Contents
Small intestine
The small intestine is the longest part of the alimentary canal and it begins at the pyloric sphincter of the stomach, coils through the central and inferior part of the abdominal cavity, and eventually opens into the large intestine.
The term small intestine refers not to its length but to its diameter—about 2.5 cm (1 in.).
The small intestine is approximately 6 to 7 m long (range 3–7 m) with a narrowing diameter from beginning to end.
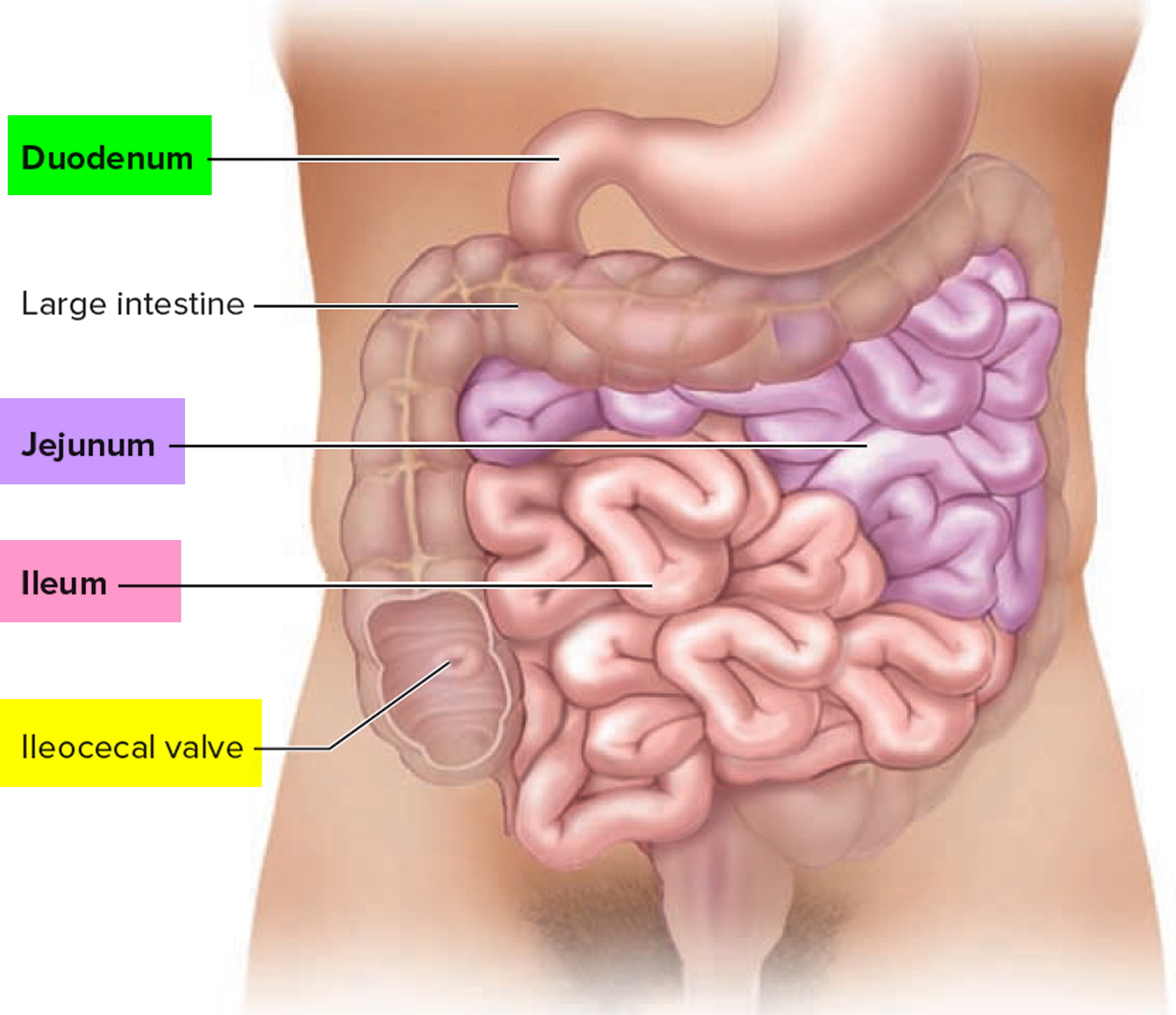
The small intestine consists of the duodenum, the jejunum, and the ileum.
The small intestine receives chyme from the stomach and secretions from the pancreas, liver, and gallbladder. It completes digestion of the nutrients in chyme, absorbs the products of digestion, and transports the residue to the large intestine.
The small intestine the site of most enzymatic digestion and about 90 percent of all nutrients absorption and most of the rest occurs in the proximal portion of the large intestine. Its length alone provides a large surface area for digestion and absorption, and that area is further increased by circular folds, villi, and microvilli. Most digestive enzymes that operate within the small intestine are secreted not by the intestine, but by the pancreas. During digestion, the small intestine undergoes active segmentation movements, shuffling the chyme back and forth and thereby maximizing its contact with the nutrient-absorbing mucosa.
Peristalsis propels chyme through the small intestine in about 3–6 hours.
Figure 1. Small intestine
Function of the Small Intestine
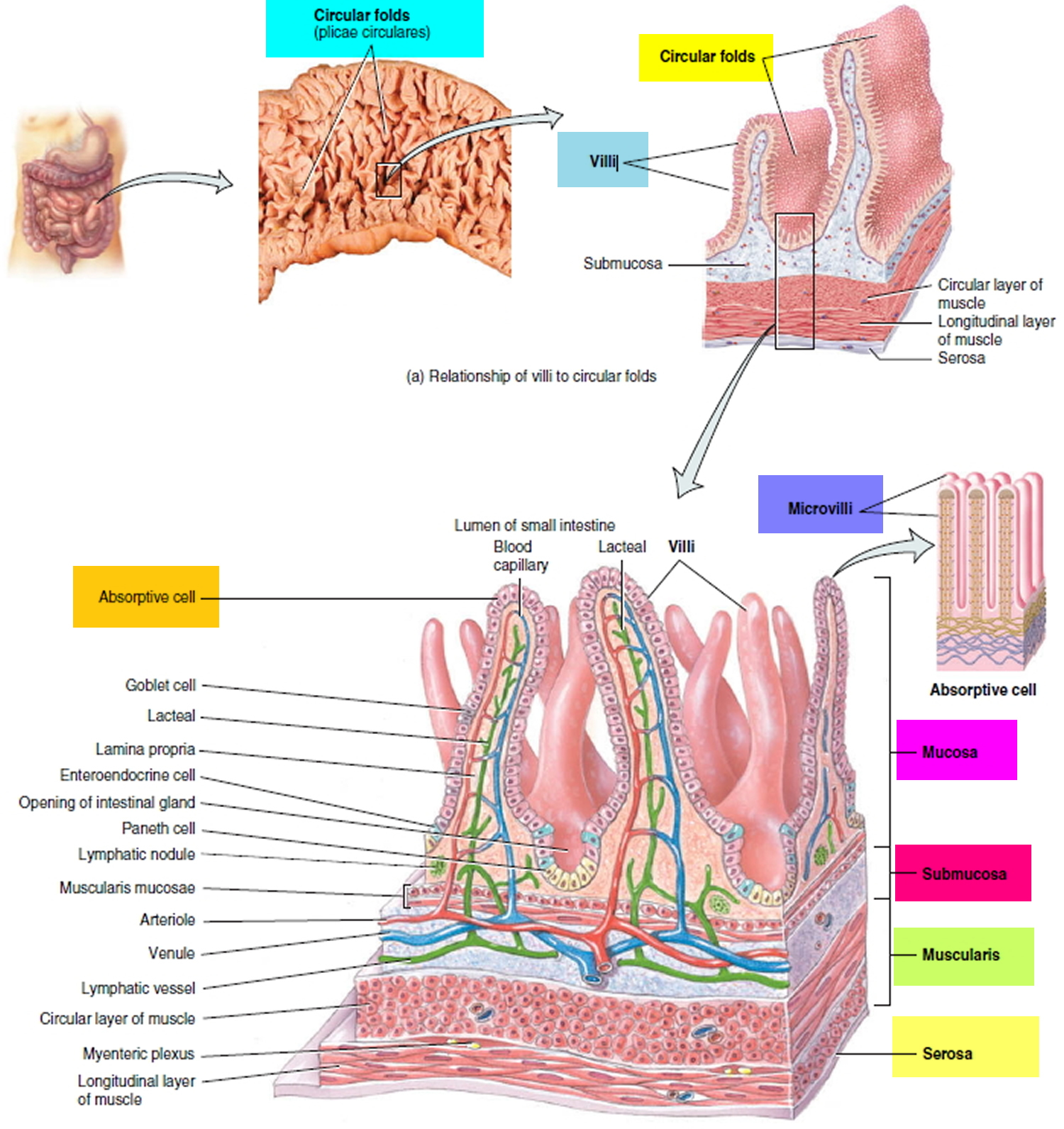
The anatomy of the small intestine is specialized to increase its surface area for absorption and secretion. The intestinal lining has a series of ring-shaped projections called circular folds or plicae circulares.
These folds do not disappear as the small intestine fills. Roughly 800 circular folds (about 2 per centimeter) are found along the length of the duodenum, jejunum, and proximal half of the ileum. The mucosa possesses intestinal villi, and each cell of the surface epithelium has small microvilli on its apical surface.
- Segmentations mix chyme with digestive juices and bring food into contact with mucosa for absorption; peristalsis propels chyme through small intestine.
- Completes digestion of carbohydrates, proteins, and lipids; begins and completes digestion of nucleic acids.
- Absorbs about 90% of nutrients and water that pass through digestive system.
Figure 2. Circular folds, villi and microvilli increase the surface area of the small intestine for digestion and absorption
The tissue layers of the small intestine are reminiscent of those in the esophagus and stomach with modifications appropriate for nutrient digestion and absorption. The lumen is lined with simple columnar epithelium. The muscularis externa is notable for a thick inner circular layer and a thinner outer longitudinal layer.
The jejunum and ileum are intraperitoneal and thus covered on all sides with a serosa, which is continuous with the complex, folded mesentery that suspends the small intestine from the posterior abdominal wall. Most of the duodenum is retroperitoneal and has a serosa only on its anterior surface; its other surfaces are covered by adventitia.
Effective digestion and absorption require the small intestine to have a large internal surface area. This is provided by its relatively great length and by three kinds of internal folds or projections: the circular folds, villi, and microvilli. If the mucosa were smooth, like the inside of a hose, it would have a surface area of about 0.3 to 0.5 m2, but with these surface elaborations, its actual surface area is about 200 m2—clearly a great advantage for nutrient absorption.
The circular folds increase the surface area by a factor of 2 to 3, the villi by a factor of 10, and the microvilli by a factor of 20.
Circular folds, the largest of these elaborations, are transverse to spiral ridges up to 1 cm high. These involve only the mucosa and submucosa; they are not visible on the external surface, which is smooth. They slow the progress of the chyme and make it flow on a somewhat spiral path, which increases its contact with the mucosa and promotes more thorough mixing and nutrient absorption. Circular folds begin in the duodenum. In the jejunum, they are especially large, tall, and closely spaced. They become smaller and more sparse in the ileum. These changes are correlated with the relative amount of nutrient absorption occurring in each region. Circular folds are absent from the distal half of the ileum, but most nutrient absorption is completed by that point.
Villi (singular, villus) are tiny projections that give the inner lining of the intestine a fuzzy texture, like a terry cloth towel. They are about 0.5 to 1.0 mm high, with tongue- to finger like shapes. Villi are largest in the duodenum and become progressively smaller in more distal regions of the intestine. Villi are covered with two kinds of epithelial cells: columnar enterocytes (absorptive cells) and mucus-secreting goblet cells. Like epithelial cells of the stomach, those of the small intestine are joined by tight junctions that prevent digestive enzymes from seeping between them and eroding the underlying tissue.
The core of a villus is filled with areolar tissue of the lamina propria and contains an arteriole, blood capillaries, a venule, and a lymphatic capillary called a lacteal. The blood capillaries absorb most nutrients, but the lacteal absorbs most lipids. Lipids give its contents a milky appearance for which the lacteal is named. The core of the villus also has a few smooth muscle cells that contract periodically. This enhances mixing of the chyme in the intestinal lumen and milks lymph down the lacteal to larger lymphatics in the submucosa.
Microvilli are much smaller plasma membrane extensions, about 1 μm high, that form a fuzzy brush border on the surface of each enterocyte. In addition to increasing surface area, they contain brush border enzymes in the plasma membrane. These enzymes carry out some of the final stages of chemical digestion. They are not secreted into the lumen; instead, the chyme must contact the brush border for digestion to occur. This process, called contact digestion, is one reason why it is so important that intestinal contractions churn the chyme and ensure that it all contacts the mucosa.
On the floor of the small intestine, between the bases of the villi, there are numerous pores that open into tubular glands called intestinal crypts. These crypts, similar to the gastric glands, extend as far as the muscularis mucosae. In the upper half, they consist of enterocytes and goblet cells like those of the villi. The lower half is dominated by dividing stem cells. In its life span of 3 to 6 days, an epithelial cell migrates up the crypt to the tip of the villus, where it is sloughed off and digested. A few Paneth cells are clustered at the base of each crypt. They secrete lysozyme, phospholipase, and defensins—defensive proteins that resist bacterial invasion of the mucosa.
The duodenum has prominent duodenal glands in the submucosa. They secrete an abundance of bicarbonate-rich mucus, which neutralizes stomach acid and shields the mucosa from its erosive effects. Throughout the small intestine, the lamina propria and submucosa have a large population of lymphocytes that intercept pathogens before they can invade the bloodstream. In some places, these are aggregated into conspicuous lymphatic nodules such as the Peyer patches of the ileum.
Digestion and Absorption of the Major Organic Nutrients in the Small Intestine
Chemical digestion and nutrient absorption are essentially finished by the time food residue leaves the small intestine and enters the cecum.
Absorption of Water
The digestive system is one of several systems involved in fluid balance. The digestive tract receives about 9 L of water per day—0.7 L in food, 1.6 L in drink, and 6.7 L in the gastrointestinal secretions: saliva, gastric juice, bile, pancreatic juice, and intestinal juice. About 8 L of this is absorbed by the small intestine and 0.8 L by the large intestine, leaving 0.2 L voided in the daily fecal output.
Water is absorbed by osmosis, following the absorption of salts and organic nutrients that create an osmotic gradient from the intestinal lumen to the extracellular fluid (ECF). Diarrhea occurs when the large intestine absorbs too little water. This occurs when the intestine is irritated by bacteria and feces pass through too quickly for adequate reabsorption, or when the feces contain abnormally high concentrations of a solute such as lactose that opposes osmotic absorption of water. Constipation occurs when fecal movement is slow, too much water is reabsorbed, and the feces become hardened. This can result from lack of dietary fiber, lack of exercise, emotional upset, or long-term laxative abuse.
Absorption of Carbohydrates
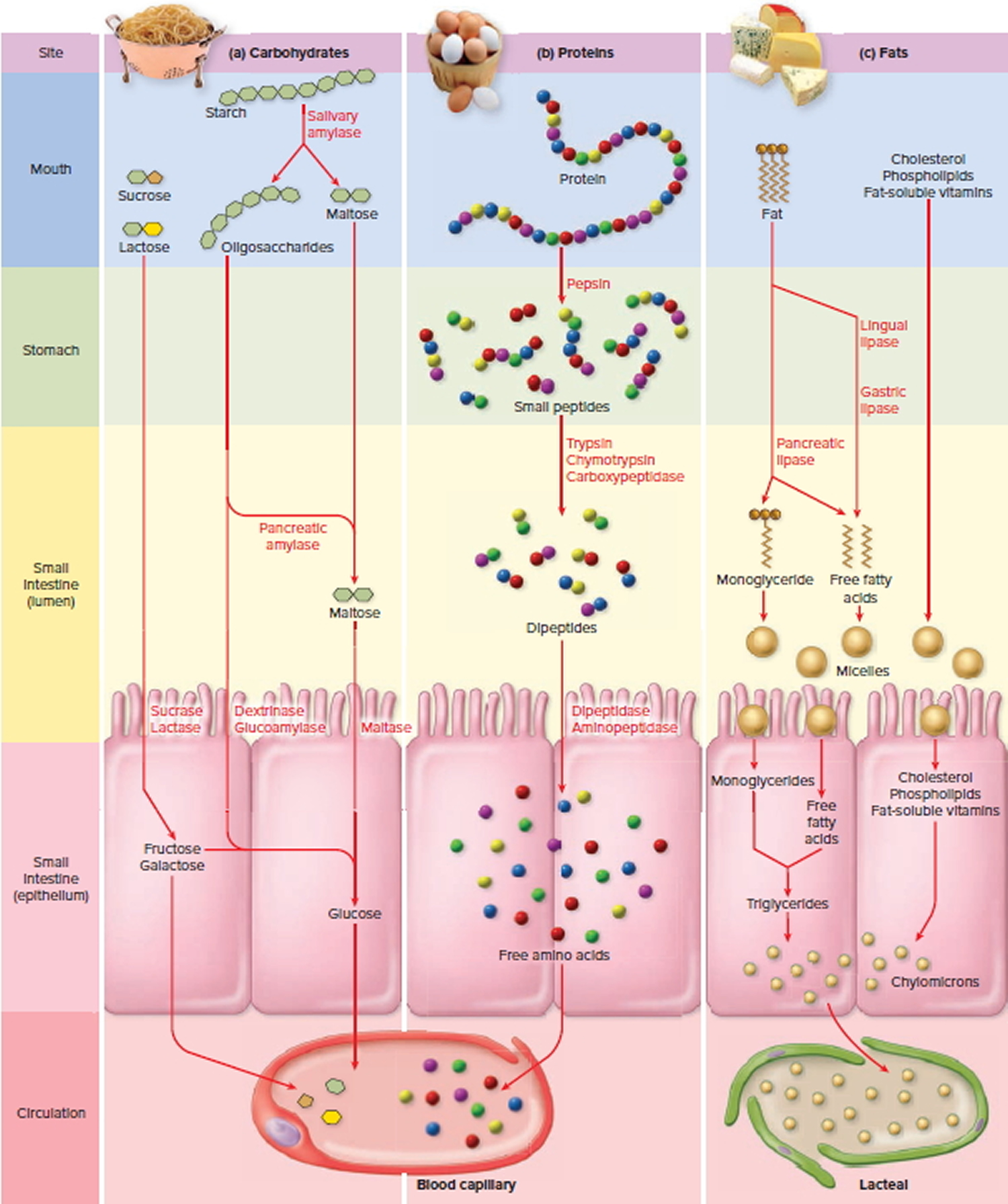
Most digestible dietary carbohydrate is starch. Starch is digested first to oligosaccharides up to eight glucose residues long, then into the disaccharide maltose, and finally to glucose, which is absorbed by the small intestine.
The process begins in the mouth, where salivary amylase breaks starch down into shorter segments (oligosaccharides). Salivary amylase functions best at pH 6.8 to 7.0, typical of the oral cavity. It is quickly denatured upon contact with stomach acid, but it can digest starch for as long as 1 to 2 hours in the stomach as long as it is in the middle of a food mass and escapes contact with the acid. Amylase therefore works longer when the meal is larger, especially in the fundus, where gastric motility is weakest and a food bolus takes longer to break up. As acid, pepsin, and the churning contractions of the stomach break up the bolus, amylase is denatured; it cannot function at a pH any lower than 4.5. Being a protein, amylase is then digested by pepsin along with the dietary proteins.
About 50% of the dietary starch is digested before it reaches the small intestine. Its digestion resumes in the small intestine when the chyme mixes with pancreatic amylase. Starch is entirely converted to oligosaccharides and maltose within 10 minutes. Its digestion is completed as the chyme contacts the brush border of the enterocytes. Two brush border enzymes, dextrinase and glucoamylase, hydrolyze oligosaccharides that are three or more residues long, while maltase hydrolyzes maltose. The end product of all of these is glucose, which is then absorbed.
Maltose is also present in some foods, but the major dietary disaccharides are sucrose (cane sugar) and lactose (milk sugar). They are digested by the brush border enzymes sucrase and lactase, respectively, and the resulting monosaccharides are immediately absorbed (glucose and fructose from the former; glucose and galactose from the latter). In most of the world population, however, lactase production ceases or declines to a low level after age 4 and lactose becomes indigestible.
The plasma membrane of the enterocytes has transport proteins that absorb monosaccharides as fast as they are produced by the foregoing enzymes. About 80% of the absorbed sugar is glucose, which is taken up by a sodium–glucose transporter like that of the kidney tubules. The glucose is subsequently transported out the base of the cell into the extracellular fluid (ECF). Sugar entering the extracellular fluid (ECF) increases its osmolarity, and this draws water osmotically from the lumen of the intestine, through the now-leaky tight junctions between the epithelial cells. Water carries more glucose and other nutrients with it by solvent drag, much as it does in the kidney. After a high-carbohydrate meal, solvent drag absorbs two to three times as much glucose as the sodium–glucose transporter.
The sodium–glucose transporter also absorbs galactose, whereas fructose is absorbed by facilitated diffusion using a separate carrier that doesn’t depend on Na+. Inside the enterocyte, most fructose is converted to glucose. Glucose, galactose, and the small amount of remaining fructose are then transported out the base of the cell by facilitated diffusion and absorbed by the blood capillaries of the villus. The hepatic portal system delivers them to the liver.
Absorption of Vitamins
Vitamins are not digested, but absorbed unchanged. The fat-soluble vitamins A, D, E, and K are absorbed with other lipids. Therefore, if they are ingested without fat-containing food, such as by simply popping vitamin tablets with a glass of water, they’re not absorbed at all but passed in the feces and wasted. Water-soluble vitamins (the B complex and vitamin C) are absorbed by simple diffusion. An exception is vitamin B12, an unusually large molecule that is absorbed poorly unless bound to the intrinsic factor (IF) secreted by the stomach’s parietal cells. Then as it passes down the small intestine, the B12–IF complex binds to receptors on absorptive cells of the distal ileum, where it is taken up by receptor-mediated endocytosis.
Absorption of Proteins
The amino acids absorbed by the small intestine come from three sources: (1) dietary proteins, (2) digestive enzymes digested by each other, and (3) sloughed epithelial cells digested by these enzymes. Amino acids from the last two sources total about 30 g/day, compared with about 44 to 60 g/day from the diet.
Enzymes that digest proteins are called proteases (peptidases). They are absent from the saliva but first encountered in the stomach. Here, pepsin hydrolyzes any peptide bond between tyrosine and phenylalanine, thereby digesting 10% to 15% of the dietary protein into shorter polypeptides and a small amount of free amino acids. Pepsin has an optimal pH of 1.5 to 3.5, so it is inactivated when it passes into the duodenum and mixes with the alkaline pancreatic juice (pH 8).
In the small intestine, the pancreatic enzymes trypsin and chymotrypsin take over protein digestion by hydrolyzing polypeptides into even shorter oligopeptides. Finally, these are taken apart one amino acid at a time by three more enzymes: (1) Carboxypeptidase removes amino acids from the —COOH end of the chain; (2) aminopeptidase removes them from the —NH2 end; and (3) dipeptidase splits dipeptides in the middle and releases the last two free amino acids. The last two of these are brush border enzymes, whereas carboxypeptidase is a pancreatic secretion.
Amino acid absorption is similar to that of monosaccharides. Enterocytes have several sodium-dependent amino acid cotransporters for different classes of amino acids. Dipeptides and tripeptides can also be absorbed, but they are hydrolyzed within the enterocytes before their amino acids are released to the bloodstream. At the basal surfaces of the cells, amino acids behave like the monosaccharides discussed previously—they leave the cell by facilitated diffusion, enter the capillaries of the villus, and are carried away in the hepatic portal circulation.
The absorptive cells of infants can take up intact proteins by pinocytosis and release them to the blood by exocytosis. This allows IgA from breast milk to pass into an infant’s bloodstream and confer passive immunity from mother to infant. It has the disadvantage, however, that intact proteins entering the infant’s blood are detected as foreign antigens and sometimes trigger food allergies. As the intestine matures, its ability to pinocytose protein declines but never completely ceases.
Figure 3. Digestion and Absorption of the Major Organic Nutrients in the Small Intestine
Absorption of Lipids
The hydrophobic quality of lipids makes their digestion and absorption more complicated than that of carbohydrates and proteins. Fats are digested by enzymes called lipases. Lingual lipase, secreted by the intrinsic salivary glands of the tongue, digests a small amount of fat while food is still in the mouth, but becomes more active at the acidic pH of the stomach. Here it is joined by gastric lipase, which makes a much larger contribution to preduodenal fat digestion. About 10% to 15% of dietary fat is digested before the chyme passes on to the duodenum.
Being hydrophobic, ingested fat takes the form of large globules that, without further physical processing, could be attacked by these lipases only at their surface. This would result in rather slow, inefficient digestion. The stomach’s vigorous antral pumping, however, breaks the fat up into small droplets dispersed through the watery chyme—that is, it emulsifies the fat, exposing much more of its surface to enzymatic action. The resulting emulsification droplets are promptly passed on to the duodenum and coated by certain components of the bile—lecithin and bile acids. These agents have hydrophobic regions attracted to the surface of a fat droplet and hydrophilic regions attracted to the surrounding water. The agitation produced by intestinal segmentation breaks the fat up further into droplets as small as 1 μm, and the coating of lecithin and bile acids keeps it broken up, preventing the droplets from coalescing into larger globules.
There is enough pancreatic lipase in the small intestine after a meal to digest the average daily fat intake in as little as 1 or 2 minutes. When lipase acts on a triglyceride, it removes the first and third fatty acids from the glycerol backbone and usually leaves the middle one. The products of lipase action are therefore two free fatty acids (FFAs) and a monoglyceride. Being smaller than triglycerides, these are more soluble in the enterocyte plasma membrane and thus easier to absorb.
The absorption of lipids depends on minute droplets in the bile called micelles. Micelles, made in the liver, consist of 20 to 40 bile acid molecules aggregated with their hydrophilic side groups facing outward and their hydrophobic steroid rings facing inward. Bile phospholipids and cholesterol diffuse into the center of the micelle to form its core. The micelles pass down the bile duct into the duodenum, where they absorb fat-soluble vitamins, more cholesterol, and the fatty acids and monoglycerides produced by fat digestion. Because of their charged, hydrophilic surfaces, micelles remain suspended in water more easily than free lipids do. They travel to the surfaces of the enterocytes, where they release their lipid cargo. Some of the lipids simply diffuse through the plasma membrane into the enterocytes, but these cells also have specific carrier proteins that facilitate their uptake. The micelles are reused, picking up another cargo of lipids and ferrying them to the enterocytes. Without micelles, the small intestine absorbs only about 40% to 50% of the dietary fat and almost no cholesterol.
Within the enterocytes, fatty acids and monoglycerides are transported into the smooth endoplasmic reticulum and resynthesized into triglycerides. The Golgi complex combines these with a small amount of cholesterol and coats the complex with a film of phospholipid and protein, forming droplets 75 to 1,200 nm in diameter called chylomicrons. It packages chylomicrons into secretory vesicles that migrate to the basal surface of the cell and release their contents into the core of the villus. Although some free fatty acids enter the blood capillaries, chylomicrons are too large to penetrate the endothelium. They are taken up instead by the more porous lacteals into the lymph. This fatty, milk-white intestinal lymph, called chyle, flows through larger and larger lymphatic vessels of the mesenteries, eventually passing through the cisterna chyli to the thoracic duct, then entering the bloodstream at the left subclavian vein.
Absorption of Minerals
Minerals (electrolytes) are absorbed along the entire length of the small intestine. Sodium ions are cotransported with sugars and amino acids. Chloride ions are actively transported in the distal ileum by a pump that exchanges them for bicarbonate ions, reversing the chloride–bicarbonate exchange that occurs in the stomach. Potassium ions are absorbed by simple diffusion. The K+ concentration of chyme rises as water is absorbed, creating a gradient favorable to K+ absorption. In diarrhea, when water absorption is hindered, potassium ions remain in the intestine and pass with the feces; therefore, chronic diarrhea can lead to hypokalemia.
Most minerals are absorbed at fairly constant rates regardless of need, leaving it to the kidneys to excrete any excess. Iron is one exception; its absorption is hormonally regulated. Intestinal enterocytes bind ferrous ions (Fe2+) and take them in by active transport; they cannot absorb ferric ions (Fe3+), but stomach acid (HCl) reduces most Fe3+ to absorbable Fe2+. Fe2+ is transported to the basal surface of the cell and there taken up by the extracellular protein transferrin. The transferrin–iron complex diffuses into the blood and is carried to such places as the bone marrow for hemoglobin synthesis, muscular tissue for myoglobin synthesis, and the liver for storage.
Excess dietary iron, if absorbed, binds irreversibly to ferritin in the enterocyte and is held there until that cell sloughs off and passes in the feces. Iron absorption and mobilization are regulated by the liver hormone hepcidin. An iron overload is dangerously toxic (indeed, a leading cause of death in young children who get into a parent’s iron supplement pills), but hepcidin normally prevents overload. It inhibits intestinal iron absorption and the mobilization of iron from the liver, thus preventing the blood iron level from rising too high. Anemia and hypoxia reduce hepcidin synthesis, removing its inhibitory effect and thus allowing increased absorption of dietary iron and mobilization of stored iron so it becomes available for hemoglobin synthesis.
The small intestine absorbs nearly all dietary phosphate, predominantly by active transport. By contrast, it absorbs only about 40% of the dietary calcium, leaving the rest to pass in the feces. In the duodenum, calcium is absorbed by the transcellular route. It enters the enterocytes through calcium channels in the apical plasma membrane and binds to a cytoplasmic protein called calbindin. This keeps the intracellular concentration of free calcium low, maintaining a gradient that favors uptake. What free calcium exists in the cytoplasm is then pumped out the basal side of the cell by active transport, using a protein called calcium–ATPase as well as a sodium–calcium antiport. From there, it enters the blood capillaries of the villus.
Transcellular calcium uptake is under hormonal influence. Parathyroid hormone is secreted in response to a drop in blood calcium level. It stimulates the kidneys to synthesize vitamin D from the precursors made by the epidermis and liver. Vitamin D then affects the absorptive cells of the duodenum in three ways: It increases the number of calcium channels in the apical membrane, the amount of calbindin in the cytoplasm, and the number of calcium–ATPase pumps in the basal membrane. Thus, it increases absorption of dietary calcium and raises the level of calcium in the blood.
Because of their much greater length, the jejunum and ileum absorb much more calcium than the duodenum does, but here it is by the paracellular route (passing between cells) and is independent of hormones. Most absorbed calcium is from meat and dairy products. Although green leafy vegetables are high in calcium, little of this is absorbed because they also contain an agent, oxalate, that binds calcium and makes it unavailable for absorption. Dietary fat retards calcium absorption by reacting with it to form poorly absorbed calcium soaps.
Absorption of Nucleic Acids
The nucleic acids, DNA and RNA, are present in much smaller quantities than the polymers discussed previously. The nucleases (ribonuclease and deoxyribonuclease) of pancreatic juice hydrolyze these to their constituent nucleotides. Nucleosidases and phosphatases of the brush border then decompose the nucleotides into phosphate ions, nitrogenous bases, and simple sugars (ribose from RNA and deoxyribose from DNA). These products are transported across the intestinal epithelium by membrane carriers and enter the capillary blood of the villus.
Intestinal Secretion
The intestinal crypts secrete 1 to 2 L of intestinal juice per day, especially in response to acid, hypertonic chyme, and distension of the intestine. This fluid has a pH of 7.4 to 7.8. It contains water and mucus but relatively little enzyme. Most enzymes that function in the small intestine are found in the brush border and pancreatic juice.
Regulation of Small Intestinal Secretions
Goblet cells and intestinal glands secrete their products when chyme provides both mechanical and chemical stimulation. Distension of the intestinal wall activates the nerve plexuses within the wall and stimulates parasympathetic reflexes that also trigger release of small intestinal secretions.
Intestinal Motility
Contractions of the small intestine serve three functions: (1) to mix chyme with intestinal juice, bile, and pancreatic juice, allowing these fluids to neutralize acid and digest nutrients more effectively; (2) to churn chyme and bring it into contact with the mucosa for contact digestion and nutrient absorption; and (3) to move residue toward the large intestine.
Segmentation is a movement in which stationary ringlike constrictions appear at several places along the intestine and then relax as new constrictions form elsewhere. This is the most common type of intestinal contraction. Its effect is to knead or churn the contents. Pacemaker cells of the muscularis externa set the rhythm of segmentation, with contractions about 12 times per minute in the duodenum and 8 to 9 times per minute in the ileum. Since the contractions are less frequent distally, segmentation causes slow progression of the chyme toward the colon. The intensity (but not frequency) of contractions is modified by nervous and hormonal influences.
When most nutrients have been absorbed and little remains but undigested residue, segmentation declines and peristalsis begins. The duodenum secretes a hormone called motilin that triggers a peristaltic wave beginning in the duodenum. The wave travels 10 to 70 cm and dies out, only to be followed by another wave that begins a little farther down the tract than the first one. These successive, overlapping waves of contraction are called a migrating motor complex. They milk the chyme toward the colon over a period of about 2 hours. A second complex then expels residue and bacteria from the small intestine, thereby helping to limit bacterial colonization. Refilling of the stomach at the next meal suppresses peristalsis and reactivates segmentation as new chyme enters the small intestine.
The ileocecal valve is usually closed. Food in the stomach, however, triggers both the release of gastrin and the gastroileal reflex, both of which enhance segmentation in the ileum and relax the valve. As the cecum fills with residue, the pressure pinches the valve shut and prevents the reflux of cecal contents into the ileum.
Chemical digestion and nutrient absorption are essentially finished by the time food residue leaves the small intestine and enters the cecum. But before going on to the functions of the large intestine, we trace each major class of nutrients—especially carbohydrates, proteins, and fats—from the mouth through the small intestine to see how it is chemically degraded and absorbed.
Parts of the small intestine
The small intestine consists of three parts: the duodenum, the jejunum, and the ileum.
The small intestine is a coiled mass of hollow tube filling most of the abdominal cavity inferior to the stomach and liver. A double-layered fold of peritoneal membrane called mesentery suspends the jejunum and ileum from the posterior abdominal wall. The mesentery supports the blood vessels, nerves, and lymphatic vessels that supply the intestinal wall. A filmy, double fold of peritoneal membrane called the greater omentum drapes like an apron from the stomach over the transverse colon and the folds of the small intestine. If the wall of the alimentary canal becomes infected, cells from the omentum may adhere to the inflamed region, helping to wall off the area. This action prevents spread of the infection to the peritoneal cavity.
Duodenum
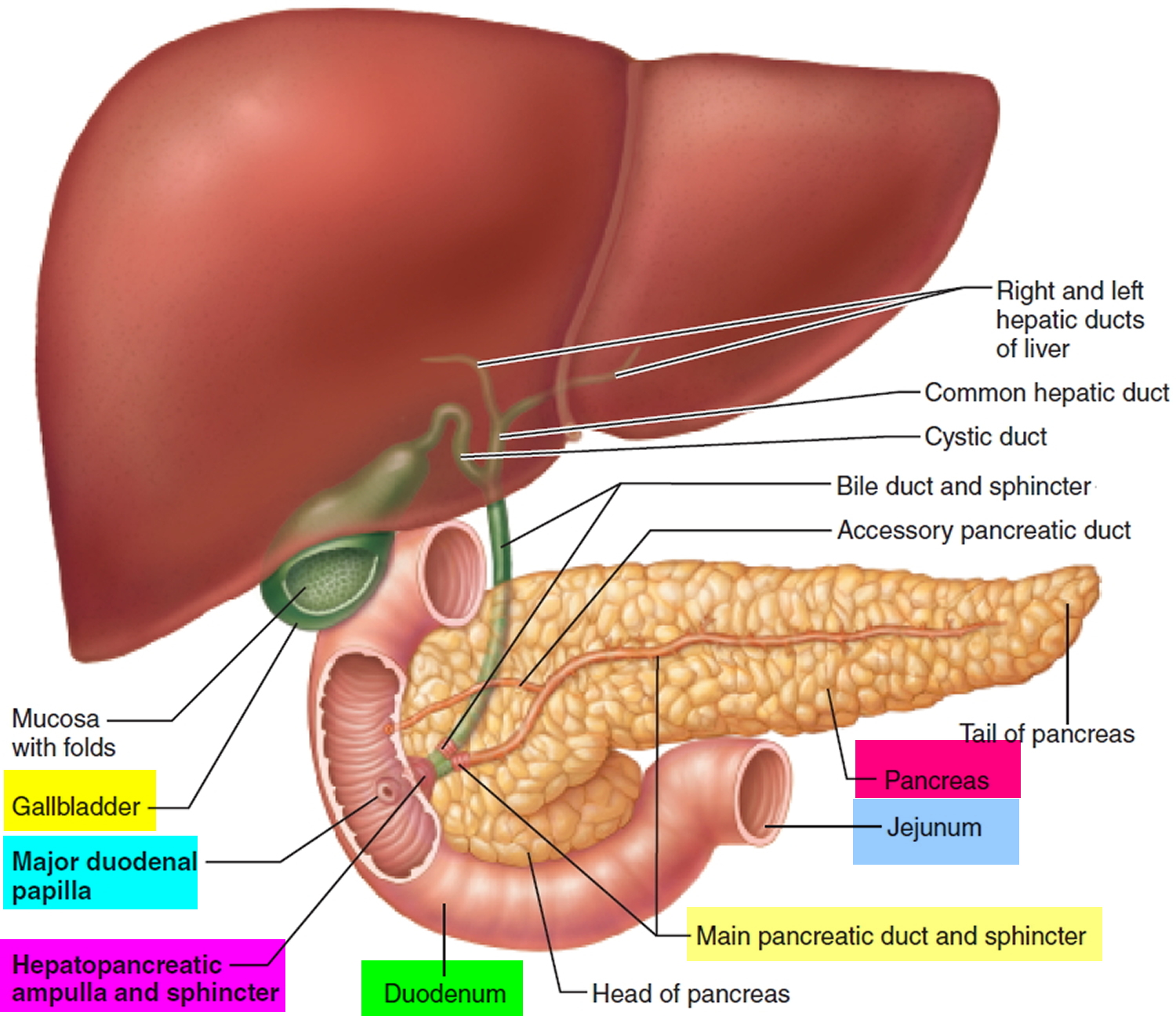
The first part of the small intestine is the duodenum. This C-shaped structure, adjacent to the head of the pancreas, is 20 to 25 cm long and 5 centimeters in diameter. The duodenum is a “mixing bowl” that receives chyme from the stomach and digestive secretions from the pancreas and liver. Almost all essential digestive enzymes enter the small intestine from the pancreas.
The duodenum begins at the pyloric valve, arcs around the head of the pancreas and passes to the left, and ends at a sharp bend called the duodenojejunal flexure. Slightly distal to the pyloric valve, it exhibits wrinkles called the major and minor duodenal papillae, where it receives the pancreatic duct and accessory pancreatic duct, respectively. Along with the pancreas, most of the duodenum is retroperitoneal. It receives the stomach contents, pancreatic juice, and bile. Stomach acid is neutralized here, fats are physically broken up (emulsified) by the bile acids, pepsin is inactivated by the elevated pH, and pancreatic enzymes take over the job of chemical digestion.
The duodenum is above the level of the umbilicus; its lumen is the widest of the small intestine. It is retroperitoneal except for its beginning, which is connected to the liver by the hepatoduodenal ligament, a part of the lesser omentum.
Figure 4. Duodenum
The duodenum is divided into four parts.
- The superior part (first part) extends from the pyloric orifice of the stomach to the neck of the gallbladder, is just to the right of the body of vertebra LI, and passes anteriorly to the bile duct, gastroduodenal artery, portal vein, and inferior vena cava. Clinically, the beginning of this part of the duodenum is referred to as the ampulla or duodenal cap, and most duodenal ulcers occur in this part of the duodenum.
- The descending part (second part) of the duodenum is just to the right of midline and extends from the neck of the gallbladder to the lower border of vertebra LIII. Its anterior surface is crossed by the transverse colon, posterior to it is the right kidney, and medial to it is the head of the pancreas. This part of the duodenum contains the major duodenal papilla, which is the common entrance for the bile and pancreatic ducts, and the minor duodenal papilla, which is the entrance for the accessory pancreatic duct, and the junction of the foregut and the midgut just below the major duodenal papilla.
- The inferior part (third part) of the duodenum is the longest section, crossing the inferior vena cava, the aorta, and the vertebral column. It is crossed anteriorly by the superior mesenteric artery and vein.
- The ascending part (fourth part) of the duodenum passes upward on, or to the left of, the aorta to approximately the upper border of vertebra LII and terminates at the duodenojejunal flexure. This duodenoj ejunal flexure is surrounded by a fold of peritoneum containing muscle fibers called the suspensory muscle (ligament) of duodenum (ligament of Treitz).
Jejunum
The jejunum, by definition, is the first 40% of the small intestine beyond the duodenum—about 1.0 to 1.7 m in a living person. The jejunum begins in the upper left quadrant of the abdomen but lies mostly within the umbilical region. Its wall is thick and muscular, and it has an especially rich blood supply, which gives it a relatively red color. Most digestion and nutrient absorption occur here.
The arterial supply to the jejunum includes jejunal arteries from the superior mesenteric artery.
Ileum
The ileum forms the last 60% of the post-duodenal small intestine (about 1.6 to 2.7 m). It occupies mainly the hypogastric region and part of the pelvic cavity. Compared with the jejunum, its wall is thinner, less muscular, less vascular, and has a paler pink color. On the side opposite from its mesenteric attachment, the ileum contains 20–30 masses of lymphoid tissue called submucous aggregated lymphoid nodules or Peyer’s patches, which are readily visible to the naked eye and become progressively larger approaching the large intestine. These lymphoid tissues are most abundant in the terminal portion of the ileum, near the entrance to the large intestine. The lymphocytes in the aggregated lymphoid nodules protect the small intestine from bacteria that normally inhabit the large intestine.
The end of the small intestine is the ileocecal junction, where the ileum joins the cecum of the large intestine. The muscularis of the ileum is thickened at this point to form a sphincter, the ileocecal valve, which protrudes into the cecum. The ileocecal valve surrounds the opening into the large intestine and it regulates the passage of food residue into the large intestine and prevents feces from backing up into the ileum.
The ileocecal valve is usually closed. Food in the stomach, however, triggers both the release of gastrin and the gastroileal reflex, both of which enhance segmentation in the ileum and relax the valve. As the cecum fills with residue, the pressure pinches the valve shut and prevents the reflux of cecal contents into the ileum.
Blood supply of the small intestine
The small intestine receives nearly all of its blood supply from the superior mesenteric artery, which fans out through the mesentery to give rise to 12 to 15 jejunal and ileal arteries leading to the intestinal wall. Branches of these arteries travel through the submucosa and give rise to capillary beds in the villi (see Figure 2), where the blood picks up all absorbed nutrients except lipids. Blood from here converges on another fanlike array of mesenteric veins, which leave by way of the superior mesenteric vein. This joins the splenic vein and then flows into the hepatic portal system, headed for the liver with its load of nutrients.


{kind=link}