whey protein (0, 10, 20 and 40 g) in conjunction with an acute bout of resistance exercise and concluded that a minimum protein dose of 20 g optimally promoted muscle protein synthesis rates. Finally, Yang and colleagues 169 had 37 elderly men (average age of 71 years) consume incremental doses of whey protein isolate (0, 10, 20 and 40 g/dose) in combination with a single bout of lower body resistance exercise and concluded that a 40 g dose of whey protein isolate is needed in this population to maximize rates of muscle protein synthesis. Furthermore, while results from these studies offer indications of what optimal absolute dosing amounts may be, Phillips 170 concluded that a relative dose of 0.25 g of protein per kg of body weight per dose might operate as an optimal supply of high-quality protein. Once a total daily target protein intake has been achieved, the frequency and pattern with which optimal doses are ingested may serve as a key determinant of overall changes in protein synthetic rates.
Research indicates that rates of muscle protein synthesis rapidly rise to peak levels within 30 min of protein ingestion and are maintained for up to three hours before rapidly beginning to lower to basal rates of muscle protein synthesis even though amino acids are still elevated in the blood 171. Using an oral ingestion model of 48 g of whey protein in healthy young men, rates of myofibrillar protein synthesis increased three-fold within 45–90 min before slowly declining to basal rates of muscle protein synthesis all while plasma concentration of EAAs remained significantly elevated 172. While human models have not fully explored the mechanistic basis of this ‘muscle-full’ phenomenon, an energy deficit theory has been proposed which hypothesizes that rates of muscle protein synthesis were blunted even though plasma concentrations of amino acids remained elevated because a relative lack of cellular ATP was available to drive the synthetic process 60. While largely unexplored in a human model, these authors relied upon an animal model and were able to reinstate increases in muscle protein synthesis using the consumption of leucine and carbohydrate 135 min after ingestion of the first meal. As such, it is suggested that individuals attempting to restrict caloric intake should consume three to four whole meals consisting of 20–40 g of protein per meal. While this recommendation stems primarily from initial work that indicated protein doses of 20–40 g favorably promote increased rates of muscle protein synthesis, Kim and colleagues 173 recently reported that a 70 g dose of protein promoted a more favorable net balance of protein when compared to a 40 g dose due to a stronger attenuation of rates of muscle protein breakdown.
Key points
- The current RDA for protein is 0.8 g/kg/day with multiple lines of evidence indicating this value is not an appropriate amount for a training athlete to meet their daily needs.
- While previous recommendations have suggested a daily intake of 1.2–1.3 g/kg/day is an appropriate amount, most of this work was completed using the nitrogen balance technique, which is known to systematically underestimate protein needs.
- Daily and per dose needs are combinations of many factors including volume of exercise, age, body composition, total energy intake and training status of the athlete.
- Daily intakes of 2.3 to 3.1 g/kg per day operate as a minimum recommended amount while greater amounts may be needed for people attempting to restrict energy intake while maintaining fat-free mass.
- Recommendations regarding the optimal protein intake per serving for athletes to maximize MPS are mixed and are dependent upon age and recent resistance exercise stimuli. General recommendations are 0.25 g of a high-quality protein per kg of body weight, or an absolute dose of 20–40 g.
- Higher doses (~40 g) are likely needed to maximize MPS responses in elderly individuals.
- Even higher amounts (~70 g) appear to be necessary to promote attenuation of muscle protein breakdown.
- Pacing or spreading these feeding episodes approximately three hours apart has been consistently reported to promote sustained, increased levels of MPS and performance benefits.
Arginine
“NO supplements” containing arginine are consumed by bodybuilders pre-workout in an attempt to increase blood flow to the muscle during exercise, increase protein synthesis, and improve exercise performance. However, there is little scientific evidence to back these claims. Fahs et al. 174 supplemented healthy young men with 7 g arginine or a placebo prior to exercise and observed no significant change in blood flow following exercise. Additionally, Tang et al. 175 supplemented either 10 g arginine or a placebo prior to exercise and found no significant increase in blood flow or protein synthesis following exercise. Moreover, arginine is a non essential amino acid and prior work has established that essential amino acids alone stimulate protein synthesis 176. Based on these findings, it appears that arginine does not significantly increase blood flow or enhance protein synthesis following exercise.
The effects of arginine supplementation on performance are controversial. Approximately one-half of acute and chronic studies on arginine and exercise performance have found significant benefits with arginine supplementation, while the other one-half has found no significant benefits 177. Moreover, Greer et al. 178 found that arginine supplementation significantly reduced muscular endurance by 2–4 repetitions on chin up and push up endurance tests. Based on these results, the authors of a recent review concluded that arginine supplementation had little impact on exercise performance in healthy individuals 179. Although the effects of arginine on blood flow, protein synthesis, and exercise performance require further investigation, dosages commonly consumed by athletes are well below the observed safe level of 20 g/d and do not appear to be harmful 180.
Citrulline malate
Citrulline malate (CitM) has recently become a popular supplement among bodybuilders; however, there has been little scientific research in healthy humans with this compound. CitM is hypothesized to improve performance through three mechanisms: 1) citrulline is important part of the urea cycle and may participate in ammonia clearance, 2) malate is a tricarboxylic acid cycle intermediate that may reduce lactic acid accumulation, and 3) citrulline can be converted to arginine; however, as discussed previously, arginine does not appear to have an ergogenic effect in young healthy athletes so it is unlikely CitM exerts an perfromance enhancing effect through this mechanism 181.
Supplementation with CitM for 15 days has been shown to increase ATP production by 34% during exercise, increase the rate of phosphocreatine recovery after exercise by 20%, and reduce perceptions of fatigue [184]. Moreover, ingestion of 8 g CitM prior to a chest workout significantly increased repetitions performed by approximately 53% and decreased soreness by 40% at 24 and 48 hours post-workout 181. Furthermore, Stoppani et al. 182 in an abstract reported a 4 kg increase in lean mass, 2 kg decrease in body fat percentage, and a 6 kg increase in 10 repetition maximum bench press after consumption of a drink containing 14 g BCAA, glutamine, and CitM during workouts for eight weeks; although, it is not clear to what degree CitM contributed to the outcomes observed. However, not all studies have supported ergogenic effects of CitM. Sureda et al. 183 found no significant difference in race time when either 6 g CitM or a placebo were consumed prior to a 137 km cycling stage. Hickner et al. 184 found that treadmill time to exhaustion was significantly impaired, with the time taken to reach exhaustion occurring on average seven seconds earlier following CitM consumption.
Additionally, the long-term safety of CitM is unknown. Therefore, based on the current literature a decision on the efficacy of CitM cannot be made. Future studies are needed to conclusively determine if CitM is ergogenic and to determine its long term safety.
Glutamine
Glutamine is the most abundant non-essential amino acid in muscle and is commonly consumed as a nutritional supplement. Glutamine supplementation in quantities below 14 g/d appear to be safe in healthy adults 180; however, at present there is little scientific evidence to support the use of glutamine in healthy athletes 150. Acutely, glutamine supplementation has not been shown to significantly improve exercise performance 185, 186, improve buffering capacity 186, help to maintain immune function or reduce muscle soreness after exercise 150. Long-term supplementation studies including glutamine in cocktails along with creatine monohydrate, whey protein, BCAA’s, and/or CitM have shown 1.5 – 2 kg increases in lean mass and 6 kg increase in 10RM bench press strength 182, 187. However, the role of glutamine in these changes is unclear. Only one study 188 has investigated the effects of glutamine supplementation alone in conjunction with a six week strength training program. No significant differences in muscle size, strength, or muscle protein degradation were observed between groups. Although the previous studies do not support the use of glutamine in bodybuilders during contest preparation, it should be noted that glutamine may be beneficial for gastrointestinal health and peptide uptake in stressed populations 189; therefore, it may be beneficial in dieting bodybuilders who represent a stressed population. As a whole, the results of previous studies do not support use of glutamine as an ergogenic supplement; however, future studies are needed to determine the role of glutamine on gastrointestinal health and peptide transport in dieting bodybuilders.
Whey protein
Milk protein is mostly composed of whey protein 20% and casein 80% 190. During cheese manufacturing, whey protein is generated as a by-product of casein precipitation. Whey protein is the most popular protein supplement sold in powder format. It contains valuable food ingredients because of its nutritional value and functional bioactivity. Whey protein contains β-lactoglobulin, α-lactalbumin, immunoglobulins, bovine serum albumin, lactoferrin, lactoperoxidase, phospholipoprotein, bioactive factors, and enzymes in order of abundance 191. The biological components of whey protein and its isolates have been reported to benefit antioxidation 192 and regulation of lipid metabolism 193 and have antifatigue 194 and antidiabetic properties 195.
Whey protein isolates contain enriched essential amino acids, including branched chain amino acids, which the body needs for tissue synthesis, energy, and health. The high leucine content (50%–75% more than other protein sources), one of the branched chain amino acids, in whey protein could explain its ability to stimulate muscle protein synthesis 196 and upregulate mammalian target of rapamycin signaling in high concentration. With whey protein supplementation, resistance exercises can result in muscle adaption and hypertrophy, regardless of the contraction mode (Whey protein hydrolysate augments tendon and muscle hypertrophy independent of resistance exercise contraction mode. Farup J, Rahbek SK, Vendelbo MH, Matzon A, Hindhede J, Bejder A, Ringgard S, Vissing K. Scand J Med Sci Sports. 2014 Oct; 24(5):788-98. https://www.ncbi.nlm.nih.gov/pubmed/23647357/()). Whey protein is marketed as a dietary supplement and as an aid for muscle development with resistance training. Because of its rapid rate of digestion, whey protein provides a rapid source of amino acids that can be taken up by the muscles to repair and rebuild muscular tissue. The use of whey protein to enhance aerobic exercises and swimming training has only been reported in terms of glycogen storage 197, antioxidation 198, and lipid metabolism. A combination of resistant exercise and WP benefitted the lipid profile, especially plasma triglycerides and cholesterol 199. Few reports have shown the beneficial synergistic effects of whey protein and long-term aerobic exercise training on biochemical profiles in specific tissues.
Resistance exercise, eccentric (muscle lengthening), isometric (non-lengthening) and concentric (shortening) contractions cause skeletal muscle damage and generate inflammatory markers (muscle proteins in blood) (The exercise-induced stress response of skeletal muscle, with specific emphasis on humans. Morton JP, Kayani AC, McArdle A, Drust B. Sports Med. 2009; 39(8):643-62. https://www.ncbi.nlm.nih.gov/pubmed/19769414/()). Anabolic interventions with protein hydrolysates and amino acid supplements have been evidenced to expedite the repair. Leucine-derived metabolite β-hydroxy-β-methylbutyrate ingestion has proved beneficial in recovery from the soreness. Resistance exercise (weight-lifting) elevates oxidation products in plasma, perturbs leukocyte redistribution and leukocyte functionality. Whey protein isolate nanoparticles were prepared using ethanol desolvation and their capacity to incorporate ZnCl2 was analysed. The amount of zinc incorporated in the particle suspensions was within the range of daily zinc requirements for healthy adults. Also, the nanoparticles remained stable after 30 days of storage at 22 °C. Cell surface glucose transporter 4 (GLUT 4) is the major glucose transporter isoform expressed in skeletal muscle that determines the rate of muscle glucose transport in the cell membrane, in response to insulin and muscle contraction. Whey protein hydrolysate was evaluated for its ability to translocate GLUT 4 and accumulate them in the membrane thereby augmenting glucose trapping by skeletal muscle. The amino acid l-isoleucine and the peptide l-leucyl-l-isoleucine in the hydrolysate contributed the most 200. The effect of whey supplementation in comparison to casein diet, on the recovery of muscle functional properties such as contractility, extensibility, elasticity and excitability was investigated in rats. The whey protein diet promoted a faster recovery from injury sustained due to isometric as well as concentric exercise in comparison to the casein diet 201. The effect of the supplementation of a beverage with varying doses of leucine or a mixture of branched chain amino acids on myofibrillar protein synthesis after resistance exercise was assessed. Results showed that low-protein (6.25 g) beverage can be as effective as a high-protein dose (25 g) at stimulating myofibrillar protein synthesis rates when supplemented with a high (5 g) leucine content. As leucine comprises 10 % of the total whey amino acid, the latter appears important for augmenting muscle hypertrophy. Health parameters, performance and body composition effects produced by 12 week intake of hydrolysed whey protein were compared in players. Intervention with the hydrolysed whey protein resulted in significant reduction in the muscle damage markers (creatine kinase and lactate dehydrogenase). Muscle mass growth by daily consumption of whey protein was compared with that of soy protein, using a randomized study on subjects undergoing resistance exercise 202. Lean body mass gains were significantly high in whey protein than soy protein group and the remarkable response was correlated with the elevated levels of leucine and faster absorption.
Concerning lean body mass, many studies reported that protein synthesis could be upregulated by the branched chain amino acids of whey protein, especially leucine 203. The combination of daily supplementation with whey protein and resistance exercise training was effective in promoting muscle hypertrophy 204.
Whey protein consumption at intake levels up to 3 g/kg per day had a no-observed-adverse-effect level 205 and the hydrolysate of whey protein at 2 g/kg as a food additive resulted in no adverse effects or mortality 206. In this study, the whey protein dose was 4.1 g/kg, which is equivalent to 20 g of whey protein per 60 kg body weight for humans did not reveal any adverse effects 207.
Based on their concentration and attributes, whey proteins are marketed in various forms such as whey protein concentrate (has fat and lactose along with proteins (29–89 %)), whey protein isolate (90 % protein) and whey protein hydrolysate (partially digested for ease of metabolism and hypo-allergenicity) 208. A broad range of functionality has been assigned to whey protein and its derivatives, such as reduction of oxidative stress, promotion of muscle growth and lean body mass, appetite suppression, hypoglycemia, cardiovascular risk mitigation, phenylketonuria management and protection from ultraviolet (UV) radiation 209. Further, its role in food processing such as emulsifier, texturizer, fat-replacer, encapsulating agent, delivery vehicle and antimicrobial film are being recognized 210.
Protein timing
Thanks to seminal work by pioneering research groups, by the 1990’s it was clear that exercise and macronutrient consumption interact synergistically to provide a net anabolic effect far greater than either feeding or exercise alone. In the absence of feeding, muscle protein balance remains negative in response to an acute bout of resistance exercise. Tipton et al. 211 were one of the first groups to illustrate that an acute feeding of amino acids significantly increases rates of muscle protein synthesis (MPS). Later, Burd et al. 212 indicated that the combination of acute, exhaustive resistance exercise increases the muscle’s anabolic responsiveness to whey protein provision for up to 24 h. In addition to heightened anabolic sensitivity that stems from the combination of resistance exercise and protein/amino acid feeding, the importance of the EAAs with respect to muscle protein growth has also been elucidated. Tipton et al. 213 first indicated that nonessential amino acids were not necessary to stimulate MPS. The study by Borsheim also documented a dose-response outcome characterized by a near doubling of net protein balance in response to a three to six gram dose of the EAAs 47. Building on this work, Tipton et al. 214 reported that EAAs (9–15 g dose) before and after resistance exercise promoted higher net protein accretion, not just 3 or 4 h post exercise but also over a 24-h period 215. These findings formed the theoretical concept of protein timing for resistance exercise that has since been transferred to not only other short-duration, high-intensity activities, but also endurance-based sports and subsequent performance outcomes. The strategic consumption of nutrition, namely protein or various forms of amino acids, in the hours immediately before and during exercise (i.e., peri-workout nutrition) has been shown to maximize muscle repair and optimize strength- and hypertrophy-related adaptations 216. While earlier investigations reported positive effects from consumption of amino acids, it is now clear that intact protein supplements such as egg, whey, casein, beef, soy and even whole milk can evoke an anabolic response that can be similar or greater in magnitude to free form amino acids, assuming ingestion of equal EAA amounts.
For instance, whey protein ingested close to resistance exercise, promotes a higher activation (phosphorylation) of mTOR (a key signaling protein found in myocytes that is linked to the synthesis of muscle proteins) and its downstream mRNA translational signaling proteins (i.e., p70s6 kinase and eIF4BP) that further suggests timed ingestion of protein may favorably promote heightened muscle hypertrophy 217. Moreover, it was found that the increased mTOR signaling corresponded with significantly greater muscle hypertrophy after 10 weeks of training 218. However, the hypertrophic differences between protein consumption and a non-caloric placebo appeared to plateau by week 21, despite a persistently greater activation of this molecular signaling pathway from supplementation. Results from other research groups show that timing of protein near (± 2 h) aerobic and anaerobic exercise training appears to provide a greater activation of the molecular signalling pathways that regulate myofibrillar and mitochondrial protein synthesis as well as glycogen synthesis.
It is widely reported that protein consumption directly after resistance exercise is an effective way to acutely promote a positive muscle protein balance 219, which if repeated over time should translate into a net gain or hypertrophy of muscle. Pennings and colleagues 220 reported an increase in both the delivery and incorporation of dietary proteins into the skeletal muscle of young and older adults when protein was ingested shortly after completion of exercise. These findings and others add to the theoretical basis for consumption of post-protein sooner rather than later after exercise, since post workout MPS rates peak within three hours and remain elevated for an additional 24–72 h. This extended time frame also provides a rationale for both immediate and sustained (i.e., every 3–4 h) feedings to optimize impact. These temporal considerations would also capture the peak elevation in signalling proteins shown to be pivotal for increasing the initiation of translation of muscle proteins, which for the most part appears to peak between 30 and 60 min after exercise. Finally, while some investigations have shown that a rapid increase in amino acids (aminoacidemia) from a protein dose immediately after or surrounding exercise stimulates increased adaptations to resistance training, others examining competitive strength/power athletes reported no advantage from pre/post supplement feedings compared to similar feedings in morning and evening hours. However, these differences may be related to the type of protein used between the studies. The studies showing positive effects of protein timing used milk proteins, whereas the latter study used a collagen based protein supplement.
While a great deal of work has focused on post-exercise protein ingestion, other studies have suggested that pre-exercise and even intra-exercise ingestion may also support favorable changes in MPS and muscle protein breakdown. Initially, Tipton and colleagues 214 directly compared immediate pre-exercise and immediate post-exercise ingestion of a mixture of carbohydrate (35 g) and EAAs (6 g) combination on changes in MPS. They reported that pre-exercise ingestion promoted higher rates of MPS while also demonstrating that nutrient ingestion prior to exercise increased nutrient delivery to a much greater extent than other (immediate or one hour post-exercise) time points. These results were later challenged by Fujita in 2009 221 who employed an identical study design with a different tracer incorporation approach and concluded there was no difference between pre- or post-exercise ingestion. Subsequent work by Tipton 91 also found that similar elevated rates of MPS were achieved when ingesting 20 g of a whey protein isolate immediately before or immediately after resistance exercise.
At this point, whether any particular time of protein ingestion confers any unique advantage over other time points throughout a 24-h day to improve strength and hypertrophy has yet to be adequately investigated. To date, although a substantial amount of literature discusses this concept, a limited number of training studies have assessed whether immediate pre- and post-exercise protein consumption provides unique advantages compared to other time points 222. Each study differed in population, training program, environment and nutrition utilized, with each reporting a different result. What is becoming clear is that the subject population, nutrition habits, dosing protocols on both training and non-training days, energy and macronutrient intake, as well as the exercise bout or training program itself should be carefully considered alongside the results. In particular, the daily amount of protein intake seems to operate as a key consideration because the benefits of protein timing in relation to the peri-workout period seem to be lessened for people who are already ingesting appropriate amounts of protein (e.g. ≥1.6 g/kg/day). A literature review by Aragon and Schoenfeld 223 determined that while compelling evidence exists showing muscle is sensitized to protein ingestion following training, the increased sensitivity to protein ingestion might be greatest in the first five to six hours following exercise. Thus, the importance of timing may be largely dependent on when a pre-workout meal was consumed, the size and composition of that meal and the total daily protein in the diet. In this respect, a pre-exercise meal will provide amino acids during and after exercise and therefore it stands to reason there is less need for immediate post-exercise protein ingestion if a pre-exercise meal is consumed less than five hours before the anticipated completion of a workout. A meta-analysis by Schoenfeld et al. 224 found that consuming protein within one-hour post resistance exercise had a small but significant effect on increasing muscle hypertrophy compared to delaying consumption by at least two hours. However, sub-analysis of these results revealed the effect all but disappeared after controlling for the total intake of protein, indicating that favorable effects were due to unequal protein intake between the experimental and control groups (∼1.7 g/kg versus 1.3 g/kg, respectively) as opposed to temporal aspects of feeding. The authors concluded that total protein intake was the strongest predictor of muscular hypertrophy and that protein timing likely influences hypertrophy to a lesser degree. However, the conclusions from this meta-analysis may be questioned because the majority of the studies analyzed were not protein timing studies but rather protein supplementation studies. In that respect, the meta-analysis provides evidence that protein supplementation (i.e., greater total daily protein intake) may indeed confer an anabolic effect. While a strong rationale remains to support the concept that the hours immediately before or after resistance exercise represents an opportune time to deliver key nutrients that will drive the accretion of fat-free mass and possibly other favorable adaptations, the majority of available literature suggests that other factors may indeed be operating to a similar degree that ultimately impact the observed adaptations. In this respect, a key variable that must be accounted for is the absolute need for energy and protein required to appropriately set the body up to accumulate fat-free mass.
A review by Bosse and Dixon 225 critically summarized the available literature on protein supplementation during resistance exercise and hypothesized that protein intake may need to increase by as much as 59% above baseline levels for significant changes in fat-free mass to occur. Finally, it should be noted that for many athletes, consuming a post- or pre-workout protein-containing meal represents a feeding opportunity with little downside, since there is no benefit from not consuming protein pre- and/or post-exercise. In other words, not consuming protein-containing foods/supplements post-exercise is a strategy that provides no benefit whatsoever. Thus, the most practical recommendation is to have athletes consume a meal during the post-workout (or pre-workout) time period since it may either help or have a neutral effect.
In younger subjects, the ingestion of 20–30 g of any high biological value protein before or after resistance exercise appears to be sufficient to maximally stimulate MPS. More recently, Macnaughton and colleagues 225 reported that 40 g of whey protein ingestion significantly increased the MPS responses compared to a 20 g feeding after an acute bout of whole-body resistance exercise, and that the absolute protein dose may operate as a more important consideration than providing a protein dose that is normalized to lean mass. Free form EAAs, soy, milk, whey, caseinate, and other protein hydrolysates are all capable of activating MPS. However, maximal stimulation of MPS, which results in higher net muscle protein accretion, is the product of the total amount of EAA in circulation as well as the pattern and appearance rate of aminoacidemia that modulates the MPS response. Recent work has clarified that whey protein provides a distinct advantage over other protein sources including soy (considered another fast absorbing protein) and casein (a slower acting protein source) on acute stimulation of MPS. Importantly, an elegant study by West and investigators 226 sought to match the delivery of EAAs in feeding patterns that replicated how whey and casein are digested. The authors reported that a 25 g dose of whey protein that promoted rapid aminoacidemia further enhanced MPS and anabolic signaling when compared to an identical total dose of whey protein when delivered as ten separate 2.5 g doses intended to replicate a slower digesting protein. The advantages of whey protein are important to consider, particularly as all three sources rank similarly in assessments of protein quality. In addition to soy, other plant sources (e.g., pea, rice, hemp, etc.) have garnered interest as potential protein sources to consider. Unfortunately, research that examines the ability of these protein sources to modulate exercise performance and training adaptations is limited at this time. One study conducted by Joy and investigators 125 compared the effect of supplementing a high-dose (48 g/day) of whey or rice protein in experienced resistance-trained subjects during an 8-week resistance training program. The investigators concluded that gains in strength, muscle thickness and body composition were similar between the two protein groups, suggesting that rice protein may be a suitable alternative to whey protein at promoting resistance training adaptations. Furthermore, differences in absorption kinetics, and the subsequent impact on muscle protein metabolism appear to extend beyond the degree of hydrolysis and amino acid profiles. For instance, unlike soy more of the EAAs from whey proteins (hydrolysates and isolates) survive splanchnic uptake and travel to the periphery to activate a higher net gain in muscle 63. Whey proteins (hydrolysates and isolates) appear to be the most extensively researched for pre/post resistance exercise supplementation, possibly because of their higher EAA and leucine content, solubility, and optimal digestion kinetics. These characteristics yield a high concentration of amino acids in the blood (aminoacidemia) that facilitates greater activation of MPS and net muscle protein accretion, in direct comparison to other protein choices. The addition of creatine to whey protein supplementation appears to further augment these adaptations; however, an optimal timing strategy for this combination remains unclear.
The timing of protein-rich meals consumed throughout a day has the potential to influence adaptations to exercise. Using similar methods, other studies over recent decades have established the following:
- MPS increases approximately 30–100% in response to a protein-containing meal to promote a positive net protein balance, and the major contributing factor to this response is the EAA content.
- The anabolic response to feeding is pronounced but transient. During the post-prandial phase (1–4 h after a meal) MPS is elevated, resulting in a positive muscle protein balance. In contrast, MPS rates are lower in a fasted state and muscle protein balance is negative. Protein accretion only occurs in the fed state. The concentration of EAA in the blood (plasma) regulates protein synthesis rates within muscle at rest and post exercise. More recent work has established that protein-carbohydrate supplementation after strenuous endurance exercise stimulates contractile MPS via similar signaling pathways as resistance exercise. Most importantly, and as mentioned initially in this section, muscle appears to be “sensitized” to protein feeding for at least 24 h after exercise. That is, the consumption of a protein-containing meal up to 24 h after a single bout of resistance exercise results in a higher net stimulation of MPS and protein accretion than the same meal consumed after 24 h of inactivity.
- The effect of insulin on MPS is dependent on its ability to increase amino acid availability, which does not occur when insulin is systematically increased (e.g., following feeding). In particular, insulin’s impact on net protein balance seems to operate most powerfully in an anti-catabolic manner on muscle. However, insulin-mediated effects that reduce muscle protein breakdown peaks at low to moderate levels of insulin (~15–30 μIU/mL) that can be achieved by consumption of a 45-g dose of whey protein isolate alone. Taken together, these results seem to indicate that post-workout carbohydrate supplementation offers very little contribution from a muscle development standpoint provided adequate protein is consumed. For example, Staples and colleagues compared the impact of a carbohydrate + protein combination on rates of MPS and reported no further increases in MPS beyond what was seen with protein ingestion alone. Importantly, these results are not to be interpreted to mean that carbohydrate administration offers no potential effect for an athlete engaging in moderate to high volumes of training, but rather that benefits derived from carbohydrate administration appear to more favorably impact aspects of muscle glycogen recovery as opposed to stimulating muscle protein accretion.
Pre-sleep protein intake
Eating before sleep has long been controversial. However, a methodological consideration in the original studies such as the population used, time of feeding, and size of the pre-sleep meal confounds firm conclusions about benefits or drawbacks. Recent work using protein-rich beverages 30-min prior to sleep and two hours after the last meal (dinner) have identified pre-sleep protein consumption/ingestion as advantageous to MPS, muscle recovery, and overall metabolism in both acute and long-term studies. Results from several investigations indicate that 30–40 g of casein protein ingested 30-min prior to sleep 227 or via nasogastric tubing increased overnight MPS in both young and old men, respectively. Likewise, in an acute setting, 30 g of whey protein, 30 g of casein protein, and 33 g of carbohydrate consumed 30-min prior to sleep resulted in an elevated morning resting metabolic rate in young fit men compared to a non-caloric placebo 228. Similarly, although not statistically significant, morning increases in resting metabolic rate were reported in young overweight and/or obese women. Interestingly, Madzima et al. 228 reported that subjects’ respiratory quotient measured during the morning after pre-sleep nutrient intake was unchanged only for the placebo and casein protein trials, while both carbohydrate and whey protein were increased compared to placebo. This infers that casein protein consumed pre-sleep maintains overnight lipolysis and fat oxidation. This finding was further supported by Kinsey et al. 229 using a microdialysis technique to measure interstitial glycerol concentrations overnight from the subcutaneous abdominal adipose tissue, reporting greater fat oxidation following consumption of 30 g of casein compared to a flavor and sensory-matched noncaloric placebo in obese men. Similar to Madzima et al. 228, Kinsey et al. 229 concluded that pre-sleep casein did not blunt overnight lipolysis or fat oxidation. Interestingly, the pre-sleep protein and carbohydrate ingestion resulted in elevated insulin concentrations the next morning and decreased hunger in this overweight population. Of note, it appears that exercise training completely ameliorates any rise in insulin when eating at night before sleep 230, while the combination of pre-sleep protein and exercise has been shown to reduce blood pressure and arterial stiffness in young obese women with prehypertension and hypertension 231. In athletes, evening chocolate milk consumption has also been shown to influence carbohydrate metabolism in the morning, but not running performance. In addition, data supports that exercise performed in the evening augments the overnight MPS response in both younger and older men.
To date, only a few studies involving nighttime protein ingestion have been carried out for longer than four weeks. Snijders et al. 232 randomly assigned young men (average age of 22 years) to consume a protein-centric supplement (27.5 g of casein protein, 15 g of carbohydrate, and 0.1 g of fat) or a noncaloric placebo every night before sleep while also completing a 12-week progressive resistance exercise training program (3 times per week). The group receiving the protein-centric supplement each night before sleep had greater improvements in muscle mass and strength over the 12-week study. Of note, this study was non-nitrogen balanced and the protein group received approximately 1.9 g/kg/day of protein compared to 1.3 g/kg/day in the placebo group. More recently, in a study in which total protein intake was equal, Antonio et al. 233 studied young healthy men and women that supplemented with casein protein (54 g) for 8 weeks either in the morning (any time before 12 pm) or the evening supplementation (90 min or less prior to sleep). They examined the effects on body composition and performance 233. All subjects maintained their usual exercise program. The authors reported no differences in body composition or performance between the morning and evening casein supplementation groups. However, it is worth noting that, although not statistically significant, the morning group added 0.4 kg of fat free mass while the evening protein group added 1.2 kg of fat free mass, even though the habitual diet of the trained subjects in this study consumed 1.7 to 1.9 g/kg/day of protein. Although this finding was not statistically significant, it supports data from Burk et al. 222 indicating that casein-based protein consumed in the morning (10 am) and evening (10:30 pm) was more beneficial for increasing fat-free mass than consuming the protein supplement in the morning (10 am) and afternoon (~3:50 pm). It should be noted that the subjects in the Burk et al. study were resistance training. A retrospective epidemiological study by Buckner et al. 234 using NHANES data (1999–2002) showed that participants consuming 20, 25, or 30 g of protein in the evening had greater leg lean mass compared to subjects consuming protein in the afternoon. Thus, it appears that protein consumption in the evening before sleep might be an underutilized time to take advantage of a protein feeding opportunity that can potentially improve body composition and performance.
Protein ingestion and meal timing
In addition to direct assessments of timed administration of nutrients, other studies have explored questions that center upon the pattern of when certain protein-containing meals are consumed. Paddon-Jones et al. 235 reported a correlation between acute stimulation of MPS via protein consumption and chronic changes in muscle mass. In this study, participants were given an EAA supplement three times a day for 28 days. Results indicated that acute stimulation of MPS provided by the supplement on day 1 resulted in a net gain of ~7.5 g of muscle over a 24-h period 235. When extrapolated over the entire 28-day study, the predicted change in muscle mass corresponded to the actual change in muscle mass (~210 g) measured by dual-energy x-ray absorptiometry (DEXA) 235. While these findings are important, it is vital to highlight that this study incorporated a bed rest model with no acute exercise stimulus while other work by Mitchell et al. 236 reported a lack of correlation between measures of acute MPS and the accretion of skeletal muscle mass.
Interestingly, supplementation with 15 g of EAAs and 30 g of carbohydrate produced a greater anabolic effect (increase in net phenylalanine balance) than the ingestion of a mixed macronutrient meal, despite the fact that both interventions contained a similar dose of EAAs. Most importantly, the consumption of the supplement did not interfere with the normal anabolic response to the meal consumed three hours later. The results of these investigations suggest that protein supplement timing between the regular “three square meals a day” may provide an additive effect on net protein accretion due to a more frequent stimulation of MPS. Areta et al. 237 were the first to examine the anabolic response in human skeletal muscle to various protein feeding strategies for a day after a single bout of resistance exercise. The researchers compared the anabolic responses of three different patterns of ingestion (a total of 80 g of protein) throughout a 12-h recovery period after resistance exercise. Using a group of healthy young adult males, the protein feeding strategies consisted of small pulsed (8 × 10 g), intermediate (4 × 20 g), or bolus (2 × 40 g) administration of whey protein over the 12-h measurement window. Results showed that the intermediate dosing (4 × 20 g) was superior for stimulating MPS for the 12-h experimental period. Specifically, the rates of myofibrillar protein synthesis were optimized throughout the day of recovery by the consumption of 20 g protein every three hours compared to large (2 × 40 g), less frequent servings or smaller but more frequent (8 × 10 g) patterns of protein intake. Previously, the effect of various protein feeding strategies on skeletal MPS during an entire day was unknown. This study provided novel information demonstrating that the regulation of MPS can be modulated by the timing and distribution of protein over 12 h after a single bout of resistance exercise. However, it should be noted that an 80 g dose of protein over a 12-h period is quite low.
The logical next step for researchers is to extend these findings into longitudinal training studies to see if these patterns can significantly affect resistance-training adaptations. Indeed, published studies by Arnal 238 and Tinsley 239 have all made some attempt to examine the impact of adjusting the pattern of protein consumption across the day in combination with various forms of exercise. Collective results from these studies are mixed. Thus, future studies in young adults should be designed to compare a balanced vs. skewed distribution pattern of daily protein intake on the daytime stimulation of MPS (under resting and post-exercise conditions) and training-induced changes in muscle mass, while taking into consideration the established optimal dose of protein contained in a single serving for young adults. Without more conclusive evidence spanning several weeks, it seems pragmatic to recommend the consumption of at least 20-25 g of protein (~0.25 g/kg/meal) with each main meal with no more than 3–4 h between meals.
Key points
- In the absence of feeding and in response to resistance exercise, muscle protein balance remains negative.
- Skeletal muscle is sensitized to the effects of protein and amino acids for up to 24 h after completion of a bout of resistance exercise.
- A protein dose of 20–40 g of protein (10–12 g of EAAs, 1–3 g of leucine) stimulates MPS, which can help to promote a positive nitrogen balance.
- The EAAs are critically needed for achieving maximal rates of MPS making high-quality, protein sources that are rich in EAAs and leucine the preferred sources of protein.
- Studies have suggested that pre-exercise feedings of amino acids in combination with carbohydrate can achieve maximal rates of MPS, but protein and amino acid feedings during this time are not clearly documented to increase exercise performance.
- Ingestion of carbohydrate + protein or EAAs during endurance and resistance exercise can help to maintain a favorable anabolic hormone profile, minimize increases in muscle damage, promote increases in muscle cross-sectional area, and increase time to exhaustion during prolonged running and cycling.
- Post-exercise administration of protein when combined with suboptimal intake of carbohydrates (<1.2 g/kg/day) can heighten muscle glycogen recovery, and may help mitigate changes in muscle damage markers.
- Total protein and calorie intake appears to be the most important consideration when it comes to promoting positive adaptations to resistance training, and the impact of timing strategies (immediately before or immediately after) to heighten these adaptations in non-athletic populations appears to be minimal.
Timing and consumption of protein and/or carbohydrate during workouts
Questions remain about the utility of consuming protein and/or carbohydrate during bodybuilding-oriented training bouts. Since these bouts typically do not resemble endurance bouts lasting 2 hours or more, nutrient consumption during training is not likely to yield any additional performance-enhancing or muscle -sparing benefits if proper pre-workout nutrition is in place. In the exceptional case of resistance training sessions that approach or exceed two hours of exhaustive, continuous work, it might be prudent to employ tactics that maximize endurance capacity while minimizing muscle damage. This would involve approximately 8–15 g protein co-ingested with 30–60 g carbohydrate in a 6-8% solution per hour of training 240. Nutrient timing is an intriguing area of study that focuses on what might clinch the competitive edge. In terms of practical application to resistance training bouts of typical length, Aragon and Schoenfeld 223 recently suggested a protein dose corresponding with 0.4-0.5 g/kg bodyweight consumed at both the pre- and post-exercise periods. However, for objectives relevant to bodybuilding, the current evidence indicates that the global macronutrient composition of the diet is likely the most important nutritional variable related to chronic training adaptations. Figure 1 below provides a continuum of importance with bodybuilding-specific context for nutrient timing.
Figure 1. Continuum of nutrient & supplement timing importance
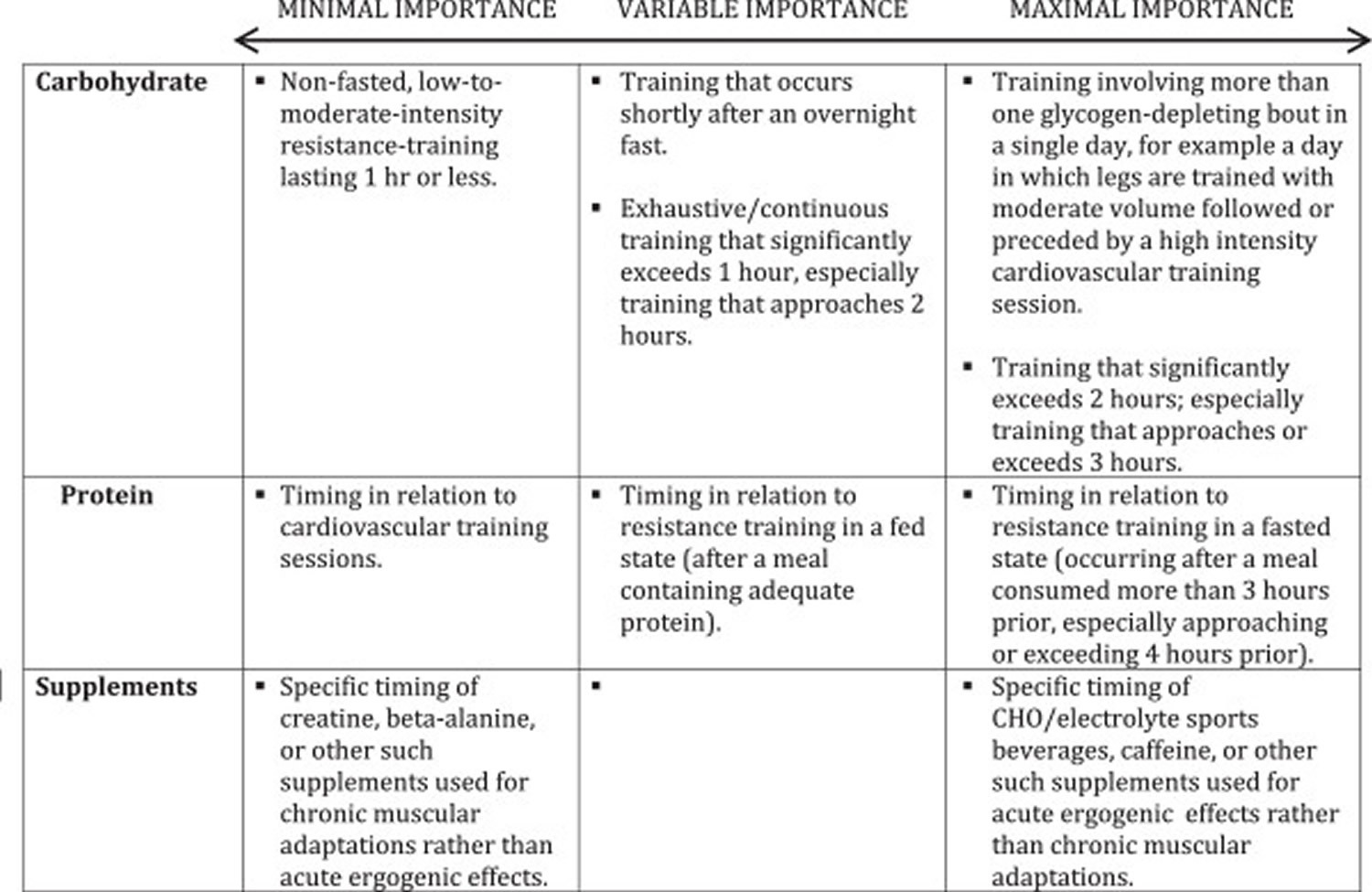
Micronutrients
Several previous studies have observed deficiencies in intakes of micronutrients, such as vitamin D, calcium, zinc, magnesium, and iron, in dieting bodybuilders 242, 243. However, it should be noted that these studies were all published nearly 2 decades ago and that micronutrient deficiencies likely occurred due to elimination of foods or food groups and monotony of food selection 244, 243. Therefore, future studies are needed to determine if these deficiencies would present while eating a variety of foods and using the contest preparation approach described herein. Although the current prevalence of micronutrient deficiencies in competitive bodybuilders is unknown, based on the previous literature, a low-dose micronutrient supplement may be beneficial for natural bodybuilders during contest preparation; however, future studies are needed to verify this recommendation.
Caffeine
Caffeine is perhaps the most common pre-workout stimulant consumed by bodybuilders. Numerous studies support the use of caffeine to improve performance during endurance training 245, 246, sprinting 247, 248, and strength training 249, 250, 251. However, not all studies support use of caffeine to improve performance in strength training 252, 253. It should be noted that many of the studies that found increases in strength training performance supplemented with larger (5–6 mg/kg) dosages of caffeine. However, this dosage of caffeine is at the end of dosages that are considered safe (6 mg/kg/day) 254. Additionally, it appears that regular consumption of caffeine may result in a reduction of ergogenic effects 255. Therefore, it appears that 5–6 mg/kg caffeine taken prior to exercise is effective in improving exercise performance; however, caffeine use may need to be cycled in order for athletes to obtain the maximum performance enhancing effect.
Beta-alanine
Beta-alanine (BA) is becoming an increasingly popular supplement among bodybuilders. Once consumed, BA enters the circulation and is up-taken by skeletal muscle where it is used to synthesize carnosine, a pH buffer in muscle that is particularly important during anaerobic exercise such as sprinting or weightlifting 256. Indeed, consumption of 6.4 g BA daily for four weeks has been shown to increase muscle carnosine levels by 64.2% 257. Moreover, supplementation with BA for 4–10 weeks has been shown to increase knee extension torque by up to 6%, improve workload and time to fatigue during high intensity cardio, improve muscle resistance to fatigue during strength training, increase lean mass by approximately 1 kg and significantly reduce perceptions of fatigue. Additionally, the combination of BA and CM may increase performance of high intensity endurance exercise and has been shown to increase lean mass and decrease body fat percentage more than CM alone. However, not all studies have shown improvements in performance with BA supplementation. To clarify these discrepancies, Hobson et al. 258 conducted a meta-analysis of 15 studies on BA supplementation and concluded that BA significantly increased exercise capacity and improved exercise performance on 60-240 seconds and >240 seconds exercise bouts.
Although BA appears to improve exercise performance, the long-term safety of BA has only been partially explored. Currently, the only known side effect of BA is unpleasant symptoms of parasthesia reported after consumption of large dosages; however, this can be minimized through consumption of smaller dosages throughout the day 257. While BA appears to be relatively safe in the short-term, the long-term safety is unknown. In cats, an addition of 5 percent BA to drinking water for 20 weeks has been shown to deplete taurine and result in damage to the brain; however, taurine is an essential amino acid for cats but not for humans and it is unknown if the smaller dosages consumed by humans could result in similar effects 259. BA may increase exercise performance and increase lean mass in bodybuilders and currently appears to be safe; however, studies are needed to determine the long-term safety of BA consumption.
Beta-hydroxy-beta-methylbutyrate
Beta-hydroxy-beta-methylbutyrate (HMB) is a metabolite of the amino acid leucine that has been shown to decrease muscle protein catabolism and increase muscle protein synthesis 260, 261. The safety of HMB supplementation has been widely studied and no adverse effects on liver enzymes, kidney function, cholesterol, white blood cells, hemoglobin, or blood glucose have been observed 262, 263. Furthermore, two meta-analyses on HMB supplementation have concluded that HMB is safe and does not result in any major side effects 262. HMB may actually decrease blood pressure, total and LDL cholesterol, especially in hypercholesterolemic individuals.
HMB is particularly effective in catabolic populations such as the elderly and patients with chronic disease 264. However, studies on the effectiveness of HMB in trained, non-calorically restricted populations have been mixed. Reasons for discrepancies in the results of HMB supplementation studies in healthy populations may be due to many factors including clustering of data in these meta-analysis to include many studies from similar groups, poorly designed, non-periodized training protocols, small sample sizes, and lack of specificity between training and testing conditions 265. However, as a whole HMB appears to be effective in a majority of studies with longer-duration, more intense, periodized training protocols and may be beneficial to bodybuilders, particularly during planned over-reaching phases of training 266. While the authors hypothesize that HMB may be effective in periods of increased catabolism, such as during contest preparation, the efficacy of HMB on maintenance of lean mass in dieting athletes has not been investigated in a long-term study. Therefore, future studies are needed to determine the effectiveness of HMB during caloric restriction in healthy, lean, trained athletes.
Creatine
Creatine is a naturally-occurring amino acid (protein building block) that’s found in meat and fish, and also made by the human body predominately in the liver, kidneys, and to a lesser extent in the pancreas 267. Creatine is produced in your body at an amount of about 1 g/day 268. The remainder of the creatine available to the body is obtained through the diet at about 1 g/day for an omnivorous diet 268. 95% of the bodies creatine stores are found in the skeletal muscle and the remaining 5% is distributed in the brain, liver, kidney, and testes 269.
Creatine is chemically known as a non-protein nitrogen; a compound which contains nitrogen but is not a protein per se 270. It is synthesized in the liver and pancreas from the amino acids arginine, glycine, and methionine 270, 271. Approximately 95% of the body’s creatine is stored in skeletal muscle. Additionally, small amounts of creatine are also found in the brain and testes 272, 273. About two thirds of the creatine found in skeletal muscle is stored as phosphocreatine (PCr) while the remaining amount of creatine is stored as free creatine 272. The total creatine pool (PCr + free creatine) in skeletal muscle averages about 120-140 grams for a 70 kg individual 274, depending on the skeletal muscle fiber type 269 and quantity of muscle mass 275. However, the average human has the capacity to store up to 160 grams of creatine under certain conditions 276, 277. The body breaks down about 1 – 2% of the creatine pool per day (about 1–2 grams/day) into creatinine in the skeletal muscle 270. The body production and dietary intake matches the rate of creatinine production from the degradation of phosphocreatine and creatine at 2.6% and 1.1%/d respectively. In general, oral creatine supplementation leads to an increase of creatine levels within the body. Creatine can be cleared from the blood by saturation into various organs and cells or by renal filtration then excreted in urine 269.
Creatine stores can be replenished by obtaining creatine in the diet or through endogenous synthesis of creatine from glycine, arginine, and methionine 278, 279. Dietary sources of creatine include meats and fish. Large amounts of fish and meat must be consumed in order to obtain gram quantities of creatine. Whereas dietary supplementation of creatine provides an inexpensive and efficient means of increasing dietary availability of creatine without excessive fat and/or protein intake.
Three amino acids (glycine, arginine and methionine) and three enzymes (L-arginine:glycine amidinotransferase, guanidinoacetate methyltransferase and methionine adenosyltransferase) are required for creatine synthesis. The impact creatine synthesis has on glycine metabolism in adults is low, however the demand is more appreciable on the metabolism of arginine and methionine 275.
As creatine is predominately present in the diet from meats, vegetarians have lower resting creatine concentrations 280. It is converted into creatine phosphate or phosphocreatine and stored in the muscles, where it is used for energy 267. During high-intensity, short-duration exercise, such as lifting weights or sprinting, phosphocreatine is converted into ATP, a major source of energy within the human body.
Biochemically speaking, the energy supplied to rephosphorylate adenosine diphosphate (ADP) to adenosine triphosphate (ATP) during and following intense exercise is largely dependent on the amount of phosphocreatine (PCr) stored in the muscle 281, 276. As phosphocreatine stores become depleted during intense exercise, energy availability diminishes due to the inability to resynthesize ATP at the rate required to sustained high-intensity exercise 281, 276. Consequently, the ability to maintain maximal-effort exercise declines. The availability of phosphocreatine in the muscle may significantly influence the amount of energy generated during brief periods of high-intensity exercise. Furthermore, it has been hypothesized that increasing muscle creatine content, via creatine supplementation, may increase the availability of phosphocreatine allowing for an accelerated rate of resynthesis of ATP during and following high-intensity, short-duration exercise 281, 276. Theoretically, creatine supplementation during training may lead to greater training adaptations due to an enhanced quality and volume of work performed. In terms of potential medical applications, creatine is intimately involved in a number of metabolic pathways. For this reason, medical researchers have been investigating the potential therapeutic role of creatine supplementation in a variety of patient populations.
Creatine supplements are popular among body builders and competitive athletes. As an oral supplement, the most widely used and researched form is creatine monohydrate 268. When orally ingested, creatine monohydrate has shown to improve exercise performance and increase fat free mass 282, 283, 284, 285, 286. The attraction of creatine is that it may increase lean muscle mass and enhance athletic performance, particularly during high-intensity, short-duration sports (like high jumping and weight lifting).
However, not all human studies show that creatine improves athletic performance 267. Nor does every person seem to respond the same way to creatine supplements. For example, people who tend to have naturally high stores of creatine in their muscles don’t get an energy-boosting effect from extra creatine. Preliminary clinical studies also suggest that creatine’s ability to increase muscle mass and strength may help fight muscle weakness associated with illnesses, such as heart failure and muscular dystrophy.
Creatine supplementation Responders vs. Non-responders
Syrotuik and Bell 287 investigated the physical characteristics of responder and non-responder subjects to creatine supplementation in recreationally resistance trained men with no history of creatine monohydrate usage. The supplement group was asked to ingest a loading dosage of 0.3 g/kg/d for 5 days. The physiological characteristics of responders were classified using Greenhaff et al 288 criterion of >20 mmol/kg dry weight increase in total intramuscular creatine and phosphocreatine and non responders as <10 mmol/kg dry weight increase, a third group labeled quasi responders were also used to classify participants who fell in between the previously mentioned groups (10-20 mmol/kg dry weight). Overall, the supplemented group showed a mean increase in total resting muscle creatine and phosphocreatine of 14.5% (from 111.12 ± 8.87 mmol/kg dry weight to 127.30 ± 9.69 mmol/kg dry weight) whilst the placebo group remained relatively unaffected (from 115.70 ± 14.99 mmol/kg dry weight to 111.74 ± 12.95 mmol/kg dry weight). However when looking at individual cases from the creatine group the results showed a variance in response. From the 11 males in the supplemented group, 3 participants were responders (mean increase of 29.5 mmol/kg dry weight or 27%), 5 quasi responders (mean increase of 14.9 mmol/kg dry weight or 13.6%) and 3 non-responders (mean increase of 5.1 mmol/kg dry weight or 4.8%). Using muscle biopsies of the vastus lateralis, a descending trend for groups and mean percentage fiber type was observed. Responders showed the greatest percentage of type II fibers followed by quasi responders and non-responders. The responder and quasi responder groups had an initial larger cross sectional area for type I, type IIa and type IIx fibers. The responder group also had the greatest mean increase in the cross sectional area of all the muscle fiber types measured (type I, type IIa and type IIx increased 320, 971 and 840 μm2 respectively) and non-responders the least (type I, type IIa and type IIx increased 60, 46 and 78 μm2 respectively). There was evidence of a descending trend for responders to have the highest percentage of type II fibers; furthermore, responders and quasi responders possessed the largest initial cross sectional area of type I, IIa and IIx fibers. Responders were seen to have the lowest initial levels of creatine and phosphocreatine. This has also been observed in a previous study 289 which found that subjects whose creatine levels were around 150 mmol/Kg dry mass did not have any increments in their creatine saturation due to creatine supplementation, neither did they experience any increases of creatine uptake, phosphocreatine resynthesis and performance. This would indicate a limit maximum size of the creatine pool.
In summary responders are those individuals with a lower initial level of total muscle creatine content, greater population of type II fibers and possess higher potential to improve performance in response to creatine supplementation.
Effects of creatine supplementation on Muscle Mass
Cribb et al (2007) 290 observed greater improvements on 1RM, lean body mass, fiber cross sectional area and contractile protein in trained young males when resistance training was combined with a multi-nutrient supplement containing 0.1 g/kg/d of creatine, 1.5 g/kg/d of protein and carbohydrate compared with protein alone or a protein carbohydrate supplement without the creatine. These findings were novel because at the time no other research had noted such improvements in body composition at the cellular and sub cellular level in resistance trained participants supplementing with creatine. The amount of creatine consumed in the study by Cribb et al 290 was greater than the amount typically reported in previous studies (a loading dose of around 20 g/d followed by a maintenance dose of 3-5 g/d is generally equivalent to approximately 0.3 g/kg/d and 0.03 g/kg/d respectively) and the length of the supplementation period or absence of resistance exercise may explain the observed transcriptional level changes that were absent in previous studies 291, 292.
Deldicque et al 293 found a 250%, 45% and 70% increase for collagen mRNA, glucose transporter 4 (GLUT4) and Myosin heavy chain IIA, respectively after 5 days creatine loading protocol (21 g/d). The authors speculated that creatine in addition to a single bout of resistance training can favor an anabolic environment by inducing changes in gene expression after only 5 days of supplementation.
When creatine supplementation is combined with heavy resistance training, muscle insulin like growth factor (IGF-1) concentration has been shown to increase. Burke et al 294 examined the effects of an 8 week heavy resistance training protocol combined with a 7 day creatine loading protocol (0.25 g/d/kg lean body mass) followed by a 49 day maintenance phase (0.06 g/kg lean mass) in a group of vegetarian and non-vegetarian, novice, resistance trained men and women. Compared to placebo, creatine groups produced greater increments in IGF-1 (78% Vs 55%) and body mass (2.2 Vs 0.6 kg). Additionally, vegetarians within the supplemented group had the largest increase of lean mass compared to non vegetarian (2.4 and 1.9 kg respectively). Changes in lean mass were positively correlated to the modifications in intramuscular total creatine stores which were also correlated with the modified levels of intramuscular IGF-1. The authors suggested that the rise in muscle IGF-1 content in the creatine group could be due to the higher metabolic demand created by a more intensely performed training session. These amplifying effects could be caused by the increased total creatine store in working muscles. Even though vegetarians had a greater increase in high energy phosphate content, the IGF-1 levels were similar to the amount observed in the non vegetarian groups. These findings do not support the observed correlation pattern by which a low essential amino acid content of a typical vegetarian diet should reduce IGF-1 production 295. According to authors opinions it is possible that the addition of creatine and subsequent increase in total creatine and phosphocreatine storage might have directly or indirectly stimulated production of muscle IGF-I and muscle protein synthesis, leading to an increased muscle hypertrophy 294.
Effects of creatine ingestion to improve recovery from injury, muscle damage and oxidative stress induced by exercise
Creatine supplementation may also be of benefit to injured athletes. Op’t Eijnde et al 296 noted that the expected decline in GLUT4 content after being observed during a immobilization period can be offset by a common loading creatine (20g/d) supplementation protocol. In addition, combining creatine monohydrate 15g/d for 3 weeks following 5 g/d for the following 7 weeks positively enhances GLUT4 content, glycogen, and total muscle creatine storage 296.
Bassit et al (Bassit RA, Pinheiro CH, Vitzel KF, Sproesser AJ, Silveira LR, Curi R. Effect of short-term creatine supplementation on markers of skeletal muscle damage after strenuous contractile activity. Eur J Appl Physiol. 2010;108:945–955. doi: 10.1007/s00421-009-1305-1. https://www.ncbi.nlm.nih.gov/pubmed/19956970()) observed a decrease in several markers of muscle damage (creatine kinase, lactate dehydrogenase, aldolase, glutamic oxaloacetic acid transaminase and glutamic pyruvic acid transaminase) in 4 athletes after an iron man competition who supplemented with 20 g/d plus 50 g maltodextrin during a 5 day period prior to the competition.
Cooke et al 297 observed positive effects of a prior (0.3 g/d kg body weight) loading and a post maintenance protocol (0.1 g/d kg body weight) to attenuate the loss of strength and muscle damage after an acute supramaximal (3 set x 10 rep with 120% 1RM) eccentric resistance training session in young males. The authors speculate that creatine ingestion prior to exercise may enhance calcium buffering capacity of the muscle and reduce calcium-activated proteases which in turn minimize sarcolemma and further influxes of calcium into the muscle. In addition creatine ingestion post exercise would enhance regenerative responses, favoring a more anabolic environment to avoid severe muscle damage and improve the recovery process. In addition, in vitro studies have demonstrated the antioxidant effects of creatine to remove superoxide anion radicals and peroxinitrite radicals 298. This antioxidant effect of creatine has been associated with the presence of Arginine in its molecule. Arginine is also a substrate for nitric oxide synthesis and can increase the production of nitric oxide which has higher vasodilatation properties, and acts as a free radical that modulates metabolism, contractibility and glucose uptake in skeletal muscle. Other amino acids contained in the creatine molecule such as glycine and methinine may be especially susceptible to free radical oxidation because of sulfhydryl groups 298. A more recent in vitro study showed that creatine exerts direct antioxidant activity via a scavenging mechanism in oxidatively injured cultured mammalian cells 299. In a recent in vivo study Rhaini et al 300 showed a positive effect of 7 days of creatine supplementation (4 x 5 g CM 20 g total) on 27 recreational resistance trained males to attenuate the oxidation of DNA and lipid peroxidation after a strenuous resistance training protocol.
Collectively the above investigations indicate that creatine supplementation can be an effective strategy to maintain total creatine pool during a rehabilitation period after injury as well as to attenuate muscle damage induced by a prolonged endurance training session. In addition, it seems that creatine can act as an effective antioxidant agent after more intense resistance training sessions 268.
Effects of creatine supplementation on glycogen stores
It is suggested 301, 302 that another mechanism for the effect of creatine could be enhanced muscle glycogen accumulation and GLUT4 expression, when creatine supplementation is combined with a glycogen depleting exercise. Whereas it has been observed 303 that creatine supplementation alone does not enhance muscle glycogen storage. Hickner et al 304 observed positive effects of creatine supplementation for enhancing initial and maintaining a higher level of muscle glycogen during 2 hours of cycling. In general, it is accepted that glycogen depleting exercises, such as high intensity or long duration exercise should combine high carbohydrate diets with creatine supplementation to achieve heightened muscle glycogen stores 296.
Effects of creatine supplementation on predominantly aerobic exercise
Although creatine supplementation has been shown to be more effective on predominantly anaerobic intermittent exercise, there is some evidence of its positive effects on endurance activities. Branch 305 highlights that endurance activities lasting more than 150 seconds rely on oxidative phosphorylation as primary energy system supplier. From this meta analysis 305, it would appear that the ergogenic potential for creatine supplementation on predominantly aerobic endurance exercise diminishes as the duration of the activity increases over 150s. However it is suggested that creatine supplementation may cause a change in substrate utilization during aerobic activity possibly leading to an increase in steady state endurance performance.
Chwalbinska-Monteta 306 observed a significant decrease in blood lactate accumulation when exercising at lower intensities as well as an increase in lactate threshold in elite male endurance rowers after consuming a short loading (5 days 20 g/d) creatine monohydrate protocol. However, the effects of creatine supplementation on endurance performance have been questioned by some studies. Graef et al 307 examined the effects of four weeks of creatine citrate supplementation and high-intensity interval training on cardio respiratory fitness. A greater increase of the ventilatory threshold was observed in the creatine group respect to placebo; however, oxygen consumption showed no significant differences between the groups. The total work presented no interaction and no main effect for time for any of the groups. Thompson et al 308 reported no effects of a 6 week 2 g creatine monohydrate/day in aerobic and anaerobic endurance performance in female swimmers.
Documented effects of creatine supplementation on physical performance
The majority of studies focusing on creatine supplementation report an increase in the body’s’ creatine pool 304, 301, 289. There is a positive relationship between muscle creatine uptake and exercise performance 289. Volek et al. 309 observed a significant increase in strength performance after 12 weeks creatine supplementation with a concurrent periodized heavy resistance training protocol. The creatine supplementation protocol consisted of a weeklong loading period of 25 g/day followed by a 5 gram maintenance dose for the remainder of the training. These positive effects were attributed to an increased total creatine pool resulting in more rapid adenosine triphosphate (ATP) regeneration between resistance training sets allowing athletes to maintain a higher training intensity and improve the quality of the workouts along the entire training period.
It is regularly reported that creatine supplementation, when combined with heavy resistance training leads to enhanced physical performance, fat free mass, and muscle morphology 310, 311, 312. A 2003 meta analysis 285 showed individuals ingesting creatine, combined with resistance training, obtain on average +8% and +14% more performance on maximum (1RM) or endurance strength (maximal repetitions at a given percent of 1RM) respectively than the placebo groups. However, contradicting studies have reported no effects of creatine supplementation on strength performance. Jakobi et al 313 found no effects of a short term creatine loading protocol upon isometric elbow flexion force, muscle activation, and recovery process. However, this study did not clearly state if creatine supplementation was administered concurrent with resistance training. Bemben et al 314 have shown no additional benefits of creatine alone or combined with whey protein for improving strength and muscle mass after a progressive 14 weeks (3 days per week) resistance training program in older men. These conflicting results can be explained by the possibility that the supplemented groups were formed by a greater amount of non-responders or even because creatine supplementation was administered on the training days only (3 times a week). This strategy has not been adequately tested as effective in middle aged and older men for maintaining post loading elevated creatine stores 282.
A quantitative, comprehensive scientific summary and view of knowledge up to 2007 on the effects of creatine supplementation in athletes and active people was published in a 100 citation review position paper by the International Society of Sports Nutrition 282. More recent literature has provided greater insight into the anabolic/performance enhancing mechanisms of creatine supplementation 304, 315 suggesting that these effects may be due to satellite cell proliferation, myogenic transcription factors and insulin-like growth factor-1 signalling 301. Saremi et al 316 reported a change in myogenic transcription factors when creatine supplementation and resistance training are combined in young healthy males. It was found that serum levels of myostatin, a muscle growth inhibitor, were decreased in the creatine group.
Collectively, in spite of a few controversial results, it seems that creatine supplementation combined with resistance training would amplify performance enhancement on maximum and endurance strength as well muscle hypertrophy 268.
Effects of creatine supplementation on predominantly anaerobic exercise
Creatine has demonstrated neuromuscular performance enhancing properties on short duration, predominantly anaerobic, intermittent exercises. Bazzucch et al 317 observed enhanced neuromuscular function of the elbow flexors in both electrically induced and voluntary contractions but not on endurance performance after 4 loading doses of 5 g creatine plus 15 g maltodextrin for 5/d in young, moderately trained men. Creatine supplementation may facilitate the reuptake of Ca2+ into the sacroplasmic reticulum by the action of the Ca2+ adenosine triphosphatase pump, which could enable force to be produced more rapidly through the faster detachment of the actomyosin bridges.
A previous meta-analysis 305 reported an overall creatine supplementation effect size (ES) of 0.24 ± 0.02 for activities lasting ≤30 s. (primarily using the ATP- phosphocreatine energy system). For this short high-intensity exercise, creatine supplementation resulted in a 7.5 ± 0.7% increase from base line which was greater than the 4.3 ± 0.6% improvement observed for placebo groups. When looking at the individual selected measures for anaerobic performance the greatest effect of creatine supplementation was observed on the number of repetitions which showed an ES of 0.64 ± 0.18. Furthermore, an increase from base line of 45.4 ± 7.2% compared to 22.9 ± 7.3% for the placebo group was observed. The second greatest ES was on the weight lifted at 0.51 ± 0.16 with an increase from base line of 13.4 ± 2.7% for the placebo group and 24.7 ± 3.9% for the creatine group. Other measures improved by creatine with a mean ES greater than 0 were for the amount of work accomplished, weight lifted, time, force production, cycle ergometer revolutions/min and power. The possible effect of creatine supplementation on multiple high intensity short duration bouts (<30 s) have shown an ES not statistically significant from 0. This would indicate that creatine supplementation might be useful to attenuate fatigue symptoms over multiple bouts of high-intensity, short duration exercise. The ES of creatine on anaerobic endurance exercise (>30 – 150s), primarily using the anaerobic glycolysis energy system, was 0.19 ± 0.05 with an improvement from baseline of 4.9 ± 1.5 % for creatine and -2.0 ± 0.6% for the placebo. The specific aspects of anaerobic endurance performance improved by creatine supplementation were work and power, both of which had a mean ES greater than 0. From the findings of this previous meta-analysis 305 it would appear that creatine supplementation has the most pronounced effect on short duration (<30s) high intensity intermittent exercises.
Effects of creatine supplementation on predominantly aerobic exercise
Although creatine supplementation has been shown to be more effective on predominantly anaerobic intermittent exercise, there is some evidence of its positive effects on endurance activities. Branch 305 highlights that endurance activities lasting more than 150 seconds rely on oxidative phosphorylation as primary energy system supplier. From this meta analysis 305, it would appear that the ergogenic potential for creatine supplementation on predominantly aerobic endurance exercise diminishes as the duration of the activity increases over 150s. However it is suggested that creatine supplementation may cause a change in substrate utilization during aerobic activity possibly leading to an increase in steady state endurance performance.
Chwalbinska-Monteta 306 observed a significant decrease in blood lactate accumulation when exercising at lower intensities as well as an increase in lactate threshold in elite male endurance rowers after consuming a short loading (5 days 20 g/d) CM protocol. However, the effects of creatine supplementation on endurance performance have been questioned by some studies. Graef et al 307 examined the effects of four weeks of creatine citrate supplementation and high-intensity interval training on cardio respiratory fitness. A greater increase of the ventilatory threshold was observed in the creatine group respect to placebo; however, oxygen consumption showed no significant differences between the groups. The total work presented no interaction and no main effect for time for any of the groups. Thompson et al 308 reported no effects of a 6 week 2 g CM/d in aerobic and anaerobic endurance performance in female swimmers. In addition, of the concern related to the dosage used in these studies, it could be possible that the potential benefits of creatine supplementation on endurance performance were more related to effects of anaerobic threshold localization.
Effects of creatine supplementation on range of motion
Sculthorpe et al 318 has shown that a 5 day (25g/d) loading protocol of creatine supplementation followed by a further 3 days of 5 g/d negatively influence both active ankle dorsiflexion and shoulder abduction and extension range of movement (ROM) in young men. There are two possible theories to explain these effects: 1) Creatine supplementation increases intracellular water content resulting in increased muscle stiffness and resistance to stretch; 2) Neural outflow from the muscle spindles is affected due to an increased volume of the muscle cell. The authors highlight that the active ROM measures were taken immediately after the loading phase and the reduced active ROM may not be seen after several weeks of maintenance phase 318. Hile et al 319 observed an increase in compartment pressure in the anterior compartment of the lower leg, which may also have been responsible for a reduced active ROM.
Creatine in combination with other supplements
Although creatine can be bought commercially as a standalone product it is often found in combination with other nutrients. A prime example is the combination of creatine with carbohydrate or protein and carbohydrate for augmenting creatine muscle retention 282 mediated through an insulin response from the pancreas 320. Steenge et al 321 found that body creatine retention of 5 g CM was increased by 25% with the addition of 50 g of protein and 47 g of carbohydrate or 96 g carbohydrate when compared to a placebo treatment of 5 g carbohydrate. The addition of 10g of creatine to 75 g of dextrose, 2 g of taurine, vitamins and minerals, induced a change in cellular osmolarity which in addition to the expected increase in body mass, seems to produce an up regulation of large scale gene expression (mRNA content of genes and protein content of kinases involved in osmosensing and signal transduction, cytoskeleton remodelling, protein and glycogen synthesis regulation, satellite cell proliferation and differentiation, DNA replication and repair, RNA transcription control, and cell survival) 315. Similar findings have also been reported for creatine monohydrate supplementation alone when combined with resistance training 322.
A commercially available pre-workout formula comprised of 2.05 g of caffeine, taurine and glucuronolactone, 7.9 g of L-leucine, L-valine, L-arginine and L-glutamine, 5 g of di-creatine citrate and 2.5 g of β-alanine mixed with 500 ml of water taken 10 minutes prior to exercise has been shown to enhance time to exhaustion during moderate intensity endurance exercise and to increase feelings of focus, energy and reduce subjective feelings of fatigue before and during endurance exercise due to a synergistic effect of the before mentioned ingredients 323. The role of creatine in this formulation is to provide a neuroprotective function by enhancing the energy metabolism in the brain tissue, promoting antioxidant activities, improving cerebral vasculation and protecting the brain from hyperosmotic shock by acting as a brain cell osmolyte. Creatine can provide other neuroprotective benefits through stabilisation of mitochondrial membranes, stimulation of glutamate uptake into synaptic vesicles and balance of intracellular calcium homeostasis 323.
- Jäger R, Kerksick CM, Campbell BI, et al. International Society of Sports Nutrition Position Stand: protein and exercise. Journal of the International Society of Sports Nutrition. 2017;14:20. doi:10.1186/s12970-017-0177-8. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5477153/[↩][↩][↩]
- The effect of resistance training combined with timed ingestion of protein on muscle fiber size and muscle strength. Andersen LL, Tufekovic G, Zebis MK, Crameri RM, Verlaan G, Kjaer M, Suetta C, Magnusson P, Aagaard P. Metabolism. 2005 Feb; 54(2):151-6. https://www.ncbi.nlm.nih.gov/pubmed/15690307/[↩][↩]
- Effects of resistance exercise and fortified milk on skeletal muscle mass, muscle size, and functional performance in middle-aged and older men: an 18-mo randomized controlled trial. Kukuljan S, Nowson CA, Sanders K, Daly RM. J Appl Physiol (1985). 2009 Dec; 107(6):1864-73. https://www.ncbi.nlm.nih.gov/pubmed/19850735/[↩]
- Effects of whey isolate, creatine, and resistance training on muscle hypertrophy. Cribb PJ, Williams AD, Stathis CG, Carey MF, Hayes A. Med Sci Sports Exerc. 2007 Feb; 39(2):298-307. https://www.ncbi.nlm.nih.gov/pubmed/17277594/[↩]
- Body composition and strength changes in women with milk and resistance exercise. Josse AR, Tang JE, Tarnopolsky MA, Phillips SM. Med Sci Sports Exerc. 2010 Jun; 42(6):1122-30. https://www.ncbi.nlm.nih.gov/pubmed/19997019/[↩]
- Control of muscle protein synthesis as a result of contractile activity and amino acid availability: implications for protein requirements. Rennie MJ. Int J Sport Nutr Exerc Metab. 2001 Dec; 11 Suppl():S170-6. https://www.ncbi.nlm.nih.gov/pubmed/11915917/[↩]
- Protein supplementation augments the adaptive response of skeletal muscle to resistance-type exercise training: a meta-analysis. Cermak NM, Res PT, de Groot LC, Saris WH, van Loon LJ. Am J Clin Nutr. 2012 Dec; 96(6):1454-64. https://www.ncbi.nlm.nih.gov/pubmed/23134885/[↩]
- The effects of protein supplements on muscle mass, strength, and aerobic and anaerobic power in healthy adults: a systematic review. Pasiakos SM, McLellan TM, Lieberman HR. Sports Med. 2015 Jan; 45(1):111-31. https://www.ncbi.nlm.nih.gov/pubmed/25169440/[↩]
- The science of muscle hypertrophy: making dietary protein count. Phillips SM. Proc Nutr Soc. 2011 Feb; 70(1):100-3. https://www.ncbi.nlm.nih.gov/pubmed/21092368/[↩]
- Dietary protein for muscle hypertrophy. Tipton KD, Phillips SM. Nestle Nutr Inst Workshop Ser. 2013; 76():73-84. https://www.ncbi.nlm.nih.gov/pubmed/23899756/[↩]
- Dietary protein and exercise have additive effects on body composition during weight loss in adult women. Layman DK, Evans E, Baum JI, Seyler J, Erickson DJ, Boileau RA. J Nutr. 2005 Aug; 135(8):1903-10. https://www.ncbi.nlm.nih.gov/pubmed/16046715/[↩][↩]
- A structured diet and exercise program promotes favorable changes in weight loss, body composition, and weight maintenance. Kreider RB, Serra M, Beavers KM, Moreillon J, Kresta JY, Byrd M, Oliver JM, Gutierrez J, Hudson G, Deike E, Shelmadine B, Leeke P, Rasmussen C, Greenwood M, Cooke MB, Kerksick C, Campbell JK, Beiseigel J, Jonnalagadda SS. J Am Diet Assoc. 2011 Jun; 111(6):828-43. https://www.ncbi.nlm.nih.gov/pubmed/21616195/[↩]
- Effects of high-protein diets on fat-free mass and muscle protein synthesis following weight loss: a randomized controlled trial. Pasiakos SM, Cao JJ, Margolis LM, Sauter ER, Whigham LD, McClung JP, Rood JC, Carbone JW, Combs GF Jr, Young AJ. FASEB J. 2013 Sep; 27(9):3837-47. https://www.ncbi.nlm.nih.gov/pubmed/23739654/[↩]
- Andersen LL, Tufekovic G, Zebis MK, Crameri RM, Verlaan G, Kjaer M, et al. The effect of resistance training combined with timed ingestion of protein on muscle fiber size and muscle strength. Metab Clin Exp. 2005;54:151–6. https://www.ncbi.nlm.nih.gov/pubmed/15690307[↩]
- Herda AA, Herda TJ, Costa PB, Ryan ED, Stout JR, Cramer JT. Muscle performance, size, and safety responses after eight weeks of resistance training and protein supplementation: a randomized, double-blinded, placebo-controlled clinical trial. J Strength Cond Res. 2013;27:3091–3100. doi: 10.1519/JSC.0b013e31828c289f. https://www.ncbi.nlm.nih.gov/pubmed/23442287[↩]
- Willoughby DS, Stout JR, Wilborn CD. Effects of resistance training and protein plus amino acid supplementation on muscle anabolism, mass, and strength. Amino Acids. 2007;32:467–477. doi: 10.1007/s00726-006-0398-7. https://www.ncbi.nlm.nih.gov/pubmed/16988909[↩]
- Candow DG, Burke NC, Smith-Palmer T, Burke DG. Effect of whey and soy protein supplementation combined with resistance training in young adults. Int J Sport Nutr Exerc Metab. 2006;16:233–244. doi: 10.1123/ijsnem.16.3.233. https://www.ncbi.nlm.nih.gov/pubmed/16948480[↩]
- Hoffman JR, Ratamess NA, Kang J, Falvo MJ, Faigenbaum AD. Effects of protein supplementation on muscular performance and resting hormonal changes in college football players. J Sports Sci Med. 2007;6:85–92. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3778704/[↩][↩]
- Hoffman JR, Ratamess NA, Kang J, Falvo MJ, Faigenbaum AD. Effect of protein intake on strength, body composition and endocrine changes in strength/power athletes. J Int Soc Sports Nutr. 2006;3:12–18. doi: 10.1186/1550-2783-3-2-12. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2129168/[↩]
- Hida A, Hasegawa Y, Mekata Y, Usuda M, Masuda Y, Kawano H, et al. Effects of egg white protein supplementation on muscle strength and serum free amino acid concentrations. Nutrients. 2012;4:1504–17. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3497008/[↩]
- Moore DR, Robinson MJ, Fry JL, Tang JE, Glover EI, Wilkinson SB, et al. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. Am J Clin Nutr. 2009;89:161–8. https://www.ncbi.nlm.nih.gov/pubmed/19056590[↩][↩]
- Schoenfeld BJ, Aragon AA, Krieger JW. The effect of protein timing on muscle strength and hypertrophy: a meta-analysis. J Int Soc Sports Nutr. 2013;10:53. doi: 10.1186/1550-2783-10-53. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3879660/[↩]
- Josse AR, Tang JE, Tarnopolsky MA, Phillips SM. Body composition and strength changes in women with milk and resistance exercise. Med Sci Sports Exerc. 2010;42:1122–1130. https://www.ncbi.nlm.nih.gov/pubmed/19997019[↩]
- Taylor LW, Wilborn C, Roberts MD, White A, Dugan K. Eight weeks of pre- and postexercise whey protein supplementation increases lean body mass and improves performance in division III collegiate female basketball players. Appl Physiol Nutr Metab. 2016;41:249–254. doi: 10.1139/apnm-2015-0463. https://www.ncbi.nlm.nih.gov/pubmed/26842665[↩]
- Cermak NM, Res PT, De Groot LC, Saris WH, Van Loon LJ. Protein supplementation augments the adaptive response of skeletal muscle to resistance-type exercise training: a meta-analysis. Am J Clin Nutr. 2012;96:1454–1464. doi: 10.3945/ajcn.112.037556. https://www.ncbi.nlm.nih.gov/pubmed/23134885[↩]
- Pasiakos SM, Mclellan TM, Lieberman HR. The effects of protein supplements on muscle mass, strength, and aerobic and anaerobic power in healthy adults: a systematic review. Sports Med. 2015;45:111–131. doi: 10.1007/s40279-014-0242-2. https://www.ncbi.nlm.nih.gov/pubmed/25169440[↩]
- Phillips SM. A Brief Review of Critical Processes in Exercise-Induced Muscular Hypertrophy. Sports Medicine (Auckland, N.z). 2014;44(Suppl 1):71-77. doi:10.1007/s40279-014-0152-3. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4008813/[↩]
- Phillips SM, Tipton KD, Aarsland A, et al. Mixed muscle protein synthesis and breakdown after resistance exercise in humans. Am J Physiol. 1997;273:E99–E107. https://www.ncbi.nlm.nih.gov/pubmed/9252485[↩][↩]
- Miller BF, Olesen JL, Hansen M, et al. Coordinated collagen and muscle protein synthesis in human patella tendon and quadriceps muscle after exercise. J Physiol. 2005;567:1021–1033. doi: 10.1113/jphysiol.2005.093690. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1474228/[↩]
- Biolo G, Tipton KD, Klein S, et al. An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. Am J Physiol. 1997;273:E122–E129. https://www.ncbi.nlm.nih.gov/pubmed/9252488[↩][↩]
- Pennings B, Koopman R, Beelen M, et al. Exercising before protein intake allows for greater use of dietary protein-derived amino acids for de novo muscle protein synthesis in both young and elderly men. Am J Clin Nutr. 2011;93:322–331. doi: 10.3945/ajcn.2010.29649. https://www.ncbi.nlm.nih.gov/pubmed/21084649[↩]
- Biolo G, Maggi SP, Williams BD, et al. Increased rates of muscle protein turnover and amino acid transport after resistance exercise in humans. Am J Physiol. 1995;268:E514–E520. https://www.ncbi.nlm.nih.gov/pubmed/7900797[↩]
- Biolo G, Williams BD, Fleming RY, et al. Insulin action on muscle protein kinetics and amino acid transport during recovery after resistance exercise. Diabetes. 1999;48:949–957. doi: 10.2337/diabetes.48.5.949. https://www.ncbi.nlm.nih.gov/pubmed/10331397[↩][↩]
- Greenhaff PL, Karagounis L, Peirce N, et al. Disassociation between the effects of amino acids and insulin on signalling, ubiquitin-ligases and protein turnover in human muscle. Am J Physiol. 2008;295:E595–E604. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2536736/[↩]
- Staples AW, Burd NA, West DW, et al. Carbohydrate does not augment exercise-induced protein accretion versus protein alone. Med Sci Sports Exerc. 2011;43:1154–1161. doi: 10.1249/MSS.0b013e31820751cb. https://www.ncbi.nlm.nih.gov/pubmed/21131864[↩][↩]
- Moore DR, Robinson MJ, Fry JL, et al. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. Am J Clin Nutr. 2009;89:161–168. doi: 10.3945/ajcn.2008.26401. https://www.ncbi.nlm.nih.gov/pubmed/19056590[↩][↩][↩][↩][↩][↩]
- Volpi E, Kobayashi H, Sheffield-Moore M, et al. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am J Clin Nutr. 2003;78:250–258. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3192452/[↩]
- Crozier SJ, Kimball SR, Emmert SW, et al. Oral leucine administration stimulates protein synthesis in rat skeletal muscle. J Nutr. 2005;135:376–382. https://www.ncbi.nlm.nih.gov/pubmed/15735066[↩]
- Wilkinson DJ, Hossain T, Hill DS, et al. Effects of leucine and its metabolite beta-hydroxy-beta-methylbutyrate on human skeletal muscle protein metabolism. J Physiol. 2013;591:2911–2923. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3690694/[↩]
- Cuthbertson D, Smith K, Babraj J, et al. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J. 2005;19:422–424. https://www.ncbi.nlm.nih.gov/pubmed/15596483[↩]
- Yang Y, Breen L, Burd NA, et al. Resistance exercise enhances myofibrillar protein synthesis with graded intakes of whey protein in older men. Br J Nutr. 2012;108:1780–1788. doi: 10.1017/S0007114511007422. https://www.ncbi.nlm.nih.gov/pubmed/22313809[↩][↩][↩][↩]
- Devkota S, Layman DK. Increased ratio of dietary carbohydrate to protein shifts the focus of metabolic signaling from skeletal muscle to adipose. Nutr Metab. 2011;8:13. doi: 10.1186/1743-7075-8-13. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3059285/[↩]
- Lemon PW. Beyond the zone: protein needs of active individuals. J Am Coll Nutr. 2000;19(5 Suppl):513S–521S. doi: 10.1080/07315724.2000.10718974. https://www.ncbi.nlm.nih.gov/pubmed/11023001[↩]
- Campbell WW, Barton ML Jr, Cyr-Campbell D, Davey SL, Beard JL, Parise G, et al. Effects of an omnivorous diet compared with a lactoovovegetarian diet on resistance-training-induced changes in body composition and skeletal muscle in older men. Am J Clin Nutr. 1999;70:1032–9. https://www.ncbi.nlm.nih.gov/pubmed/10584048[↩][↩][↩][↩]
- Katsanos CS, Chinkes DL, Paddon-Jones D, Zhang XJ, Aarsland A, Wolfe RR. Whey protein ingestion in elderly persons results in greater muscle protein accrual than ingestion of its constituent essential amino acid content. Nutr Res. 2008;28:651–658. doi: 10.1016/j.nutres.2008.06.007. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2612691/[↩][↩]
- Phillips SM, Tipton KD, Aarsland A, Wolf SE, Wolfe RR. Mixed muscle protein synthesis and breakdown after resistance exercise in humans. Am J Phys. 1997;273(1 Pt 1):E99–107. https://www.ncbi.nlm.nih.gov/pubmed/9252485[↩]
- Borsheim E, Tipton KD, Wolf SE, Wolfe RR. Essential amino acids and muscle protein recovery from resistance exercise. Am J Physiol Endocrinol Metab. 2002;283:E648–E657. doi: 10.1152/ajpendo.00466.2001. https://www.ncbi.nlm.nih.gov/pubmed/12217881[↩][↩]
- Paddon-Jones D, Sheffield-Moore M, Aarsland A, Wolfe RR, Ferrando AA. Exogenous amino acids stimulate human muscle anabolism without interfering with the response to mixed meal ingestion. Am J Physiol Endocrinol Metab. 2005;288:E761–E767. doi: 10.1152/ajpendo.00291.2004. https://www.ncbi.nlm.nih.gov/pubmed/15572657[↩]
- Cuthbertson D, Smith K, Babraj J, Leese G, Waddell T, Atherton P, et al. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J. 2005;19:422–4. https://www.ncbi.nlm.nih.gov/pubmed/15596483[↩]
- Norton LE, Layman DK. Leucine regulates translation initiation of protein synthesis in skeletal muscle after exercise. J Nutr. 2006;136:533S–537S. https://www.ncbi.nlm.nih.gov/pubmed/16424142[↩][↩][↩]
- Blomstrand E. A role for branched-chain amino acids in reducing central fatigue. J Nutr. 2006;136:544S–547S. https://www.ncbi.nlm.nih.gov/pubmed/16424144[↩]
- Brosnan JT, Brosnan ME. Branched-chain amino acids: enzyme and substrate regulation. J Nutr. 2006;136(1 Suppl):207S–211S. https://www.ncbi.nlm.nih.gov/pubmed/16365084[↩]
- Norton L, Wilson GJ. Optimal protein intake to maximize muscle protein synthesis. AgroFood Industry Hi-Tech. 2009;20:54–57.[↩][↩]
- Glynn EL, Fry CS, Drummond MJ, Timmerman KL, Dhanani S, Volpi E, et al. Excess leucine intake enhances muscle anabolic signaling but not net protein anabolism in young men and women. J Nutr. 2010;140:1970–6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2955876/[↩][↩]
- Pasiakos SM, Mcclung HL, Mcclung JP, Margolis LM, Andersen NE, Cloutier GJ, et al. Leucine-enriched essential amino acid supplementation during moderate steady state exercise enhances postexercise muscle protein synthesis. Am J Clin Nutr. 2011;94:809–18. https://www.ncbi.nlm.nih.gov/pubmed/21775557[↩]
- Churchward-Venne TA, Burd NA, Mitchell CJ, West DW, Philp A, Marcotte GR, et al. Supplementation of a suboptimal protein dose with leucine or essential amino acids: effects on myofibrillar protein synthesis at rest and following resistance exercise in men. J Physiol. 2012;590:2751–65. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3424729/[↩][↩]
- Sarwar G. (1997) The protein digestibility-corrected amino acid score method overestimates quality of proteins containing antinutritional factors and of poorly digestible proteins supplemented with limiting amino acids in rats. Journal of Nutrition 127, 758-764. https://www.ncbi.nlm.nih.gov/pubmed/9164998[↩]
- J Sports Sci Med. 2004 Sep; 3(3): 118–130. International Society of Sports Nutrition Symposium, June 18-19, 2005, Las Vegas NV, USA – Symposium – Macronutrient Utilization During Exercise: Implications For Performance And Supplementation. Protein – Which is Best ? https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3905294/[↩][↩]
- Layman DK. Role of leucine in protein metabolism during exercise and recovery. Can J Appl Physiol. 2002;27:646–663. doi: 10.1139/h02-038. https://www.ncbi.nlm.nih.gov/pubmed/12501002[↩]
- Wilson GJ, Layman DK, Moulton CJ, Norton LE, Anthony TG, Proud CG, et al. Leucine or carbohydrate supplementation reduces AMPK and eef2 phosphorylation and extends postprandial muscle protein synthesis in rats. Am J Physiol Endocrinol Metab. 2011;301:E1236–42. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4395871/[↩][↩][↩]
- Boirie Y, Dangin M, Gachon P, Vasson MP, Maubois JL, Beaufrere B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc Natl Acad Sci. 1997;94:14930–14935. doi: 10.1073/pnas.94.26.14930. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC25140/[↩]
- Norton LE, Rupassara I, Garlick PJ, Layman DK. Leucine contents of isonitrogenout protein sources predict post prandial muscle protein synthesis in rats fed a complete meal. FASEB J. 2009;23(Supplement 1):227–224[↩]
- Tang JE, Moore DR, Kujbida GW, Tarnopolsky MA, Phillips SM. Ingestion of whey hydrolysate, casein, or soy protein isolate: effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J App Physiol (Bethesda, Md: 1985) 2009;107:987–992. https://www.ncbi.nlm.nih.gov/pubmed/19589961[↩][↩][↩][↩]
- Renan M, Mekmene O, Famelart MH, Guyomarc’h F, Arnoult-Delest V, Paquet D, et al. Ph-dependent behaviour of soluble protein aggregates formed during heat-treatment of milk at ph 6.5 or 7.2. J Dairy Res. 2006;73:79–86. https://www.ncbi.nlm.nih.gov/pubmed/16433965[↩][↩][↩][↩][↩][↩]
- Silk DB, Grimble GK, Rees RG. Protein digestion and amino acid and peptide absorption. Proc Nutr Soc. 1985;44:63–72. doi: 10.1079/PNS19850011. https://www.ncbi.nlm.nih.gov/pubmed/3885229[↩][↩][↩]
- Buckley JD, Thomson RL, Coates AM, Howe PR, Denichilo MO, Rowney MK. Supplementation with a whey protein hydrolysate enhances recovery of muscle force-generating capacity following eccentric exercise. J Sci Med Sport. 2010;13:178–181. doi: 10.1016/j.jsams.2008.06.007. https://www.ncbi.nlm.nih.gov/pubmed/18768358[↩][↩]
- Morifuji M, Ishizaka M, Baba S, Fukuda K, Matsumoto H, Koga J, et al. Comparison of different sources and degrees of hydrolysis of dietary protein: effect on plasma amino acids, dipeptides, and insulin responses in human subjects. J Agric Food Chem. 2010;58:8788–97. https://www.ncbi.nlm.nih.gov/pubmed/20614926[↩][↩]
- Calbet JA, Maclean DA. Plasma glucagon and insulin responses depend on the rate of appearance of amino acids after ingestion of different protein solutions in humans. J Nutr. 2002;132:2174–2182. https://www.ncbi.nlm.nih.gov/pubmed/12163658[↩]
- Calbet JA, Holst JJ. Gastric emptying, gastric secretion and enterogastrone response after administration of milk proteins or their peptide hydrolysates in humans. Eur J Nutr. 2004;43:127–139. doi: 10.1007/s00394-004-0448-4. https://www.ncbi.nlm.nih.gov/pubmed/15168035[↩]
- Boyer N, Chuang JL, Gipner D. An acute care geriatric unit. Nurs Manag. 1986;17:22–25. https://www.ncbi.nlm.nih.gov/pubmed/3634968[↩]
- Cooke MB, Rybalka E, Stathis CG, Cribb PJ, Hayes A. Whey protein isolate attenuates strength decline after eccentrically-induced muscle damage in healthy individuals. J Int Soc Sports Nutr. 2010;7:30. doi: 10.1186/1550-2783-7-30. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2955583/[↩]
- Morifuji M, Kanda A, Koga J, Kawanaka K, Higuchi M. Post-exercise carbohydrate plus whey protein hydrolysates supplementation increases skeletal muscle glycogen level in rats. Amino Acids. 2010;38:1109–1115. doi: 10.1007/s00726-009-0321-0. https://www.ncbi.nlm.nih.gov/pubmed/19593593[↩]
- Lockwood CM, Roberts MD, Dalbo VJ, Smith-Ryan AE, Kendall KL, Moon JR, et al. Effects of hydrolyzed whey versus other whey protein supplements on the physiological response to 8 weeks of resistance exercise in college-aged males. J Am Coll Nutr. 2017;36:16–27. https://www.ncbi.nlm.nih.gov/pubmed/27710436[↩]
- Saunders MJ, Moore RW, Kies AK, Luden ND, Pratt CA. Carbohydrate and protein hydrolysate coingestions improvement of late-exercise time-trial performance. Int J Sport Nutr Exerc Metab. 2009;19:136–149. doi: 10.1123/ijsnem.19.2.136. https://www.ncbi.nlm.nih.gov/pubmed/19478339[↩]
- Van Loon LJ, Kies AK, Saris WH. Protein and protein hydrolysates in sports nutrition. Int J Sport Nutr Exerc Metab. 2007;17:S1–S4. doi: 10.1123/ijsnem.17.s1.s1. https://www.ncbi.nlm.nih.gov/pubmed/18577770[↩]
- Saunders MJ. Coingestion of carbohydrate-protein during endurance exercise: influence on performance and recovery. Int J Sport Nutr Exerc Metab. 2007;17:S87–S103. doi: 10.1123/ijsnem.17.s1.s87. https://www.ncbi.nlm.nih.gov/pubmed/18577778[↩]
- Borgstrom B, Dahlqvist A, Lundh G, Sjovall J. Studies of intestinal digestion and absorption in the human. J Clin Invest. 1957;36:1521–1536. doi: 10.1172/JCI103549. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1072752/[↩]
- Minevich J, Olson MA, Mannion JP, Boublik JH, Mcpherson JO, Lowery RP, et al. Digestive enzymes reduce quality differences between plant and animal proteins: an double-blind crossover study. J Int Soc Sports Nutr. 2015;12(Suppl 1):P26.[↩]
- Effects of an omnivorous diet compared with a lactoovovegetarian diet on resistance-training-induced changes in body composition and skeletal muscle in older men. Campbell WW, Barton ML Jr, Cyr-Campbell D, Davey SL, Beard JL, Parise G, Evans WJ. Am J Clin Nutr. 1999 Dec; 70(6):1032-9. https://www.ncbi.nlm.nih.gov/pubmed/10584048/[↩][↩]
- Maternal nutrition in early and late pregnancy in relation to placental and fetal growth. Godfrey K, Robinson S, Barker DJ, Osmond C, Cox V. BMJ. 1996 Feb 17; 312(7028):410-4. https://www.ncbi.nlm.nih.gov/pubmed/8601112/[↩]
- Effect of protein source and quantity on protein metabolism in elderly women. Pannemans DL, Wagenmakers AJ, Westerterp KR, Schaafsma G, Halliday D. Am J Clin Nutr. 1998 Dec; 68(6):1228-35. https://www.ncbi.nlm.nih.gov/pubmed/9846851/[↩]
- Hartman JW, Tang JE, Wilkinson SB, Tarnopolsky MA, Lawrence RL, Fullerton AV, et al. Consumption of fat-free fluid milk after resistance exercise promotes greater lean mass accretion than does consumption of soy or carbohydrate in young, novice, male weightlifters. Am J Clin Nutr. 2007;86(2):373–81. https://www.ncbi.nlm.nih.gov/pubmed/17684208[↩][↩][↩][↩][↩]
- Cockburn E, Stevenson E, Hayes PR, Robson-Ansley P, Howatson G. Effect of milk-based carbohydrate-protein supplement timing on the attenuation of exercise-induced muscle damage. Appl Physiol Nutr Metab. 2010;35:270–277. doi: 10.1139/H10-017. https://www.ncbi.nlm.nih.gov/pubmed/20555370[↩]
- Wojcik JR, Walber-Rankin J, Smith LL, Gwazdauskas FC. Comparison of carbohydrate and milk-based beverages on muscle damage and glycogen following exercise. Int J Sport Nutr Exerc Metab. 2001;11:406–419. doi: 10.1123/ijsnem.11.4.406. https://www.ncbi.nlm.nih.gov/pubmed/11915776[↩]
- Watson P, Love TD, Maughan RJ, Shirreffs SM. A comparison of the effects of milk and a carbohydrate-electrolyte drink on the restoration of fluid balance and exercise capacity in a hot, humid environment. Eur J Appl Physiol. 2008;104:633–642. doi: 10.1007/s00421-008-0809-4. https://www.ncbi.nlm.nih.gov/pubmed/18618137[↩]
- Dangin M, Guillet C, Garcia-Rodenas C, Gachon P, Bouteloup-Demange C, Reiffers-Magnani K, et al. The rate of protein digestion affects protein gain differently during aging in humans. J Physiol. 2003;549(Pt 2):635–44. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2342962/[↩]
- Wilson J, Wilson GJ. Contemporary issues in protein requirements and consumption for resistance trained athletes. J Int Soc Sports Nutr. 2006;3:7–27. doi: 10.1186/1550-2783-3-1-7. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2129150/[↩][↩][↩][↩]
- Dangin M, Boirie Y, Guillet C, Beaufrere B. Influence of the protein digestion rate on protein turnover in young and elderly subjects. J Nutr. 2002;132:3228S–3233S. https://www.ncbi.nlm.nih.gov/pubmed/12368423[↩]
- Nair KS. Muscle protein turnover: methodological issues and the effect of aging. J Gerontol A Biol Sci Med Sci. 1995;50(Spec Issue):107–112. https://www.ncbi.nlm.nih.gov/pubmed/7493201[↩]
- Trommelen J, Holwerda AM, Kouw IW, Langer H, Halson SL, Rollo I, et al. Resistance exercise augments postprandial overnight muscle protein synthesis rates. Med Sci Sports Exerc. 2016;48:2517–25. https://www.ncbi.nlm.nih.gov/pubmed/27643743[↩]
- Tipton KD, Elliott TA, Cree MG, Wolf SE, Sanford AP, Wolfe RR. Ingestion of casein and whey proteins result in muscle anabolism after resistance exercise. Med Sci Sports Exerc. 2004;36:2073–2081. doi: 10.1249/01.MSS.0000147582.99810.C5. https://www.ncbi.nlm.nih.gov/pubmed/15570142[↩][↩]
- Cribb PJ, Williams AD, Carey MF, Hayes A. The effect of whey isolate and resistance training on strength, body composition, and plasma glutamine. Int J Sport Nutr Exerc Metab. 2006;16:494–509. doi: 10.1123/ijsnem.16.5.494. https://www.ncbi.nlm.nih.gov/pubmed/17240782[↩]
- Zawadzki KM, Yaspelkis BB, 3rd, Ivy JL. Carbohydrate-protein complex increases the rate of muscle glycogen storage after exercise. J Appl Physiol. (Bethesda, Md : 1985) 1992;72:1854–1859. https://www.ncbi.nlm.nih.gov/pubmed/1601794[↩]
- Saunders MJ, Luden ND, Herrick JE. Consumption of an oral carbohydrate-protein gel improves cycling endurance and prevents postexercise muscle damage. J Strength Cond Res. 2007;21:678–684. https://www.ncbi.nlm.nih.gov/pubmed/17685703[↩]
- Morifuji M, Sakai K, Sanbongi C, Sugiura K. Dietary whey protein increases liver and skeletal muscle glycogen levels in exercise-trained rats. Br J Nutr. 2005;93:439–445. doi: 10.1079/BJN20051373. https://www.ncbi.nlm.nih.gov/pubmed/15946405[↩]
- Staples AW, Burd NA, West DW, Currie KD, Atherton PJ, Moore DR, et al. Carbohydrate does not augment exercise-induced protein accretion versus protein alone. Med Sci Sports Exerc. 2011;43:1154–61. https://www.ncbi.nlm.nih.gov/pubmed/21131864[↩]
- Markus CR, Olivier B, De Haan EH. Whey protein rich in alpha-lactalbumin increases the ratio of plasma tryptophan to the sum of the other large neutral amino acids and improves cognitive performance in stress-vulnerable subjects. Am J Clin Nutr. 2002;75:1051–1056. https://www.ncbi.nlm.nih.gov/pubmed/12036812[↩][↩][↩]
- Law BA, Reiter B. The isolation and bacteriostatic properties of lactoferrin from bovine milk whey. J Dairy Res. 1977;44:595–599. doi: 10.1017/S0022029900020550. https://www.ncbi.nlm.nih.gov/pubmed/338651[↩]
- Wang X, Ai T, Meng XL, Zhou J, Mao XY. In vitro iron absorption of alpha-lactalbumin hydrolysate-iron and beta-lactoglobulin hydrolysate-iron complexes. J Dairy Sci. 2014;97:2559–2566. doi: 10.3168/jds.2013-7461. https://www.ncbi.nlm.nih.gov/pubmed/24612808[↩]
- Purification and characterization of cell growth factor in bovine colostrum. Kishikawa Y, Watanabe T, Watanabe T, Kubo S. J Vet Med Sci. 1996 Jan; 58(1):47-53. https://www.ncbi.nlm.nih.gov/pubmed/8645756/[↩]
- Bovine colostrum supplementation does not affect plasma buffer capacity or haemoglobin content in elite female rowers. Brinkworth GD, Buckley JD. Eur J Appl Physiol. 2004 Mar; 91(2-3):353-6. https://www.ncbi.nlm.nih.gov/pubmed/14685867/[↩]
- The effect of bovine colostrum supplementation on exercise performance in elite field hockey players. Hofman Z, Smeets R, Verlaan G, Lugt Rv, Verstappen PA. Int J Sport Nutr Exerc Metab. 2002 Dec; 12(4):461-9. https://www.ncbi.nlm.nih.gov/pubmed/12500989/[↩]
- Geiser M. (2003) The wonders of whey protein. NSCA’s Performance Training Journal 2, 13-15[↩][↩]
- Whey protein concentrate (WPC) and glutathione modulation in cancer treatment. Bounous G. Anticancer Res. 2000 Nov-Dec; 20(6C):4785-92. https://www.ncbi.nlm.nih.gov/pubmed/11205219/[↩]
- Functional properties of whey, whey components, and essential amino acids: mechanisms underlying health benefits for active people (review). Ha E, Zemel MB. J Nutr Biochem. 2003 May; 14(5):251-8. https://www.ncbi.nlm.nih.gov/pubmed/12832028/[↩]
- Branched-chain amino acids augment ammonia metabolism while attenuating protein breakdown during exercise. MacLean DA, Graham TE, Saltin B. Am J Physiol. 1994 Dec; 267(6 Pt 1):E1010-22. https://www.ncbi.nlm.nih.gov/pubmed/7810616/[↩]
- Infusion of soy and casein protein meals affects interorgan amino acid metabolism and urea kinetics differently in pigs. Deutz NE, Bruins MJ, Soeters PB. J Nutr. 1998 Dec; 128(12):2435-45. https://www.ncbi.nlm.nih.gov/pubmed/9868192/[↩]
- Slow and fast dietary proteins differently modulate postprandial protein accretion. Boirie Y, Dangin M, Gachon P, Vasson MP, Maubois JL, Beaufrère B. Proc Natl Acad Sci U S A. 1997 Dec 23; 94(26):14930-5. https://www.ncbi.nlm.nih.gov/pubmed/9405716/[↩]
- Hu FB, Stampfer MJ, Rimm EB, Manson JE, Ascherio A, Colditz GA, et al. A prospective study of egg consumption and risk of cardiovascular disease in men and women. JAMA. 1999;281:1387–94. https://www.ncbi.nlm.nih.gov/pubmed/10217054[↩]
- Hasler CM. The changing face of functional foods. J Am Coll Nutr. 2000;19(Suppl 5):499S–506S. doi: 10.1080/07315724.2000.10718972. https://www.ncbi.nlm.nih.gov/pubmed/11022999[↩][↩]
- Safety assessment and potential health benefits of food components based on selected scientific criteria. ILSI North America technical committee on food components for health promotion. Crit Rev Food Sci Nutr. 1999;39:203–316. https://www.ncbi.nlm.nih.gov/pubmed/10367185[↩]
- Blusztajn JK. Choline, a vital amine. Science. 1998;281(5378):794–795. doi: 10.1126/science.281.5378.794. https://www.ncbi.nlm.nih.gov/pubmed/9714685[↩]
- Handelman GJ, Nightingale ZD, Lichtenstein AH, Schaefer EJ, Blumberg JB. Lutein and zeaxanthin concentrations in plasma after dietary supplementation with egg yolk. Am J Clin Nutr. 1999;70:247–251. https://www.ncbi.nlm.nih.gov/pubmed/10426702[↩]
- Symons TB, Schutzler SE, Cocke TL, Chinkes DL, Wolfe RR, Paddon-Jones D. Aging does not impair the anabolic response to a protein-rich meal. Am J Clin Nutr. 2007;86:451–456. https://www.ncbi.nlm.nih.gov/pubmed/17684218[↩][↩][↩]
- Fogelholm M. Dairy products, meat and sports performance. Sports Med. 2003;33:615–631. doi: 10.2165/00007256-200333080-00006. https://www.ncbi.nlm.nih.gov/pubmed/12797842[↩]
- Kreider RB, Wilborn CD, Taylor L, Campbell B, Almada AL, Collins R, et al. Issn exercise & sports nutrition review: research & recommendations. J Int Soc Sports Nutr. 2010;7:7.[↩]
- Harris RC, Lowe JA, Warnes K, Orme CE. The concentration of creatine in meat, offal and commercial dog food. Res Vet Sci. 1997;62:58–62. doi: 10.1016/S0034-5288(97)90181-8. https://www.ncbi.nlm.nih.gov/pubmed/9160426[↩]
- Shomrat A, Weinstein Y, Katz A. Effect of creatine feeding on maximal exercise performance in vegetarians. Eur J Appl Physiol. 2000;82:321–325. doi: 10.1007/s004210000222. https://www.ncbi.nlm.nih.gov/pubmed/10958375[↩]
- Sharp MH, Lowery RP, Shields KA, Lane JR, Gray JL, Partl JM, Hayes DW, Wilson GJ, Hollmer CA, Minivich JR, and Wilson JM. The effects of beef, chicken, or whey protein post-workout on body composition and muscle performance. J Strength Cond Res. 2017. doi: 10.1519/JSC.0000000000001936. https://www.ncbi.nlm.nih.gov/pubmed/28399016[↩]
- Larsson SC, Wolk K, Brismar K, Wolk A. Association of diet with serum insulin-like growth factor i in middle-aged and elderly men. Am J Clin Nutr. 2005;81:1163–1167. https://www.ncbi.nlm.nih.gov/pubmed/15883443[↩]
- Juul A, Scheike T, Davidsen M, Gyllenborg J, Jorgensen T. Low serum insulin-like growth factor i is associated with increased risk of ischemic heart disease: a population-based case-control study. Circulation. 2002;106:939–944. doi: 10.1161/01.CIR.0000027563.44593.CC. https://www.ncbi.nlm.nih.gov/pubmed/12186797[↩]
- Cederroth CR, Vinciguerra M, Gjinovci A, Kuhne F, Klein M, Cederroth M, et al. Dietary phytoestrogens activate amp-activated protein kinase with improvement in lipid and glucose metabolism. Diabetes. 2008;57:1176–85. https://www.ncbi.nlm.nih.gov/pubmed/18420492[↩]
- Gorissen SH, Horstman AM, Franssen R, Crombag JJ, Langer H, Bierau J, et al. Ingestion of wheat protein increases in vivo muscle protein synthesis rates in healthy older men in a randomized trial. J Nutr. 2016;146:1651–9. https://www.ncbi.nlm.nih.gov/pubmed/27440260[↩]
- Purpura M, Lowery RP, Joy JM, De Souza EO, Kalman D. A comparison of blood amino acid concentrations following ingestion of rice and whey protein isolate: a double-blind, crossover study. J Nutr Health Sci. 2014;1:306.[↩]
- Joy JM, Lowery RP, Wilson JM, Purpura M, De Souza EO, Wilson SM, et al. The effects of 8 weeks of whey or rice protein supplementation on body composition and exercise performance. Nutr J. 2013;12:86. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3698202/[↩][↩]
- Paul GL. The rationale for consuming protein blends in sports nutrition. J Am Coll Nutr. 2009;28(sup4):464S–472S. doi: 10.1080/07315724.2009.10718113. https://www.ncbi.nlm.nih.gov/pubmed/20234034[↩]
- Kerksick CM, Rasmussen CJ, Lancaster SL, Magu B, Smith P, Melton C, et al. The effects of protein and amino acid supplementation on performance and training adaptations during ten weeks of resistance training. J Strength Cond Res. 2006;20:643–53. https://www.ncbi.nlm.nih.gov/pubmed/16937979[↩]
- Kerksick CM, Rasmussen C, Lancaster S, Starks M, Smith P, Melton C, et al. Impact of differing protein sources and a creatine containing nutritional formula after 12 weeks of resistance training. Nutrition. 2007;23:647–56. https://www.ncbi.nlm.nih.gov/pubmed/17679046[↩]
- Wilkinson SB, Tarnopolsky MA, Macdonald MJ, Macdonald JR, Armstrong D, Phillips SM. Consumption of fluid skim milk promotes greater muscle protein accretion after resistance exercise than does consumption of an isonitrogenous and isoenergetic soy-protein beverage. Am J Clin Nutr. 2007;85:1031–1040. https://www.ncbi.nlm.nih.gov/pubmed/17413102[↩]
- Reidy PT, Walker DK, Dickinson JM, Gundermann DM, Drummond MJ, Timmerman KL, et al. Protein blend ingestion following resistance exercise promotes human muscle protein synthesis. J Nutr. 2013;143:410–6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3738242/[↩]
- Reidy PT, Walker DK, Dickinson JM, Gundermann DM, Drummond MJ, Timmerman KL, et al. Soy-dairy protein blend and whey protein ingestion after resistance exercise increases amino acid transport and transporter expression in human skeletal muscle. J Appl Physiol (Bethesda, Md: 1985). 2014;116:1353–64. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4044402/[↩]
- Reidy PT, Borack MS, Markofski MM, Dickinson JM, Deer RR, Husaini SH, et al. Protein supplementation has minimal effects on muscle adaptations during resistance exercise training in young men: a double-blind randomized clinical trial. J Nutr. 2016;146:1660–9. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4997282/[↩]
- Fouque D, Laville M. Low protein diets for chronic kidney disease in non diabetic adults. Cochrane Database Syst Rev. 2009 Jul 8;(3):CD001892. doi: 10.1002/14651858.CD001892.pub3. https://www.ncbi.nlm.nih.gov/pubmed/19588328[↩]
- Poortmans JR, Dellalieux O. Do regular high protein diets have potential health risks on kidney function in athletes? Int J Sport Nutr Exerc Metab. 2000;10:28–38. doi: 10.1123/ijsnem.10.1.28. https://www.ncbi.nlm.nih.gov/pubmed/10722779[↩]
- World Health Organization . Technical report series 935. Protein and amino acid requirements in human nutrition: report of a joint fao/who/uni expert consultation. 2011. https://www.ncbi.nlm.nih.gov/pubmed/18330140[↩]
- Brandle E, Sieberth HG, Hautmann RE. Effect of chronic dietary protein intake on the renal function in healthy subjects. Eur J Clin Nutr. 1996;50:734–740. https://www.ncbi.nlm.nih.gov/pubmed/8933120[↩]
- Antonio J, Peacock CA, Ellerbroek A, Fromhoff B, Silver T. The effects of consuming a high protein diet (4.4 g/kg/d) on body composition in resistance-trained individuals. J Int Soc Sports Nutr. 2014;11:19. doi: 10.1186/1550-2783-11-19. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4022420/[↩]
- Antonio J, Ellerbroek A, Silver T, Orris S, Scheiner M, Gonzalez A, et al. A high protein diet (3.4 g/kg/d) combined with a heavy resistance training program improves body composition in healthy trained men and women–a follow-up investigation. J Int Soc Sports Nutr. 2015;12:39. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4617900/[↩]
- Antonio J, Ellerbroek A, Silver T, Vargas L, Peacock C. The effects of a high protein diet on indices of health and body composition–a crossover trial in resistance-trained men. J Int Soc Sports Nutr. 2016;13:3. doi: 10.1186/s12970-016-0114-2. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4715299/[↩]
- Antonio J, Ellerbroek A, Silver T, Vargas L, Tamayo A, Buehn R, et al. A high protein diet has no harmful effects: a one-year crossover study in resistance-trained males. J Nutr Metab. 2016;2016:9104792. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5078648/[↩]
- Thomson DM, Fick CA, Gordon SE. AMPK activation attenuates S6K1, 4E-BP1, and eEF2 signaling responses to high-frequency electrically stimulated skeletal muscle contractions. J Appl Physiol. 2008;104:625–632. doi: 10.1152/japplphysiol.00915.2007. https://www.ncbi.nlm.nih.gov/pubmed/18187610[↩]
- Rose AJ, Alsted TJ, Jensen TE, et al. A Ca(2+)–calmodulin–eEF2K–eEF2 signalling cascade, but not AMPK, contributes to the suppression of skeletal muscle protein synthesis during contractions. J Physiol. 2009;587:1547–1563. doi: 10.1113/jphysiol.2008.167528. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2678225/[↩]
- Phillips SM. Insulin and muscle protein turnover in humans: stimulatory, permissive, inhibitory, or all of the above? Am J Physiol. 2008;295:E731. https://www.ncbi.nlm.nih.gov/pubmed/18628353[↩]
- Koopman R, Beelen M, Stellingwerff T, et al. Coingestion of carbohydrate with protein does not further augment postexercise muscle protein synthesis. Am J Physiol. 2007;293:E833–E842. doi: 10.1152/ajpcell.00194.2006. https://www.ncbi.nlm.nih.gov/pubmed/17609259[↩]
- Glynn EL, Fry CS, Drummond MJ, et al. Muscle protein breakdown has a minor role in the protein anabolic response to essential amino acid and carbohydrate intake following resistance exercise. Am J Physiol. 2010;299:R533–R540. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2928613/[↩]
- Wilkinson SB, Kim PL, Armstrong D, et al. Addition of glutamine to essential amino acids and carbohydrate does not enhance anabolism in young human males following exercise. Appl Physiol Nutr Metab. 2006;31:518–529. doi: 10.1139/h06-028. https://www.ncbi.nlm.nih.gov/pubmed/17111006[↩]
- Wilkinson SB, Phillips SM, Atherton PJ, et al. Differential effects of resistance and endurance exercise in the fed state on signalling molecule phosphorylation and protein synthesis in human muscle. J Physiol. 2008;586:3701–3717. doi: 10.1113/jphysiol.2008.153916. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2538832/[↩]
- Candow DG, Chilibeck PD, Burke DG, et al. Effect of glutamine supplementation combined with resistance training in young adults. Eur J Appl Physiol. 2001;86:142–149. doi: 10.1007/s00421-001-0523-y. https://www.ncbi.nlm.nih.gov/pubmed/11822473[↩][↩]
- Soeters PB, Grecu I. Have we enough glutamine and how does it work? A clinician’s view. Ann Nutr Metab. 2012;60:17–26. doi: 10.1159/000334880. https://www.ncbi.nlm.nih.gov/pubmed/22212454[↩]
- Gleeson M. Dosing and efficacy of glutamine supplementation in human exercise and sport training. J Nutr. 2008;138:2045S–2049S. https://www.ncbi.nlm.nih.gov/pubmed/18806122[↩][↩][↩]
- Willoughby DS, Boucher T, Reid J, et al. Effects of 7 days of arginine-alpha-ketoglutarate supplementation on blood flow, plasma l-arginine, nitric oxide metabolites, and asymmetric dimethyl arginine after resistance exercise. Int J Sport Nutr Exerc Metab. 2011;21:291–299. https://www.ncbi.nlm.nih.gov/pubmed/21813912[↩]
- Tang JE, Lysecki PJ, Manolakos JJ, et al. Bolus arginine supplementation affects neither muscle blood flow nor muscle protein synthesis in young men at rest or after resistance exercise. J Nutr. 2011;141:195–200. doi: 10.3945/jn.110.130138. https://www.ncbi.nlm.nih.gov/pubmed/21191143[↩][↩]
- West DW, Burd NA, Tang JE, et al. Elevations in ostensibly anabolic hormones with resistance exercise enhance neither training-induced muscle hypertrophy nor strength of the elbow flexors. J Appl Physiol. 2010;108:60–67. doi: 10.1152/japplphysiol.01147.2009. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2885075/[↩]
- Alvares TS, Conte CA, Paschoalin VM, et al. Acute l-arginine supplementation increases muscle blood volume but not strength performance. Appl Physiol Nutr Metab. 2012;37:115–126. doi: 10.1139/h11-144. https://www.ncbi.nlm.nih.gov/pubmed/22251130[↩]
- Tarnopolsky MA, Macdougall JD, Atkinson SA. Influence of protein intake and training status on nitrogen balance and lean body mass. J App Physiol (Bethesda, Md: 1985) 1988;64:187–193. https://www.ncbi.nlm.nih.gov/pubmed/3356636[↩]
- Phillips SM, Atkinson SA, Tarnopolsky MA, Macdougall JD. Gender differences in leucine kinetics and nitrogen balance in endurance athletes. J App Physiol (Bethesda, Md: 1985) 1993;75:2134–2141. https://www.ncbi.nlm.nih.gov/pubmed/8307870[↩]
- Lemon PW. Effect of exercise on protein requirements. J Sports Sci. 1991;9(S1):53–70. doi: 10.1080/02640419108729866. https://www.ncbi.nlm.nih.gov/pubmed/1895363[↩]
- Dietary protein for athletes: from requirements to optimum adaptation. Phillips SM, Van Loon LJ. J Sports Sci. 2011; 29 Suppl 1():S29-38. https://www.ncbi.nlm.nih.gov/pubmed/22150425/[↩][↩]
- A systematic review of dietary protein during caloric restriction in resistance trained lean athletes: a case for higher intakes. Helms ER, Zinn C, Rowlands DS, Brown SR. Int J Sport Nutr Exerc Metab. 2014 Apr; 24(2):127-38. https://www.ncbi.nlm.nih.gov/pubmed/24092765/[↩][↩]
- What is the required energy deficit per unit weight loss ? Hall KD. Int J Obes (Lond). 2008 Mar; 32(3):573-6. https://www.ncbi.nlm.nih.gov/pubmed/17848938/[↩]
- Differences in fat, carbohydrate, and protein metabolism between lean and obese subjects undergoing total starvation. Elia M, Stubbs RJ, Henry CJ. Obes Res. 1999 Nov; 7(6):597-604. https://www.ncbi.nlm.nih.gov/pubmed/10574520/[↩]
- Butterfield GE. Whole-body protein utilization in humans. Med Sci Sports Exerc. 1987;19:S157–S165. https://www.ncbi.nlm.nih.gov/pubmed/3316915[↩]
- Celejowa I, Homa M. Food intake, nitrogen and energy balance in Polish weight lifters, during a training camp. Nutr Metab. 1970;12:259–274. doi: 10.1159/000175300. https://www.ncbi.nlm.nih.gov/pubmed/5510411[↩]
- Walberg JL, Leidy MK, Sturgill DJ, Hinkle DE, Ritchey SJ, Sebolt DR. Macronutrient content of a hypoenergy diet affects nitrogen retention and muscle function in weight lifters. Int J Sports Med. 1988;9:261–266. doi: 10.1055/s-2007-1025018. https://www.ncbi.nlm.nih.gov/pubmed/3182156[↩][↩][↩][↩][↩]
- Mettler S, Mitchell N, Tipton KD. Increased protein intake reduces lean body mass loss during weight loss in athletes. Med Sci Sports Exerc. 2010;42:326–337. https://www.ncbi.nlm.nih.gov/pubmed/19927027[↩][↩][↩][↩]
- Pasiakos SM, Cao JJ, Margolis LM, Sauter ER, Whigham LD, McClung JP, Rood JC, Carbone JW, Combs GF Jr, Young AJ. Effects of high-protein diets on fat-free mass and muscle protein synthesis following weight loss: a randomized controlled trial. FASEB J. 2013;27:3837–3847. doi: 10.1096/fj.13-230227. https://www.ncbi.nlm.nih.gov/pubmed/23739654[↩][↩]
- Maestu J, Eliakim A, Jurimae J, Valter I, Jurimae T. Anabolic and catabolic hormones and energy balance of the male bodybuilders during the preparation for the competition. J Strength Cond Res. 2010;24:1074–1081. doi: 10.1519/JSC.0b013e3181cb6fd3. https://www.ncbi.nlm.nih.gov/pubmed/20300017[↩]
- Witard OC, Jackman SR, Breen L, Smith K, Selby A, Tipton KD. Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am J Clin Nutr. 2014;99:86–95. doi: 10.3945/ajcn.112.055517. https://www.ncbi.nlm.nih.gov/pubmed/24257722[↩]
- Yang Y, Breen L, Burd NA, Hector AJ, Churchward-Venne TA, Josse AR, et al. Resistance exercise enhances myofibrillar protein synthesis with graded intakes of whey protein in older men. Br J Nutr. 2012;108:1780–8. https://www.ncbi.nlm.nih.gov/pubmed/22313809[↩]
- Phillips SM. A brief review of higher dietary protein diets in weight loss: a focus on athletes. Sports Med. 2014;44(Suppl 2):S149–S153. doi: 10.1007/s40279-014-0254-y. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4213385/[↩]
- Bohe J, Low JF, Wolfe RR, Rennie MJ. Latency and duration of stimulation of human muscle protein synthesis during continuous infusion of amino acids. J Physiol. 2001;532(Pt 2):575–579. doi: 10.1111/j.1469-7793.2001.0575f.x. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2278544/[↩]
- Atherton PJ, Etheridge T, Watt PW, Wilkinson D, Selby A, Rankin D, et al. Muscle full effect after oral protein: time-dependent concordance and discordance between human muscle protein synthesis and mtorc1 signaling. Am J Clin Nutr. 2010;92:1080–8. https://www.ncbi.nlm.nih.gov/pubmed/20844073[↩]
- Kim IY, Schutzler S, Schrader A, Spencer HJ, Azhar G, Ferrando AA, et al. The anabolic response to a meal containing different amounts of protein is not limited by the maximal stimulation of protein synthesis in healthy young adults. Am J Physiol Endocrinol Metab. 2016;310:E73–80. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4675798/[↩]
- Fahs CA, Heffernan KS, Fernhall B. Hemodynamic and vascular response to resistance exercise with L-arginine. Med Sci Sports Exerc. 2009;41:773–779. doi: 10.1249/MSS.0b013e3181909d9d. https://www.ncbi.nlm.nih.gov/pubmed/19276857[↩]
- Tang JE, Lysecki PJ, Manolakos JJ, MacDonald MJ, Tarnopolsky MA, Phillips SM. Bolus arginine supplementation affects neither muscle blood flow nor muscle protein synthesis in young men at rest or after resistance exercise. J Nutr. 2011;141:195–200. doi: 10.3945/jn.110.130138. https://www.ncbi.nlm.nih.gov/pubmed/21191143[↩]
- Volpi E, Kobayashi H, Sheffield-Moore M, Mittendorfer B, Wolfe RR. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am J Clin Nutr. 2003;78:250–258. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3192452/[↩]
- Alvares TS, Meirelles CM, Bhambhani YN, Paschoalin VM, Gomes PS. L-Arginine as a potential ergogenic aid in healthy subjects. Sports Med. 2011;41:233–248. doi: 10.2165/11538590-000000000-00000. https://www.ncbi.nlm.nih.gov/pubmed/21395365[↩]
- Greer BK, Jones BT. Acute arginine supplementation fails to improve muscle endurance or affect blood pressure responses to resistance training. J Strength Cond Res. 2011;25:1789–1794. doi: 10.1519/JSC.0b013e3181e07569. https://www.ncbi.nlm.nih.gov/pubmed/21399536[↩]
- McConell GK. Effects of L-arginine supplementation on exercise metabolism. Curr Opin Clin Nutr Metab Care. 2007;10:46–51. doi: 10.1097/MCO.0b013e32801162fa. https://www.ncbi.nlm.nih.gov/pubmed/17143054[↩]
- Shao A, Hathcock JN. Risk assessment for the amino acids taurine, L-glutamine and L-arginine. Regul Toxicol Pharmacol. 2008;50:376–399. doi: 10.1016/j.yrtph.2008.01.004. https://www.ncbi.nlm.nih.gov/pubmed/18325648[↩][↩]
- Perez-Guisado J, Jakeman PM. Citrulline malate enhances athletic anaerobic performance and relieves muscle soreness. J Strength Cond Res. 2010;24:1215–1222. doi: 10.1519/JSC.0b013e3181cb28e0. https://www.ncbi.nlm.nih.gov/pubmed/20386132[↩][↩]
- Stoppani J, Scheett T, Pena J, Rudolph C, Charlebois D. Consuming a supplement containing branched-chain amino acids during a resistance-traning program increases lean mass, muscle strength, and fat loss. J Int Soc Sports Nutr. 2009;6:P1. doi: 10.1186/1550-2783-6-S1-P1.[↩][↩]
- Sureda A, Cordova A, Ferrer MD, Perez G, Tur JA, Pons A. L-citrulline-malate influence over branched chain amino acid utilization during exercise. Eur J Appl Physiol. 2010;110:341–351. doi: 10.1007/s00421-010-1509-4. https://www.ncbi.nlm.nih.gov/pubmed/20499249[↩]
- Hickner RC, Tanner CJ, Evans CA, Clark PD, Haddock A, Fortune C, Geddis H, Waugh W, McCammon M. L-citrulline reduces time to exhaustion and insulin response to a graded exercise test. Med Sci Sports Exerc. 2006;38:660–666. doi: 10.1249/01.mss.0000210197.02576.da. https://www.ncbi.nlm.nih.gov/pubmed/16679980[↩]
- Antonio J, Sanders MS, Kalman D, Woodgate D, Street C. The effects of high-dose glutamine ingestion on weightlifting performance. J Strength Cond Res. 2002;16:157–160. https://www.ncbi.nlm.nih.gov/pubmed/11834123[↩]
- Haub MD, Potteiger JA, Nau KL, Webster MJ, Zebas CJ. Acute L-glutamine ingestion does not improve maximal effort exercise. J Sports Med Phys Fitness. 1998;38:240–244. https://www.ncbi.nlm.nih.gov/pubmed/9830832[↩][↩]
- Colker CM, Swain MA, Fabrucini B, Shi Q, Kalman DS. Effects of supplemental protein on body composition and muscular strength in healthy athletic male adults. Curr Ther Res. 2000;61:19–28. doi: 10.1016/S0011-393X(00)88492-1.[↩]
- Candow DG, Chilibeck PD, Burke DG, Davison KS, Smith-Palmer T. Effect of glutamine supplementation combined with resistance training in young adults. Eur J Appl Physiol. 2001;86:142–149. doi: 10.1007/s00421-001-0523-y. https://www.ncbi.nlm.nih.gov/pubmed/11822473[↩]
- Camilleri M, Madsen K, Spiller R, Van Meerveld BG, Verne GN. Intestinal barrier function in health and gastrointestinal disease. Neurogastroenterol Motil. 2012;24:503–512. doi: 10.1111/j.1365-2982.2012.01921.x. https://www.ncbi.nlm.nih.gov/pubmed/22583600[↩]
- Emerging health properties of whey proteins and their clinical implications. Krissansen GW. J Am Coll Nutr. 2007 Dec; 26(6):713S-23S. https://www.ncbi.nlm.nih.gov/pubmed/18187438/[↩]
- Invited review: physiological properties of bioactive peptides obtained from whey proteins. Madureira AR, Tavares T, Gomes AM, Pintado ME, Malcata FX. J Dairy Sci. 2010 Feb; 93(2):437-55. https://www.ncbi.nlm.nih.gov/pubmed/20105516/[↩]
- Protective effect of whey protein hydrolysates on H₂O₂-induced PC12 cells oxidative stress via a mitochondria-mediated pathway. Jin MM, Zhang L, Yu HX, Meng J, Sun Z, Lu RR. Food Chem. 2013 Nov 15; 141(2):847-52. https://www.ncbi.nlm.nih.gov/pubmed/23790857/[↩]
- Whey protein isolate counteracts the effects of a high-fat diet on energy intake and hypothalamic and adipose tissue expression of energy balance-related genes. McAllan L, Keane D, Schellekens H, Roche HM, Korpela R, Cryan JF, Nilaweera KN. Br J Nutr. 2013 Dec 14; 110(11):2114-26. https://www.ncbi.nlm.nih.gov/pubmed/23731955/[↩]
- Effect of whey protein hydrolysates with different molecular weight on fatigue induced by swimming exercise in mice. Liu J, Wang X, Zhao Z. J Sci Food Agric. 2014 Jan 15; 94(1):126-30. https://www.ncbi.nlm.nih.gov/pubmed/23653310/[↩]
- Biochemical and metabolic mechanisms by which dietary whey protein may combat obesity and Type 2 diabetes. Jakubowicz D, Froy O. J Nutr Biochem. 2013 Jan; 24(1):1-5. https://www.ncbi.nlm.nih.gov/pubmed/22995389/[↩]
- The role of leucine in the regulation of protein metabolism. Garlick PJ. J Nutr. 2005 Jun; 135(6 Suppl):1553S-6S. https://www.ncbi.nlm.nih.gov/pubmed/15930468/[↩]
- Post-exercise carbohydrate plus whey protein hydrolysates supplementation increases skeletal muscle glycogen level in rats. Morifuji M, Kanda A, Koga J, Kawanaka K, Higuchi M. Amino Acids. 2010 Apr; 38(4):1109-15. https://www.ncbi.nlm.nih.gov/pubmed/19593593/[↩]
- Effect of soy- and whey protein-isolate supplemented diet on the redox parameters of trained mice. Elia D, Stadler K, Horváth V, Jakus J. Eur J Nutr. 2006 Aug; 45(5):259-66. https://www.ncbi.nlm.nih.gov/pubmed/16575496/[↩]
- Effects of the dietary amount and source of protein, resistance training and anabolic-androgenic steroids on body weight and lipid profile of rats. Aparicio VA, Sánchez C, Ortega FB, Nebot E, Kapravelou G, Porres JM, Aranda P. Nutr Hosp. 2013 Jan-Feb; 28(1):127-36. https://www.ncbi.nlm.nih.gov/pubmed/23808440/[↩]
- A dipeptide and an amino acid present in whey protein hydrolysate increase translocation of GLUT-4 to the plasma membrane in Wistar rats. Morato PN, Lollo PC, Moura CS, Batista TM, Carneiro EM, Amaya-Farfan J. Food Chem. 2013 Aug 15; 139(1-4):853-9. https://www.ncbi.nlm.nih.gov/pubmed/23561181/[↩]
- Whey proteins are more efficient than casein in the recovery of muscle functional properties following a casting induced muscle atrophy. Martin V, Ratel S, Siracusa J, Le Ruyet P, Savary-Auzeloux I, Combaret L, Guillet C, Dardevet D. PLoS One. 2013; 8(9):e75408. https://www.ncbi.nlm.nih.gov/pubmed/24069411/[↩]
- Whey protein supplementation during resistance training augments lean body mass. Volek JS, Volk BM, Gómez AL, Kunces LJ, Kupchak BR, Freidenreich DJ, Aristizabal JC, Saenz C, Dunn-Lewis C, Ballard KD, Quann EE, Kawiecki DL, Flanagan SD, Comstock BA, Fragala MS, Earp JE, Fernandez ML, Bruno RS, Ptolemy AS, Kellogg MD, Maresh CM, Kraemer WJ. J Am Coll Nutr. 2013; 32(2):122-35. https://www.ncbi.nlm.nih.gov/pubmed/24015719/[↩]
- Isoleucine and leucine independently regulate mTOR signaling and protein synthesis in MAC-T cells and bovine mammary tissue slices. Appuhamy JA, Knoebel NA, Nayananjalie WA, Escobar J, Hanigan MD. J Nutr. 2012 Mar; 142(3):484-91. https://www.ncbi.nlm.nih.gov/pubmed/22298573/[↩]
- Whey protein hydrolysate augments tendon and muscle hypertrophy independent of resistance exercise contraction mode. Farup J, Rahbek SK, Vendelbo MH, Matzon A, Hindhede J, Bejder A, Ringgard S, Vissing K. Scand J Med Sci Sports. 2014 Oct; 24(5):788-98. https://www.ncbi.nlm.nih.gov/pubmed/23647357/[↩]
- In vitro and in vivo safety studies of a proprietary whey extract. Dyer AR, Burdock GA, Carabin IG, Haas MC, Boyce J, Alsaker R, Read LC. Food Chem Toxicol. 2008 May; 46(5):1659-65. https://www.ncbi.nlm.nih.gov/pubmed/18295388/[↩]
- Acute and repeated dose (28 days) oral safety studies of ALIBIRD in rats. Anadón A, Martínez MA, Ares I, Castellano V, Martínez-Larrañaga MR, Corzo N, Olano A, Montilla A, Recio I, Martínez-Maqueda D, Miralles B, Fornari T, García-Risco MR, Gonzalez M, Reglero G. J Food Prot. 2013 Jul; 76(7):1226-39. https://www.ncbi.nlm.nih.gov/pubmed/23834798/[↩]
- CHEN W-C, HUANG W-C, CHIU C-C, CHANG Y-K, HUANG C-C. Whey Protein Improves Exercise Performance and Biochemical Profiles in Trained Mice. Medicine and Science in Sports and Exercise. 2014;46(8):1517-1524. doi:10.1249/MSS.0000000000000272. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4186725/[↩]
- Nutritional physiology of whey and whey components. Barth CA, Behnke U. Nahrung. 1997 Feb; 41(1):2-12. https://www.ncbi.nlm.nih.gov/pubmed/9157293/[↩]
- Dietary whey protein lessens several risk factors for metabolic diseases: a review. Sousa GT, Lira FS, Rosa JC, de Oliveira EP, Oyama LM, Santos RV, Pimentel GD. Lipids Health Dis. 2012 Jul 10; 11():67. https://www.ncbi.nlm.nih.gov/pubmed/22676328/[↩]
- Antimicrobial activity of whey protein isolate edible films with essential oils against food spoilers and foodborne pathogens. Fernández-Pan I, Royo M, Ignacio Maté J. J Food Sci. 2012 Jul; 77(7):M383-90. https://www.ncbi.nlm.nih.gov/pubmed/22671770/[↩]
- Tipton KD, Ferrando AA, Phillips SM, Doyle D, Jr, Wolfe RR. Postexercise net protein synthesis in human muscle from orally administered amino acids. Am J Phys. 1999;276(4 Pt 1):E628–E634. https://www.ncbi.nlm.nih.gov/pubmed/10198297[↩]
- Burd NA, West DW, Moore DR, Atherton PJ, Staples AW, Prior T, et al. Enhanced amino acid sensitivity of myofibrillar protein synthesis persists for up to 24 h after resistance exercise in young men. J Nutr. 2011;141:568–73. https://www.ncbi.nlm.nih.gov/pubmed/21289204[↩]
- Tipton KD, Gurkin BE, Matin S, Wolfe RR. Nonessential amino acids are not necessary to stimulate net muscle protein synthesis in healthy volunteers. J Nutr Biochem. 1999;10:89–95. doi: 10.1016/S0955-2863(98)00087-4. https://www.ncbi.nlm.nih.gov/pubmed/15539275[↩]
- Tipton KD, Rasmussen BB, Miller SL, Wolf SE, Owens-Stovall SK, Petrini BE, et al. Timing of amino acid-carbohydrate ingestion alters anabolic response of muscle to resistance exercise. Am J Physiol Endocrinol Metab. 2001;281:E197–206. https://www.ncbi.nlm.nih.gov/pubmed/11440894[↩][↩]
- Tipton KD, Borsheim E, Wolf SE, Sanford AP, Wolfe RR. Acute response of net muscle protein balance reflects 24-h balance after exercise and amino acid ingestion. Am J Physiol Endocrinol Metab. 2003;284:E76–E89. doi: 10.1152/ajpendo.00234.2002. https://www.ncbi.nlm.nih.gov/pubmed/12388164[↩]
- Kerksick C, Harvey T, Stout J, Campbell B, Wilborn C, Kreider R, et al. International society of sports nutrition position stand: nutrient timing. J Int Soc Sports Nutr. 2008;5:17. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2575187/[↩]
- Farnfield MM, Breen L, Carey KA, Garnham A, Cameron-Smith D. Activation of mtor signalling in young and old human skeletal muscle in response to combined resistance exercise and whey protein ingestion. Appl Physiol Nutr Metab. 2012;37:21–30. doi: 10.1139/h11-132. https://www.ncbi.nlm.nih.gov/pubmed/22148961[↩]
- Hulmi JJ, Kovanen V, Lisko I, Selanne H, Mero AA. The effects of whey protein on myostatin and cell cycle-related gene expression responses to a single heavy resistance exercise bout in trained older men. Eur J Appl Physiol. 2008;102:205–213. doi: 10.1007/s00421-007-0579-4. https://www.ncbi.nlm.nih.gov/pubmed/17924133[↩]
- Churchward-Venne TA, Murphy CH, Longland TM, Phillips SM. Role of protein and amino acids in promoting lean mass accretion with resistance exercise and attenuating lean mass loss during energy deficit in humans. Amino Acids. 2013;45:231–240. doi: 10.1007/s00726-013-1506-0. https://www.ncbi.nlm.nih.gov/pubmed/23645387[↩]
- Pennings B, Koopman R, Beelen M, Senden JM, Saris WH, Van Loon LJ. Exercising before protein intake allows for greater use of dietary protein-derived amino acids for de novo muscle protein synthesis in both young and elderly men. Am J Clin Nutr. 2011;93:322–331. doi: 10.3945/ajcn.2010.29649. https://www.ncbi.nlm.nih.gov/pubmed/21084649[↩]
- Fujita S, Dreyer HC, Drummond MJ, Glynn EL, Volpi E, Rasmussen BB. Essential amino acid and carbohydrate ingestion before resistance exercise does not enhance postexercise muscle protein synthesis. J Appl Physiol (1985) 2009;106:1730–1739. doi: 10.1152/japplphysiol.90395.2008. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2681328/[↩]
- Burk A, Timpmann S, Medijainen L, Vahi M, Oopik V. Time-divided ingestion pattern of casein-based protein supplement stimulates an increase in fat-free body mass during resistance training in young untrained men. Nutr Res. 2009;29:405–413. doi: 10.1016/j.nutres.2009.03.008. https://www.ncbi.nlm.nih.gov/pubmed/19628107[↩][↩]
- Aragon AA, Schoenfeld BJ. Nutrient timing revisited: is there a post-exercise anabolic window? J Int Soc Sports Nutr. 2013;10:5. doi: 10.1186/1550-2783-10-5. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3577439/[↩][↩]
- Bosse JD, Dixon BM. Dietary protein to maximize resistance training: a review and examination of protein spread and change theories. J Int Soc Sports Nutr. 2012;9:42. doi: 10.1186/1550-2783-9-42. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3518828/[↩]
- Macnaughton LS, Wardle SL, Witard OC, Mcglory C, Hamilton DL, Jeromson S, et al. The response of muscle protein synthesis following whole-body resistance exercise is greater following 40 g than 20 g of ingested whey protein. Physiol Rep. 2016;4:15. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4985555/[↩][↩]
- West DW, Burd NA, Coffey VG, Baker SK, Burke LM, Hawley JA, et al. Rapid aminoacidemia enhances myofibrillar protein synthesis and anabolic intramuscular signaling responses after resistance exercise. Am J Clin Nutr. 2011;94:795–803. https://www.ncbi.nlm.nih.gov/pubmed/21795443[↩]
- Res PT, Groen B, Pennings B, Beelen M, Wallis GA, Gijsen AP, et al. Protein ingestion before sleep improves postexercise overnight recovery. Med Sci Sports Exerc. 2012;44:1560–9. https://www.ncbi.nlm.nih.gov/pubmed/22330017[↩]
- Madzima TA, Panton LB, Fretti SK, Kinsey AW, Ormsbee MJ. Night-time consumption of protein or carbohydrate results in increased morning resting energy expenditure in active college-aged men. Br J Nutr. 2014;111:71–77. doi: 10.1017/S000711451300192X. https://www.ncbi.nlm.nih.gov/pubmed/23768612[↩][↩][↩]
- Kinsey AW, Cappadona SR, Panton LB, Allman BR, Contreras RJ, Hickner RC, et al. The effect of casein protein prior to sleep on fat metabolism in obese men. Nutrients. 2016;8:8. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4997367/[↩][↩]
- Ormsbee MJ, Kinsey AW, Eddy WR, Madzima TA, Arciero PJ, Figueroa A, et al. The influence of nighttime feeding of carbohydrate or protein combined with exercise training on appetite and cardiometabolic risk in young obese women. Appl Physiol Nutr Metab. 2015;40:37–45. https://www.ncbi.nlm.nih.gov/pubmed/25409324[↩]
- Figueroa A, Wong A, Kinsey A, Kalfon R, Eddy W, Ormsbee MJ. Effects of milk proteins and combined exercise training on aortic hemodynamics and arterial stiffness in young obese women with high blood pressure. Am J Hypertens. 2014;27:338–344. doi: 10.1093/ajh/hpt224. https://www.ncbi.nlm.nih.gov/pubmed/24300595[↩]
- Snijders T, Res PT, Smeets JS, Van Vliet S, Van Kranenburg J, Maase K, et al. Protein ingestion before sleep increases muscle mass and strength gains during prolonged resistance-type exercise training in healthy young men. J Nutr. 2015;145:1178–84. https://www.ncbi.nlm.nih.gov/pubmed/25926415[↩]
- Antonio J, Ellerbroek A, Peacock C, Silver T. Casein protein supplementation in trained men and women: morning versus evening. Int J Exerc Sci. 2017;10:479–486. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5421981/[↩][↩]
- Buckner SL, Leonneke JP, Loprinzi PD. Protein timing during the day and its relevance for muscle strength and lean mass. Clin Physiol Funct Imaging. doi: 10.1111/cpf.12440. https://www.ncbi.nlm.nih.gov/pubmed/28474785[↩]
- Paddon-Jones D, Sheffield-Moore M, Urban RJ, Sanford AP, Aarsland A, Wolfe RR, et al. Essential amino acid and carbohydrate supplementation ameliorates muscle protein loss in humans during 28 days bedrest. J Clin Endocrinol Metab. 2004;89:4351–8. https://www.ncbi.nlm.nih.gov/pubmed/15356032[↩][↩][↩]
- Mitchell CJ, Churchward-Venne TA, Parise G, Bellamy L, Baker SK, Smith K, et al. Acute post-exercise myofibrillar protein synthesis is not correlated with resistance training-induced muscle hypertrophy in young men. PLoS One. 2014;9:e89431. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3933567/[↩]
- Areta JL, Burke LM, Ross ML, Camera DM, West DW, Broad EM, et al. Timing and distribution of protein ingestion during prolonged recovery from resistance exercise alters myofibrillar protein synthesis. J Physiol. 2013;591:2319–31. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3650697/[↩]
- Arnal MA, Mosoni L, Boirie Y, Houlier ML, Morin L, Verdier E, et al. Protein feeding pattern does not affect protein retention in young women. J Nutr. 2000;130:1700–4. https://www.ncbi.nlm.nih.gov/pubmed/10867039[↩]
- Tinsley GM, Forsse JS, Butler NK, Paoli A, Bane AA, La Bounty PM, et al. Time-restricted feeding in young men performing resistance training: a randomized controlled trial. Eur J Sport Sci. 2017;17:200–7. https://www.ncbi.nlm.nih.gov/pubmed/27550719[↩]
- International Society of Sports Nutrition position stand: nutrient timing. Kerksick C, Harvey T, Stout J, Campbell B, Wilborn C, Kreider R, Kalman D, Ziegenfuss T, Lopez H, Landis J, Ivy JL, Antonio J. J Int Soc Sports Nutr. 2008 Oct 3; 5():17. https://www.ncbi.nlm.nih.gov/pubmed/18834505/[↩]
- Helms ER, Aragon AA, Fitschen PJ. Evidence-based recommendations for natural bodybuilding contest preparation: nutrition and supplementation. Journal of the International Society of Sports Nutrition. 2014;11:20. doi:10.1186/1550-2783-11-20. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4033492/[↩]
- Nutrient intake, body fat, and lipid profiles of competitive male and female bodybuilders. Bazzarre TL, Kleiner SM, Litchford MD. J Am Coll Nutr. 1990 Apr; 9(2):136-42. https://www.ncbi.nlm.nih.gov/pubmed/2338462/[↩]
- Nutritional status of nationally ranked elite bodybuilders. Kleiner SM, Bazzarre TL, Ainsworth BE. Int J Sport Nutr. 1994 Mar; 4(1):54-69. https://www.ncbi.nlm.nih.gov/pubmed/8167655/[↩][↩]
- Food selection patterns of bodybuilders. Sandoval WM, Heyward VH. Int J Sport Nutr. 1991 Mar; 1(1):61-8. https://www.ncbi.nlm.nih.gov/pubmed/1844403/[↩]
- Improved cycling time-trial performance after ingestion of a caffeine energy drink. Ivy JL, Kammer L, Ding Z, Wang B, Bernard JR, Liao YH, Hwang J. Int J Sport Nutr Exerc Metab. 2009 Feb; 19(1):61-78. https://www.ncbi.nlm.nih.gov/pubmed/19403954/[↩]
- The effects of caffeine ingestion on time trial cycling performance. McNaughton LR, Lovell RJ, Siegler J, Midgley AW, Moore L, Bentley DJ. Int J Sports Physiol Perform. 2008 Jun; 3(2):157-63. https://www.ncbi.nlm.nih.gov/pubmed/19208924/[↩]
- Carr A, Dawson B, Schneiker K, Goodman C, Lay B. Effect of caffeine supplementation on repeated sprint running performance. J Sports Med Phys Fitness. 2008;48:472–478. https://www.ncbi.nlm.nih.gov/pubmed/18997650[↩]
- Glaister M, Howatson G, Abraham CS, Lockey RA, Goodwin JE, Foley P, McInnes G. Caffeine supplementation and multiple sprint running performance. Med Sci Sports Exerc. 2008;40:1835–1840. doi: 10.1249/MSS.0b013e31817a8ad2. https://www.ncbi.nlm.nih.gov/pubmed/18799995[↩]
- Green JM, Wickwire PJ, McLester JR, Gendle S, Hudson G, Pritchett RC, Laurent CM. Effects of caffeine on repetitions to failure and ratings of perceived exertion during resistance training. Int J Sports Physiol Perform. 2007;2:250–259. https://www.ncbi.nlm.nih.gov/pubmed/19168925[↩]
- Woolf K, Bidwell WK, Carlson AG. The effect of caffeine as an ergogenic aid in anaerobic exercise. Int J Sport Nutr Exerc Metab. 2008;18:412–429. https://www.ncbi.nlm.nih.gov/pubmed/18708685[↩]
- Duncan MJ, Oxford SW. The effect of caffeine ingestion on mood state and bench press performance to failure. J Strength Cond Res. 2011;25:178–185. doi: 10.1519/JSC.0b013e318201bddb. https://www.ncbi.nlm.nih.gov/pubmed/21157384[↩]
- Williams AD, Cribb PJ, Cooke MB, Hayes A. The effect of ephedra and caffeine on maximal strength and power in resistance-trained athletes. J Strength Cond Res. 2008;22:464–470. doi: 10.1519/JSC.0b013e3181660320. https://www.ncbi.nlm.nih.gov/pubmed/18550961[↩]
- Hendrix CR, Housh TJ, Mielke M, Zuniga JM, Camic CL, Johnson GO, Schmidt RJ, Housh DJ. Acute effects of a caffeine-containing supplement on bench press and leg extension strength and time to exhaustion during cycle ergometry. J Strength Cond Res. 2010;24:859–865. doi: 10.1519/JSC.0b013e3181ae7976. https://www.ncbi.nlm.nih.gov/pubmed/19834348[↩]
- Nawrot P, Jordan S, Eastwood J, Rotstein J, Hugenholtz A, Feeley M. Effects of caffeine on human health. Food Addit Contam. 2003;20:1–30. https://www.ncbi.nlm.nih.gov/pubmed/12519715[↩]
- Tarnopolsky MA, Atkinson SA, MacDougall JD, Sale DG, Sutton JR. Physiological responses to caffeine during endurance running in habitual caffeine users. Med Sci Sports Exerc. 1989;21:418–424. https://www.ncbi.nlm.nih.gov/pubmed/2674593[↩]
- Artioli GG, Gualano B, Smith A, Stout J, Lancha AH Jr. Role of beta-alanine supplementation on muscle carnosine and exercise performance. Med Sci Sports Exerc. 2010;42:1162–1173. https://www.ncbi.nlm.nih.gov/pubmed/20479615[↩]
- Harris RC, Tallon MJ, Dunnett M, Boobis L, Coakley J, Kim HJ, Fallowfield JL, Hill CA, Sale C, Wise JA. The absorption of orally supplied beta-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids. 2006;30:279–289. doi: 10.1007/s00726-006-0299-9. https://www.ncbi.nlm.nih.gov/pubmed/16554972[↩][↩]
- Hobson RM, Saunders B, Ball G, Harris RC, Sale C. Effects of beta-alanine supplementation on exercise performance: a meta-analysis. . Amino Acids. 2012;43:25–37. doi: 10.1007/s00726-011-1200-z. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3374095/[↩]
- Lu P, Xu W, Sturman JA. Dietary beta-alanine results in taurine depletion and cerebellar damage in adult cats. J Neurosci Res. 1996;43:112–119. doi: 10.1002/jnr.490430115. https://www.ncbi.nlm.nih.gov/pubmed/8838582[↩]
- Smith HJ, Mukerji P, Tisdale MJ. Attenuation of proteasome-induced proteolysis in skeletal muscle by {beta}-hydroxy-{beta}-methylbutyrate in cancer-induced muscle loss. Cancer Res. 2005;65:277–283. https://www.ncbi.nlm.nih.gov/pubmed/15665304[↩]
- Eley HL, Russell ST, Baxter JH, Mukerji P, Tisdale MJ. Signaling pathways initiated by beta-hydroxy-beta-methylbutyrate to attenuate the depression of protein synthesis in skeletal muscle in response to cachectic stimuli. Am J Physiol Endocrinol Metab. 2007;293:E923–E931. doi: 10.1152/ajpendo.00314.2007. https://www.ncbi.nlm.nih.gov/pubmed/17609254[↩]
- Rathmacher JA, Nissen S, Panton L, Clark RH, Eubanks May P, Barber AE, D’Olimpio J, Abumrad NN. Supplementation with a combination of beta-hydroxy-beta-methylbutyrate (HMB), arginine, and glutamine is safe and could improve hematological parameters. JPEN J Parenter Enteral Nutr. 2004;28:65–75. doi: 10.1177/014860710402800265. https://www.ncbi.nlm.nih.gov/pubmed/15080599[↩][↩]
- Gallagher PM, Carrithers JA, Godard MP, Schulze KE, Trappe SW. Beta-hydroxy-beta-methylbutyrate ingestion, part II: effects on hematology, hepatic and renal function. Med Sci Sports Exerc. 2000;32:2116–2119. doi: 10.1097/00005768-200012000-00023. https://www.ncbi.nlm.nih.gov/pubmed/11128860[↩]
- Fitschen PJ, Wilson GJ, Wilson JM, Wilund KR. Efficacy of beta-hydroxy-beta-methylbutyrate supplementation in elderly and clinical populations. Nutrition. 2013;29:29–36. doi: 10.1016/j.nut.2012.05.005. https://www.ncbi.nlm.nih.gov/pubmed/23085015[↩]
- Wilson GJ, Wilson JM, Manninen AH. Effects of beta-hydroxy-beta-methylbutyrate (HMB) on exercise performance and body composition across varying levels of age, sex, and training experience: a review. Nutr Metab (Lond) 2008;5:1. doi: 10.1186/1743-7075-5-1. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2245953/[↩]
- Wilson J, Fitschen P, Campbell B, Wilson G, Zanchi N, Taylor L, Wilborn C, Kalman D, Stout J, Hoffman J, Ziegenfuss T, Lopez H, Kreider R, Smith-Ryan A, Antonio J. International Society of Sports Nutrition Position Stand: beta-hydroxy-beta-methylbutyrate (HMB) J Int Soc Sports Nutr. 2013;10:6. doi: 10.1186/1550-2783-10-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3568064/[↩]
- University of Maryland Medical Center. Creatine. http://www.umm.edu/health/medical/altmed/supplement/creatine[↩][↩][↩]
- Cooper R, Naclerio F, Allgrove J, Jimenez A. Creatine supplementation with specific view to exercise/sports performance: an update. Journal of the International Society of Sports Nutrition. 2012;9:33. doi:10.1186/1550-2783-9-33. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3407788/[↩][↩][↩][↩][↩]
- Clinical pharmacology of the dietary supplement creatine monohydrate. Persky AM, Brazeau GA. Pharmacol Rev. 2001 Jun; 53(2):161-76. https://www.ncbi.nlm.nih.gov/pubmed/11356982/[↩][↩][↩]
- Brunzel NA. Renal function: Nonprotein nitrogen compounds, function tests, and renal disease. In: Scardiglia J, Brown M, McCullough K, Davis K, editor. Clinical Chemistry. McGraw-Hill: New York, NY; 2003. pp. 373–399.[↩][↩][↩]
- Potential ergogenic effects of arginine and creatine supplementation. Paddon-Jones D, Børsheim E, Wolfe RR. J Nutr. 2004 Oct; 134(10 Suppl):2888S-2894S; discussion 2895S. http://jn.nutrition.org/content/134/10/2888S.long[↩]
- Creatine in humans with special reference to creatine supplementation. Balsom PD, Söderlund K, Ekblom B. Sports Med. 1994 Oct; 18(4):268-80. https://www.ncbi.nlm.nih.gov/pubmed/7817065/[↩][↩]
- Muscle creatine loading in men. Hultman E, Söderlund K, Timmons JA, Cederblad G, Greenhaff PL. J Appl Physiol (1985). 1996 Jul; 81(1):232-7. https://www.ncbi.nlm.nih.gov/pubmed/8828669/[↩]
- Buford TW, Kreider RB, Stout JR, et al. International Society of Sports Nutrition position stand: creatine supplementation and exercise. Journal of the International Society of Sports Nutrition. 2007;4:6. doi:10.1186/1550-2783-4-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2048496/[↩]
- The metabolic burden of creatine synthesis. Brosnan JT, da Silva RP, Brosnan ME. Amino Acids. 2011 May; 40(5):1325-31. https://www.ncbi.nlm.nih.gov/pubmed/21387089/[↩][↩]
- Hultman E, Bergstrom J, Spreit L, Soderlund K. Energy metabolism and fatigue. In: Taylor A, Gollnick PD, Green H, editor. Biochemistry of Exercise VII. Human Kinetics: Champaign, IL; 1990. pp. 73–92.[↩][↩][↩][↩]
- Greenhaff P. The nutritional biochemistry of creatine. J Nutrit Biochem. 1997;11:610–618. doi: 10.1016/S0955-2863(97)00116-2.[↩]
- Creatine supplementation and exercise performance: an update. Williams MH, Branch JD. J Am Coll Nutr. 1998 Jun; 17(3):216-34. https://www.ncbi.nlm.nih.gov/pubmed/9627907/[↩]
- Williams MH, Kreider R, Branch JD. Creatine: The power supplement. Champaign, IL: Human Kinetics Publishers; 1999. p. 252.[↩]
- Effect of creatine supplementation and resistance-exercise training on muscle insulin-like growth factor in young adults. Burke DG, Candow DG, Chilibeck PD, MacNeil LG, Roy BD, Tarnopolsky MA, Ziegenfuss T. Int J Sport Nutr Exerc Metab. 2008 Aug; 18(4):389-98. https://www.ncbi.nlm.nih.gov/pubmed/18708688/[↩]
- Chanutin A. The fate of creatine when administered to man. J Biol Chem. 1926;67:29–34.[↩][↩][↩]
- Buford T, Kreider R, Stout J, Greenwood M, Campbell B, Spano M, Ziegenfuss T, Lopez H, Landis J, Antonio J. International Society of Sports Nutrition position stand: creatine supplementation and exercise. J Int Soc Sports Nutr. 2007;4:6. doi: 10.1186/1550-2783-4-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2048496/[↩][↩][↩][↩]
- American College of Sport Medicine. Round Table, the physiological and health effects of oral creatine supplementation. Med Sci Sports Exc. 2000;32:706–717. https://www.ncbi.nlm.nih.gov/pubmed/10731017[↩]
- Branch JD. Effects of creatine supplementation on body composition and performace: a meta análisis. Int J Sports Nutr Exerc Metabol. 2003;13:I198-122. https://www.ncbi.nlm.nih.gov/pubmed/12945830[↩]
- Rawson ES, Volek JS. Effects of creatine supplementation and resistance training on muscle strength and weightlifting performance. J Strength Cond Res. 2003;17:822–831. https://www.ncbi.nlm.nih.gov/pubmed/14636102[↩][↩]
- Volek JS, Kraemer WJ. Creatine suplemetation: its effects on human muscular performance and body composition. J Strength Cond Res. 1996;10:200–210.[↩]
- Syrotuik DG, Bell GJ. Acute creatine monohydrate supplementation: a descriptive physiological profile of responders vs. nonresponders. J Strength Cond Res. 2004;18:610–617. https://www.ncbi.nlm.nih.gov/pubmed/15320650[↩]
- Greenhaff PL, Bodin K, Soderlund K, Hultman E. Effect of oral creatine supplementation on skeletal muscle phosphocreatine resynthesis. Am J Physiol. 1994;266:E725–E730. https://www.ncbi.nlm.nih.gov/pubmed/8203511[↩]
- Casey A, Greenhaff P. Does dietary creatine supplementation play a role in skeletal muscle metabolism and performance? Am J Clin Nutr. 2000;72:607S–617S. https://www.ncbi.nlm.nih.gov/pubmed/10919967[↩][↩][↩]
- Cribb PJ, Williams AD, Hayes A. A creatine-protein-carbohydrate supplement enhances responses to resistance training. Med Sci Sports Exerc. 2007;39:1960–1968. doi: 10.1249/mss.0b013e31814fb52a. https://www.ncbi.nlm.nih.gov/pubmed/17986903[↩][↩]
- Parise G, Mihic S, MacLennan D, Yarasheski KE, Tarnopolsky MA. Effects of acute creatine monohydrate supplementation on leucine kinetics and mixed-muscle protein synthesis. J Appl Physiol. 2001;91:1041–1047. https://www.ncbi.nlm.nih.gov/pubmed/11509496[↩]
- Louis M, Poortmans JR, Francaux M, Hultman E, Berre J, Boisseau N, Young VR, Smith K, Meier-Augenstein W, Babraj JA. et al. Creatine supplementation has no effect on human muscle protein turnover at rest in the postabsorptive or fed states. Am J Physiol Endocrinol Metab. 2003;284:E764–E770. https://www.ncbi.nlm.nih.gov/pubmed/12475751[↩]
- Deldicque L, Atherton P, Patel R, Theisen D, Nielens H, Rennie M, Francaux M. Effects of resistance exercise with and without creatine supplementation on gene expression and cell signaling in human skeletal muscle. J Appl Physiol. 2008;104:371–378. https://www.ncbi.nlm.nih.gov/pubmed/18048590[↩]
- Burke DG, Candow DG, Chilibeck PD, MacNeil LG, Roy BD, Tarnopolsky MA, Ziegenfuss T. Effect of creatine supplementation and resistance-exercise training on muscle insulin-like growth factor in young adults. Int J Sport Nutr Exerc Metab. 2008;18:389–398. https://www.ncbi.nlm.nih.gov/pubmed/18708688[↩][↩]
- Harp JB, Goldstein S, Phillips LS. Nutrition and somatomedin. XXIII. Molecular regulation of IGF-I by amino acid availability in cultured hepatocytes. Diabetes. 1991;40:95–101. doi: 10.2337/diabetes.40.1.95. https://www.ncbi.nlm.nih.gov/pubmed/1901809[↩]
- Op ‘t Eijnde B, Urso B, Richter EA, Greenhaff PL, Hespel P. Effect of oral creatine supplementation on human muscle GLUT4 protein content after immobilization. Diabetes. 2001;50:18–23. doi: 10.2337/diabetes.50.1.18. https://www.ncbi.nlm.nih.gov/pubmed/11147785[↩][↩][↩]
- Cooke MB, Rybalka E, Williams AD, Cribb PJ, Hayes A. Creatine supplementation enhances muscle force recovery after eccentrically-induced muscle damage in healthy individuals. J Int Soc Sports Nutr. 2009;6:13. doi: 10.1186/1550-2783-6-13. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2697134/[↩]
- Lawler JM, Barnes WS, Wu G, Song W, Demaree S. Direct antioxidant properties of creatine. Biochem Biophys Res Commun. 2002;290:47–52. doi: 10.1006/bbrc.2001.6164. https://www.ncbi.nlm.nih.gov/pubmed/11779131[↩][↩]
- Sestili P, Martinelli C, Bravi G, Piccoli G, Curci R, Battistelli M, Falcieri E, Agostini D, Gioacchini AM, Stocchi V. Creatine supplementation affords cytoprotection in oxidatively injured cultured mammalian cells via direct antioxidant activity. Free Radic Biol Med. 2006;40:837–849. doi: 10.1016/j.freeradbiomed.2005.10.035. https://www.ncbi.nlm.nih.gov/pubmed/16520236[↩]
- Rahimi R. Creatine supplementation decreases oxidative DNA damage and lipid peroxidation induced by a single bout of resistance exercise. J Strength Cond Res. 2011;25:3448–3455. doi: 10.1519/JSC.0b013e3182162f2b. https://www.ncbi.nlm.nih.gov/pubmed/22080314[↩]
- Hespel P, Derave W. Ergogenic effects of creatine in sports and rehabilitation. Subcell Biochem. 2007;46:245–259. doi: 10.1021/bi061646s. https://www.ncbi.nlm.nih.gov/pubmed/18652080[↩][↩][↩]
- Nelson A, Arnall D, Kokkonen J, Day R, Evans J. Muscle glycogen supercompensation is enhanced by prior creatine supplementation. Med Sci Sports Exerc. 2001;33:1096–1100. https://www.ncbi.nlm.nih.gov/pubmed/11445755[↩]
- Sewell D, Robinson T, Greenhaff P. Creatine supplementation does not affect human skeletal muscle glycogen content in the absence of prior exercise. J Appl Physiol. 2008;104:508–512. https://www.ncbi.nlm.nih.gov/pubmed/18032580[↩]
- Hickner R, Dyck D, Sklar J, Hatley H, Byrd P. Effect of 28 days of creatine ingestion on muscle metabolism and performance of a simulated cycling road race. J Int Soc Sports Nutr. 2010;7:26. doi: 10.1186/1550-2783-7-26. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2909923/[↩][↩][↩]
- Branch J. Effect of creatine supplementation on body composition and performance: a meta-analysis. Int J Sport Nutr Exerc Metab. 2003;13:198–226. https://www.ncbi.nlm.nih.gov/pubmed/12945830[↩][↩][↩][↩][↩][↩]
- Chwalbiñska-Moneta J. Effect of creatine supplementation on aerobic performance and anaerobic capacity in elite rowers in the course of endurance training. Int J Sport Nutr Exerc Metab. 2003;13:173–183. https://www.ncbi.nlm.nih.gov/pubmed/12945828[↩][↩]
- Graef J, Smith A, Kendall K, Fukuda D, Moon J, Beck T, Cramer J, Stout J. The effects of four weeks of creatine supplementation and high-intensity interval training on cardiorespiratory fitness: a randomized controlled trial. J Int Soc Sports Nutr. 2009;6:18. doi: 10.1186/1550-2783-6-18. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2780977/[↩][↩]
- Thompson C, Kemp G, Sanderson A, Dixon R, Styles P, Taylor D, Radda G. Effect of creatine on aerobic and anaerobic metabolism in skeletal muscle in swimmers. Br J Sports Med. 1996;30:222–225. doi: 10.1136/bjsm.30.3.222. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1332335/[↩][↩]
- Volek J, Duncan N, Mazzetti S, Staron R, Putukian M, Gómez A, Pearson D, Fink W, Kraemer W. Performance and muscle fiber adaptations to creatine supplementation and heavy resistance training. Med Sci Sports Exerc. 1999;31:1147–1156. doi: 10.1097/00005768-199908000-00011. https://www.ncbi.nlm.nih.gov/pubmed/10449017[↩]
- Volek J, Rawson E. Scientific basis and practical aspects of creatine supplementation for athletes. Nutrition. 2004;20:609–614. doi: 10.1016/j.nut.2004.04.014. https://www.ncbi.nlm.nih.gov/pubmed/15212742[↩]
- van Loon L, Oosterlaar A, Hartgens F, Hesselink M, Snow R, Wagenmakers A. Effects of creatine loading and prolonged creatine supplementation on body composition, fuel selection, sprint and endurance performance in humans. Clin Sci (Lond) 2003;104:153–162. doi: 10.1042/CS20020159. https://www.ncbi.nlm.nih.gov/pubmed/12546637[↩]
- Kreider RB. Effects of creatine supplementation on performance and training adaptations. Mol Cell Biochem. 2003;244:89–94. doi: 10.1023/A:1022465203458. https://www.ncbi.nlm.nih.gov/pubmed/12701815[↩]
- Jakobi J, Rice C, Curtin S, Marsh G. Contractile properties, fatigue and recovery are not influenced by short-term creatine supplementation in human muscle. Exp Physiol. 2000;85:451–460. doi: 10.1017/S0958067000020212. https://www.ncbi.nlm.nih.gov/pubmed/10918084[↩]
- Bemben MG, Witten MS, Carter JM, Eliot KA, Knehans AW, Bemben DA. The effects of supplementation with creatine and protein on muscle strength following a traditional resistance training program in middle-aged and older men. J Nutr Health Aging. 2010;14:155–159. doi: 10.1007/s12603-009-0124-8. https://www.ncbi.nlm.nih.gov/pubmed/20126965[↩]
- Safdar A, Yardley N, Snow R, Melov S, Tarnopolsky M. Global and targeted gene expression and protein content in skeletal muscle of young men following short-term creatine monohydrate supplementation. Physiol Genomics. 2008;32:219–228. https://www.ncbi.nlm.nih.gov/pubmed/17957000[↩][↩]
- Saremi A, Gharakhanloo R, Sharghi S, Gharaati M, Larijani B, Omidfar K. Effects of oral creatine and resistance training on serum myostatin and GASP-1. Mol Cell Endocrinol. 2010;317:25–30. doi: 10.1016/j.mce.2009.12.019. https://www.ncbi.nlm.nih.gov/pubmed/20026378[↩]
- Bazzucchi I, Felici F, Sacchetti M. Effect of short-term creatine supplementation on neuromuscular function. Med Sci Sports Exerc. 2009;41:1934–41. doi: 10.1249/MSS.0b013e3181a2c05c. https://www.ncbi.nlm.nih.gov/pubmed/19727018[↩]
- Sculthorpe N, Grace F, Jones P, Fletcher I. The effect of short-term creatine loading on active range of movement. Appl Physiol Nutr Metab. 2010;35:507–511. doi: 10.1139/H10-036. https://www.ncbi.nlm.nih.gov/pubmed/20725117[↩][↩]
- Hile A, Anderson J, Fiala K, Stevenson J, Casa D, Maresh C. Creatine supplementation and anterior compartment pressure during exercise in the heat in dehydrated men. J Athl Train. 2006;41:30–35. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1421498/[↩]
- Steenge GR, Lambourne J, Casey A, Macdonald IA, Greenhaff PL. Stimulatory effect of insulin on creatine accumulation in human skeletal muscle. Am J Physiol. 1998;275:E974–E979. https://www.ncbi.nlm.nih.gov/pubmed/9843739[↩]
- Steenge G, Simpson E, Greenhaff P. Protein- and carbohydrate-induced augmentation of whole body creatine retention in humans. J Appl Physiol. 2000;89:1165–1171. https://www.ncbi.nlm.nih.gov/pubmed/10956365[↩]
- Olsen S, Aagaard P, Kadi F, Tufekovic G, Verney J, Olesen JL, Suetta C, Kjaer M. Creatine supplementation augments the increase in satellite cell and myonuclei number in human skeletal muscle induced by strength training. J Physiol. 2006;573:525–534. doi: 10.1113/jphysiol.2006.107359. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1779717/[↩]
- Walsh AL, Gonzalez AM, Ratamess NA, Kang J, Hoffman JR. Improved time to exhaustion following ingestion of the energy drink Amino Impact. J Int Soc Sports Nutr. 2010;7:14. doi: 10.1186/1550-2783-7-14. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2861014/[↩][↩]



{kind=link}