
Contents
- What is capsaicin
- Pain management with topical capsaicin
- Capsaicin health benefits
- Capsaicin impact on gastric pathology
- Capsaicin stimulates the TRPV1 receptor
- Capsaicin can increase expression and activation of eNOS
- Capsaicin efficacy in metabolic syndrome
- Capsaicin boosts UCP2 expression in certain tissues
- Thermogenic and appetite control effects of capsaicin and capsiate
- Capsaicin dosage considerations
What is capsaicin
Capsaicin is a chili pepper extract (responsible for the spiciness of peppers) with analgesic properties. Capsaicin comes from plants of the genus Capsicum, which belong to the Solanaceae family. Capsaicin is the main pungent chemical, the others being dihydrocapsaicin, nordihydrocapsaicin, homodihydrocapsaicin and homocapsaicin 1. Capsaicin has long been available in various formulations as lotions, creams or patches in low concentrations of 0.025% to 0.075%. These products could be purchased over the counter to treat neuropathic and nociceptive musculoskeletal pain, such as postherpetic neuralgia, diabetic neuropathy, postsurgical pain, osteoarthritis and rheumatic arthritis. Chilli extracts have also been used to treat itching, psoriasis, cluster headache and migraines 2. A recent Cochrane review on low-concentration capsaicin in the treatment of neuropathic pain concluded that there were insufficient data to make any treatment recommendations for clinical practice 3. It also suggested that it had no effect beyond that of placebo 3. Due to the nature of interaction with the transient receptor potential vanilloid (TRPV1) receptor, a thermal nociceptor, capsaicin suffered two main disadvantages. Firstly, there were burning sensations and skin reactions that were not tolerated by many, and secondly, multiple daily applications for at least 4–6 weeks were required to see any response, leading to issues with compliance.
The main site of action of capsaicin is the transient receptor potential vanilloid 1 (TRPV1) channel. Although the channel is nonselectively activated by many endogenous and exogenous agonists, capsaicin itself is highly selective and potent at this channel.
Transient receptor potential vanilloid 1 (TRPV1) is a ligand-gated cation channel selectively expressed in the polymodal nociceptive nerve fibres, mainly the C and A delta fibres. It was the first pain transducer to be discovered. It is activated by heat (>43° C) and by acidosis (pH < 6). It is also activated by a number of endogenous agonists like anandamide, N-acetyldopamines, leukotriene B4, long-chain unsaturated fatty acids and, as recent evidence has shown, 9- and 13-hydroxyoctadecadienoic acid 4. When activated by a combination of heat, acidosis, or endogenous/exogenous agonists, TRPV1 may open transiently and initiate depolarization mediated by the influx of sodium and calcium ions. In the nociceptive sensory nerves which selectively express TRPV1 (mostly C- and some Aδ-fibres), depolarization results in action potentials, which propagate into the spinal cord and brain, and may be experienced as warming, burning, stinging, or itching sensations (Figure 1).
Figure 1. Activation of TRPV1 by capsaicin results in sensory neuronal depolarization
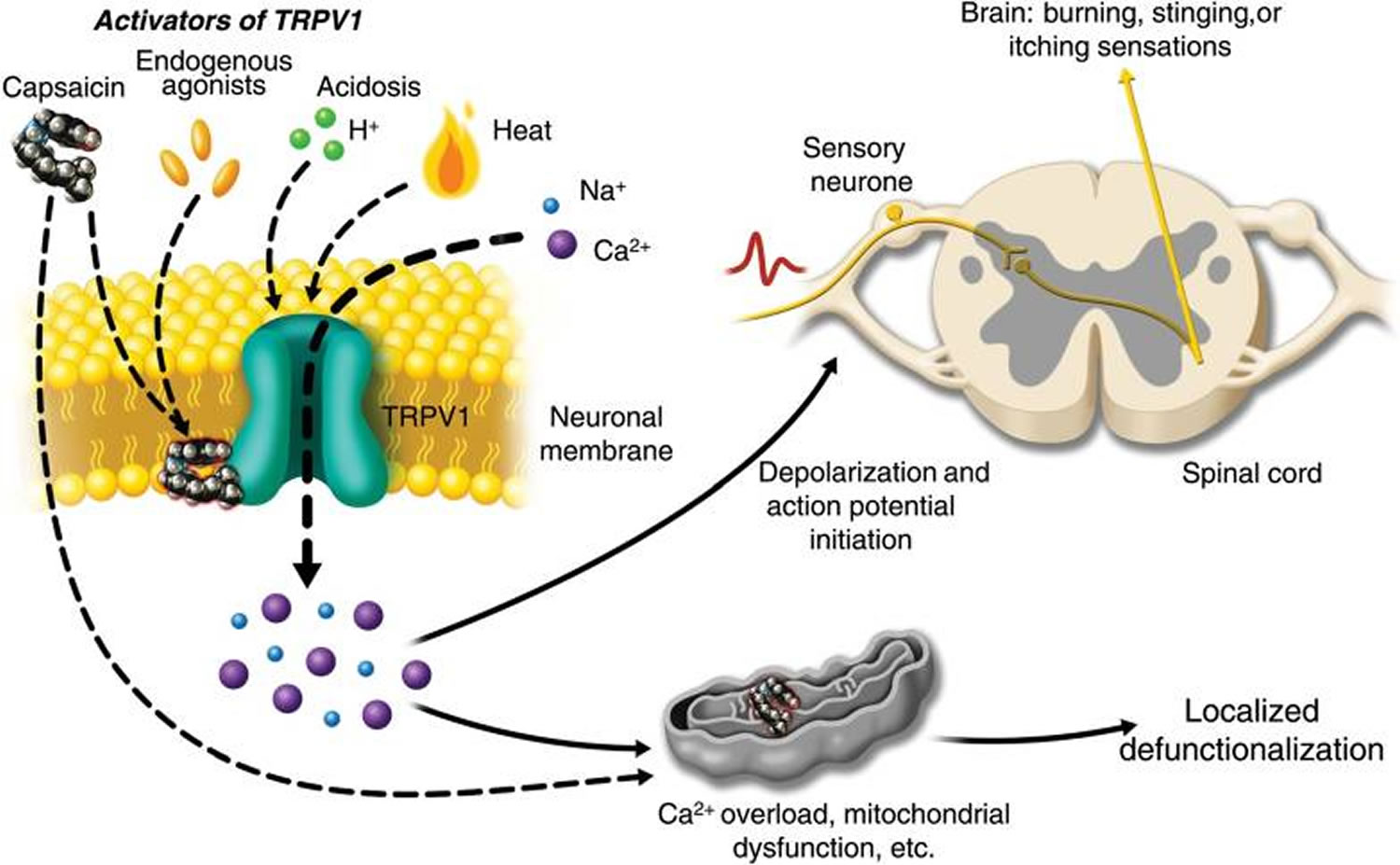
Note: Activation of TRPV1 by capsaicin results in sensory neuronal depolarization, and can induce local sensitization to activation by heat, acidosis, and endogenous agonists. Topical exposure to capsaicin leads to the sensations of heat, burning, stinging, or itching. High concentrations of capsaicin or repeated applications can produce a persistent local effect on cutaneous nociceptors, which is best described as defunctionalization and constituted by reduced spontaneous activity and a loss of responsiveness to a wide range of sensory stimuli.
[Source 5]In contrast to endogenous chemicals, capsaicin, being a stable compound, has a more persistent rather than transient effect on the channel. Moreover, the calcium:sodium permeability is increased from a baseline of 8:1 to 25:1. There is a massive influx of calcium ions down the electrochemical gradient. Furthermore, as TRPV1 is also expressed on intracellular organelles, external capsaicin application can cause release of calcium from the endoplasmic reticulum and induce additional intracellular calcium release from internal stores via calcium-dependent calcium release. The excess intracellular calcium triggers calcium-dependent protease enzymes causing cytoskeletal breakdown. Microtubule depolymerization causes a halt in fast axonal transport 6. Chloride influx accompanies the influx of positive ions causing osmotic swelling. A TRPV1-independent mechanism also exists by causing direct inhibition of electron-chain transport and subsequent mitochondrial dysfunction 7. Thus, multiple mechanisms ultimately lead to loss of cellular integrity and ‘defunctionalization’ of the nociceptor fibres. The nerve fibres retract to a depth at which mitochondrial function is preserved. Immunohistochemical studies have shown that capsaicin produces a highly localized loss of nerve fibres in the epidermis and dermis 8.
This replaces the hypothesis from the mid-1980s that substance P depletion is responsible for capsaicin-induced pain relief. A persistent confusion which continues to appear in the medical literature involves the role of ‘substance P depletion’ in capsaicin-induced pain relief. The neurogenic inflammation which follows application of topical capsaicin is due to the vascular actions of substance P and calcitonin gene-related peptide (CGRP) released from C-fibres. Mast cell degranulation is contributory but not necessary. There is no evidence that the neurogenic inflammation which accompanies topical capsaicin administration is related to prolonged pain relief, even though it has long been appreciated that systemic capsaicin can cause substance P release by nociceptors. It is now widely recognized that of all the neuropeptides released by C-fibres, calcitonin gene-related peptide (CGRP) is a more likely potential contributor to pain pathophysiology, particularly in migraine 9. If nociceptive nerve fibres retract from the epidermis and dermis then all markers they contain will be lost, and substance P is just one of many. The reduction of substance P content in skin after topical capsaicin administration is thus consequent to this process of nerve fibre defunctionalization and retraction. The ‘substance P depletion’ hypothesis was used to describe the mechanism of action of the low-concentration capsaicin formulations which became available in the 1980s, and unfortunately over the years, this hypothesis continues to be repeated even in recent review articles and textbooks.
Capsaicin is rapidly metabolized in the liver by cytochrome enzymes. It has a high mean apparent clearance of 54,598 L/h, and is therefore rapidly eliminated with a t1/2 of 1.64 h. Oral capsaicin has an elimination t1/2 of 24.9 min 10. In vitro studies suggest that capsaicin is metabolized very slowly in the skin 11. A clear clinical advantage of this is that capsaicin lasts for a long time in the skin, which is its site of action, and any systemically absorbed drug is rapidly cleared. Also, because such small quantities of drug reach the systemic circulation, there are no implications for dose adjustments in hepatic or renal failure.
It is the theoretical ability of capsaicin to desensitise nociceptors that is exploited for therapeutic pain relief 12.
Neuropathic pain occurs as a consequence of damage to the central nervous system (CNS) (e.g. cerebrovascular accident, multiple sclerosis or spinal cord injury) or peripheral nervous system (e.g. painful diabetic neuropathy, postherpetic neuralgia, surgery). Topical agents are most likely to be used for localised, peripheral neuropathies and to treat some peripheral painful states, such as rheumatoid arthritis, osteoarthritis, post-herpetic neuralgia, cluster headaches, post-mastectomy pain syndrome, pruritus, psoriasis and diabetic neuropathy 13. In addition, capsaicin may be useful in controlling chemotherapy- and radiotherapy-induced mucositis.
Pain management with topical capsaicin
The first formal report of the pain-reducing properties of topical capsaicin in the West appeared in 1850 as a recommendation to use an alcoholic hot pepper extract on burning or itching extremities 14. Creams, lotions, and patches containing capsaicin, generally in the range of 0.025–0.1% by weight, are now sold in many countries, often without the requirement of a prescription, for the management of neuropathic and musculoskeletal pain. Clinical studies of these medications, usually involving three to five topical skin applications per day for periods of 2–6 weeks, have generally suggested modest beneficial effects against various pain syndromes, including post-herpetic neuralgia, diabetic neuropathy, and chronic musculoskeletal pain 15, 16. Since low-concentration, capsaicin-based products often result in contamination of the patient’s environment (clothing, bedding, contact lenses, etc.) and each application may be associated with a burning sensation, poor patient compliance with these products is often cited as a likely contributor to limited efficacy 17.
Topical capsaicin formulations are widely used to manage pain. Low-concentration creams, lotions, and patches intended for daily skin application have been available in most countries since the early 1980s. Prescriptions are usually not needed for these self-administered medicines, which often have not been reviewed formally by drug regulatory authorities. The recent approval in the EU and USA of a prescription-strength high-concentration single-administration capsaicin 8% patch (Qutenza™), which provides rapid, long-lasting pain relief with a single application. Following positive trials in postherpetic neuralgia and HIV-associated distal sensory polyneuropathy (HIV-DSP), the European Union (EU) approved its use in peripheral neuropathy in nondiabetic patients, while the US Food and Drug Administration (FDA) approved its use in the USA only to treat postherpetic neuralgia.
The capsaicin 8% dermal patch is available as a 280 cm2 patch containing 179 mg of capsaicin (640 µg/cm2). It is different from conventional transdermal patches in that it works locally on the skin and there is little systemic absorption 18. Capsaicin is extremely lipophilic and therefore is easily absorbed into the epidermal and dermal layers with little affinity for the aqueous blood phase. The amount of drug absorbed is greater with the duration of exposure and also the area applied. The pharmacokinetic data derived from clinical trials in peripheral neuropathy suggest that quantifiable capsaicin concentrations were more often seen in postherpetic neuralgia patients than HIV-associated distal sensory polyneuropathy (HIV-DSP) patients, as the skin of the feet is thicker compared with the rest of the body. Among HIV-DSP patients, they were more often found in those treated for 90 min 11. Blood samples were tested from 173 patients in the trials at varying intervals after patch removal (0, 1, 3, 6 and 24 h). Only 34 (20%) patients had quantifiable levels of capsaicin, and among all the blood samples only 6% were above the lower limit (0.5 mg/ml) of quantifiable plasma capsaicin 19. The Cmax or mean maximum plasma concentrations after 60- and 90-min applications were 1.38 ng/ml and 2.96 ng/ml, respectively 19; the Tmax to reach these levels were 1.46 and 1.51 h 19. Interestingly, after oral ingestion of 26.6 mg of capsaicin, the Cmax was 2.47 ng/ml in a mean time of 47 min 20.
Qutenza has not been directly compared to any other medications for the treating postherpetic neuralgia. Two pivotal clinical trials compared Qutenza to a control patch (0.04% capsaicin) in postherpetic neuralgia. The primary endpoint of both trials was the reduction in numeric pain rating scale score. Qutenza reduced pain from baseline to weeks 2 to 8 (29.6% and 32% reductions) compared to control (19.9% and 24.4% reductions). The improvement in numeric pain rating scale score scores persisted, with score reductions greater with Qutenza (29.9% and 32.3% reductions) compared to control (20.4% and 25% reductions) for the period 2 to 12 weeks. Safety and efficacy of capsaicin 8% has been demonstrated in open-label trials for up to 48 weeks. The most common adverse drug reactions occurring with capsaicin 8% are application site erythema (63%) and application site pain (42%). Some patients experienced transient increases in blood pressure during Qutenza application. Qutenza must be administered by a physician or under the close supervision of a physician. Prior to application, pretreat the affected area with a topical anesthetic to reduce application site pain. Some patients may require systemic analgesics during and after treatment for treatment-associated pain. Applications of Qutenza can be repeated no sooner than once every 3 months, as needed.
Capsaicin health benefits
Capsaicin impact on gastric pathology
Ironically, many people are under the impression that spicy foods can cause ulcers; to the contrary, there is evidence that capsaicin tends to prevent and accelerate healing of gastric ulcers 21, 22. This phenomenon reflects capsaicin’s ability to inhibit gastric acid secretion, boost secretion of alkali and mucous, and stimulate gastric blood flow. A clinical study found that the gastric tissue damage and microbleeding induced acutely by indomethacin or ethanol ingestion was blunted if capsaicin was administered concurrently 21. These findings have prompted the suggestion that capsaicin could be used as a protective adjuvant to non-steroidal anti-inflammatory drug therapy 21, 22. Limited epidemiology suggests that gastric ulcers may be less common in ethnic groups that prefer spicy foods 23.
With respect to risk of gastric cancer, the epidemiology on spicy foods is rather perplexing. A recent meta-analysis of pertinent studies in Korea and Mexico, where heavy consumption of spicy foods is common, concludes that moderate daily intakes of capsaicin (less than 30 mg daily) are associated with a significant decrease in gastric cancer risk relative to non-consumption—perhaps reflecting the gastroprotective effects of capsaicin—whereas, heavy daily consumption is associated with a notable increase in risk 24. Bley et al 25 suggest that the increase in risk associated with heavy consumption of spicy traditional foods might reflect mutagens present in these foods, rather than an effect of capsaicin per se. Aflatoxins, pesticides and nitrosamines or their precursors have been detected in chillies sold for human consumption 25. The traditional Korean dish kimchi, a salty pickled cabbage usually fermented with red pepper and linked to increased risk of gastric cancer, is typically high in nitrate and contains N-nitroso compounds with mutagenic potential; the high salt content of this food may act as gastric co-carcinogen 26. Studies with high-purity capsaicin indicate that it is not genotoxic; in animal studies, capsaicin lacks carcinogenicity, and opposes the carcinogenicity of certain mutagens 25. Further clarification of this situation will be desirable if in the future people are encouraged to consume more capsaicin for potential health benefits.
Capsaicin stimulates the TRPV1 receptor
Capsaicin has recently been shown to modulate metabolism via activation of transient receptor potential vanilloid 1 (TRPV1) receptors, which are found not only on nociceptive (pain) sensory neurons, but also in a range of other tissues. TRPV1 activation induces calcium influx, and in certain tissues this is associated with increased activation or expression of key proteins such as endothelial nitric oxide synthase (eNOS), uncoupling protein 2 (UCP2), KLF2, PPARdelta, PPARgamma, and LXRα.
Transient receptor potential vanilloid 1 (TRPV1) is a membrane receptor that, when activated, acts as a non-specific cation channel, allowing influx of calcium. Endogenous activators of TRPV1 include heat, low pH, and certain lipid metabolites; the best known exogenous activator is the phytochemical capsaicin 27. In as much as nanomolar concentrations of capsaicin can activate this receptor and capsaicin is efficiently absorbed, a sufficiently high oral intake of capsaicin can induce systemic activation of TRPV1 28.
TRPV1 is expressed by many nociceptive sensory neurons, and its activation triggers pain sensations. However, the massive neuronal calcium influx triggered by topical exposure to sufficient concentrations of capsaicin is potentially cytotoxic, and triggers a reflex down-regulation of TRPV1 activity 29. Hence, these neurons become less responsive to endogenous agonists for TRPV1, resulting in analgesia 30. Capsaicin skin patches are currently employed clinically for local pain control 31.
TRPV1 is also expressed by vascular endothelial cells, hepatocytes, adipocytes, smooth muscle cells, fibroblasts, various epithelia, T cells, mast cells, and by neurons and astrocytes in the brain and spinal column 32. Hence, TRPV1 has the potential to modulate the function of these cells by boosting their intracellular-free calcium levels (Cai). At present, there does not appear to be any evidence that the desensitisation phenomenon evoked by capsaicin in sensory neurons is pertinent to these other tissues; no down-regulation of TRPV1 expression or function was noted in the vasculature of newborn rats that had been injected with potent doses of capsaicin for 5 days 33.
Capsaicin can increase expression and activation of eNOS
The impact of TRPV1 activation on vascular endothelium is of particular interest, since an increase in Cai is a key mediator of the protective impact of pulsatile shear stress—and of aerobic exercise—on endothelial function. This increase in Cai acts rapidly to stimulate endothelial nitric oxide synthase (eNOS) activity via binding of the Ca2+/calmodulin complex; in addition, Cai-mediated activation of AMPK and Sirt1 stimulates eNOS activity by modifying its phosphorylation and acetylation status 34. In the longer term, expression of eNOS increases as well. Increased Cai acts to boost the expression and activity of the endothelium-specific transcription factor KLF2 via a complex chain of events involving activation of Ca2+/calmodulin-dependent kinase kinase-β and downstream phosphorylations of AMPK, ERK2, HDAC5, and the transcription factor MEF2 35. KLF2, in turn, promotes transcription of the eNOS, thrombomodulin, and Nrf2-responsive antioxidant enzymes, and works indirectly to suppress transcription of various proinflammatory proteins 36.
Capsaicin feeding has shown an antihypertensive effect in rats genetically prone to this disorder, and this compound also blunts the nocturnal rise in blood pressure or development of hypertension in mice fed a high salt diet 37. Conceivably, improved NO function may underlie these effects. Capsaicin dilates the coronary arteries of pigs ex vivo, an effect that is half-maximal at 116 nM; this effect is blocked by endothelial denudation and inhibitors of eNOS, and is less notable with coronaries from pigs experiencing metabolic syndrome, which disrupts eNOS function via oxidative stress.34 Release of CGRP from perivascular sensory neurons may also contribute to the vasodilatory impact of capsaicin 38. Paradoxically, the direct impact of capsaicin on vascular smooth muscle is to provoke constriction, owing to increased calcium influx 39. Hence, the net impact of capsaicin on vascular tone and blood pressure may reflect complex interactions and countervailing effects.
The antihypertensive effect of dietary capsaicin in salt-fed rats may reflect, in part, an inhibitory effect on renal sodium retention. In the kidney, cortical collecting duct epithelium expresses TPRV1, and its activation decreases the function and expression of epithelial sodium channels in these cells, resulting in increased urinary sodium loss 37. However, whether and how moderate, clinically tolerable dosing with capsaicin would influence human hypertension has not yet been assessed.
Capsaicin efficacy in metabolic syndrome
In obese mice, capsaicin injections exert an anti-inflammatory effect on adipose tissue, suppressing production of IL-6, TNF-α, MCP-1, and cox-2, while boosting that of adiponectin, and decreasing macrophage infiltration 40.
The beneficial effects of capsaicin on metabolic syndrome in mice may be mediated in part by increased secretion of glucagon-like peptide-1 (GLP-1). Indeed, gastric administration of capsaicin has been shown to evoke increased secretion of GLP-1 by the gastrointestinal (GI) tract, and to raise plasma levels of this factor 41.
Another intriguing TRPV1-dependent effect of capsaicin ingestion is activation of brown adipose tissue. Activation of TRPV1-expressing neurons in the digestive tract sends a signal to the brain via the vagal nerve; this in turn evokes an activation of sympathetic neurons that is selective for brown fat—that is, the heart rate is not impacted 42. Many clinical trials have evaluated the impact of capsaicin ingestion on metabolic rate, respiratory quotient and appetite; these conclude that capsaicin can modestly enhance energy expenditure, while boosting fat oxidation (lower RQ) and diminishing appetite—effects conducive to weight control 43.
Capsaicin boosts UCP2 expression in certain tissues
TRPV1 activation has also been shown to increase expression of uncoupling protein 2 (UCP2) in endothelial cells, hepatocytes and cardiac tissue 44. In the heart, this effect may be downstream from increased expression of PPARdelta, a factor which opposes cardiac hypertrophy and fibrosis 45. Hence, dietary capsaicin was found to oppose the cardiac hypertrophy induced by a high salt diet in mice—an effect not seen in TRPV1 knockout mice 46. With respect to UCP2, this functions as a mitochondrial uncoupling protein when mitochondrial substrate oxidation is high and superoxide generation is elevated; by diminishing the proton gradient across the mitochondrial inner membrane, UCP2 relieves the resistance to electron flow down the respiratory chain and hence decreases the rate at which electrons are shunted to superoxide generation at complexes I and III 47.
UCP2 can be of particular value when cells that are constitutively permeable to glucose—such as vascular endothelium—are subjected to hyperglycaemia. Under these circumstances, elevated glucose oxidation in the Krebs cycle tends to boost mitochondrial superoxide generation, an effect opposed by UCP2 48. In diabetic mice, capsaicin administration was shown to alleviate vascular oxidative stress and improve endothelium-dependent vasodilation—a phenomenon not seen in UCP2 knockout mice rendered diabetic 49. In men with diabetes, a polymorphism in the UCP2 promoter (-866G>A), linked to increased expression of UCP2 in some studies, was found to be associated with significantly lower risk for developing coronary disease49—consistent with a protective impact of UCP2 expression on cardiovascular risk in diabetics.
Hepatocyte expression of UCP2 can be protective in the context of non-alcoholic fatty liver disease. Under these circumstances, increased mitochondrial oxidation of fatty acids contributes to the oxidative stress that plays a mediating role in this syndrome. The uncoupling activity of UCP2 decreases this generation of superoxide, and by boosting the rate at which mitochondria can metabolise fatty acids, helps to mitigate the surplus of fatty acids within hepatocytes 50. Indeed, capsaicin-rich diets have been found to alleviate non-alcoholic fatty liver disease in mouse models of this disorder 51. TRPV1-mediated induction of PPARdelta likely plays a role in this effect, and may be upstream from UCP2 induction 51. Capsaicin-mediated induction of UCP2 in hepatocytes may have potential as an adjuvant to weight control strategies which attempt to optimise hunger control, selective fat oxidation, and thermogenesis by improving the efficiency of hepatic fatty acid oxidation 52.
Thermogenic and appetite control effects of capsaicin and capsiate
Another intriguing TRPV1-dependent effect of capsaicin ingestion is activation of brown adipose tissue. Activation of TRPV1-expressing neurons in the digestive tract sends a signal to the brain via the vagal nerve; this in turn evokes an activation of sympathetic neurons that is selective for brown fat—that is, the heart rate is not impacted 53. Many clinical trials have evaluated the impact of capsaicin ingestion on metabolic rate, respiratory quotient and appetite; these conclude that capsaicin can modestly enhance energy expenditure, while boosting fat oxidation (lower RQ) and diminishing appetite—effects conducive to weight control 54.
Similar effects are seen with a non-spicy analogue of capsaicin, capsiate, which owing to lower stability does not induce pain in the oral cavity and appears to have limited systemic availability 55. Capsiate is found in certain sweet peppers; it is very similar in structure to capsaicin, and can activate TRPV1, with an affinity about one-third that of capsaicin 56. Whereas capsaicin contains an amide linkage that is relatively stable, capsiate contains an ester that is readily cleaved; when administered orally, intact capsiate fails to reach oral TPRV1-expressing neurons, but does manage to stimulate such neurons lower in the gastrointestinal (GI) tract. No intact capsiate appears in the portal blood after oral administration, but its hydrolysis products are detectible, implying that capsiate is hydrolysed during the process of absorption 55. Hence, the effects of capsiate attributable to TPRV1 agonism appear to be mediated by stimulation of GI sensory neurons.
Both capsaicin and capsiate may have modest utility as adjuvants to weight control programmes. Supplementation with capsiate (9 mg daily) for 12 weeks in a double-blind study was shown to decrease abdominal fat mass relative to placebo, albeit to a modest extent 57. Over the 12 weeks, the capsiate group, on average, lost 0.4 kg of weight and 1 cm of waist girth beyond that achieved with placebo—not an effect of much practical importance unless it persists and increases over time. Not surprisingly, the effects of capsaicin or capsiate on thermogenesis are most notable in humans bearing detectible amounts of brown fat 58; however, there is some evidence that prolonged ingestion of these agents may lead to recruitment of brown fat in humans 59. These effects on thermogenesis are modest in magnitude; there do not appear to be any reports of clinically significant hyperthermia with ingestion of capsaicin or capsiate.
Some studies have also evaluated the impact of oral capsaicin or capsiate on appetite and subsequent food consumption in various contexts. The findings of these studies have been inconsistent, though an overview of these studies by Ludy et al 60 concludes that, on balance, consumption of these agents tends to decrease orexigenic sensations. In positive studies, capsaicin-treated subjects reported less desire to consume fatty foods, sweet foods, salty foods and food overall, and achieved greater satiety after meals. Also, calorie consumption during subsequent meals was sometimes reported to drop after capsaicin consumption. The fact that capsiate blunted appetite in some studies suggests that these effects are mediated by TRPV1-expressing GI neurons. Arguably, capsaicin in the GI tract triggers a vagal signal to appetite-regulatory centres in the brain; however, increased secretion of GLP-1 may also play a role in capsaicin’s impact on appetite.
Capsaicin dosage considerations
In rodents, large metabolic effects have been reported with dietary capsaicin intakes in the range of 0.01–0.02% of diet. If a human were to eat (say) 400 g dry weight of food daily, 0.01% of diet would correspond to 40 mg capsaicin. Oral administration of capsaicin represents a clinical challenge—many people, especially those not acclimated to a spicy diet, do not enjoy the oral pain associated with capsaicin-laced foods, and capsaicin capsules may cause GI distress in some persons; this latter effect is mitigated somewhat by ingesting capsaicin capsules with meals. When Lejeune et al 61 had study volunteers take 45 mg capsaicin three times daily with meals, 24% of them experienced significant stomach discomfort and were allowed to cut this dose in half; however, this dose regimen seems likely to be a higher dose than would be required for metabolic benefits.
Hot peppers typically contain capsaicin in conjunction with lesser amounts of its analogues, dihydrocapsaicin and nordihydrocapsaicin; the latter is a very minor component, but dihyrocapsaicin may constitute as much as 40% of total capsaicinoids. The relative proportion of capsaicin and dihydrocapsaicin in a food is of little practical import, as the abilities of these compounds to activate TRPV1 are roughly equivalent. Commercial capsules of cayenne pepper are available that provide 40 000–100 000 Scoville heat units per capsule. The Scoville scale quantifies the spicy heat (or pungency) of foods which contain capsaicinoids; a gram of capsaicin corresponds to 16 million Scoville heat units; a gram of dihydrocapsaicin to 15 million units; and a gram of nordihydrocapsaicin to 9.1 million units. Therefore, a capsule claiming 100 000 Scoville heat units can be expected to contain about 6.6 mg of capsaicinoids. Consuming three of these daily with meals will provide about 20 mg, and those who enjoy spicy foods could supplement this with peppers, pepper sauces, or cayenne powder added to foods. Perhaps this would be an appropriate ‘baseline’ regimen to study clinically. Topical administration of capsaicin in patches may represent a reasonable alternative in people unable to tolerate it orally—albeit this will be a more expensive option, and local pain is commonly experienced for an hour or more after patch application 62.
Summary of benefits
This brief overview should make it clear that dietary capsaicin—and, likely to a more limited degree, non-pungent capsiate—has intriguing potential for health promotion. Rodent studies suggest that capsaicin may merit clinical evaluation with respect to endothelial function, progression of atherosclerosis (most notably in diabetics), angina, non-alcoholic fatty liver disease, cardiac hypertrophy, metabolic syndrome, hypertension, obesity and gastric ulceration. (See Table 1 for a summary of these potential benefits and the mechanisms that may underlie them.) In addition to the many studies assessing capsaicin’s impact on metabolic rate and adiposity, the trial of topical capsaicin in patients with angina, and the studies documenting capsaicin’s gastroprotective effects, represent initial efforts in this regard. A study examining endothelium-dependent vasodilation in diabetics might be particularly useful, as a systemically adequate dose of capsaicin could be expected to have a notably favourable impact on this parameter. Assessment of the dose-dependency of this effect could provide useful insight into capsaicin clinical dosage schedules which could provide systemic metabolic benefits. Both oral and topical application of capsaicin could be tested in this regard. The rodent literature is sufficiently intriguing that serious efforts to evaluate the feasibility of capsaicin administration as a clinical or lifestyle strategy appear to be warranted. However, owing to the fact that TRPV1 receptors are expressed on a wide range of tissues, the possibility that high-dose capsaicin might exert unanticipated or unwanted physiological effects should be borne in mind.
Table 1. Health benefits of capsaicin administration suggested by preclinical and clinical research
| Condition benefited | Likely mechanisms of action |
|---|---|
| Atherosclerosis63 | Improved endothelial function, including eNOS activation/induction; induction of LXRalpha in foam cells, promoting cholesterol export |
| Diabetic vasculopathy49 | Induction of UCP2 and eNOS in endothelium |
| Stroke64 | Improved endothelial function, including eNOS activation/induction |
| Angina65 | Improved endothelium-dependent vasodilation of coronary arteries |
| Hypertension66 | Activation/induction of eNOS; decreased renal sodium retention |
| Metabolic syndrome67 | Decreased adipose inflammation—reflecting PPARgamma induction |
| Cardiac hypertrophy46 | Induction of PPARdelta |
| Fatty liver51 | Induction of UCP2 in hepatocytes; decreased adipose inflammation Increased GLP-1 secretion |
| Obesity68 | Sympathetic activation of brown fat thermogenesis Improved appetite control—vagal signal to appetite centers, ↑ GLP-1; increased adipocyte capacity for lipolysis |
| Gastric ulceration22 | Decreased acid secretion; increased alkali; increased gastric blood flow |
eNOS = endothelial nitric oxide synthase; GLP-1 = glucagon-like peptide-1; UCP2 = uncoupling protein 2
- Recent advances in the study on capsaicinoids and capsinoids. Luo XJ, Peng J, Li YJ. Eur J Pharmacol. 2011 Jan 10; 650(1):1-7. https://www.ncbi.nlm.nih.gov/pubmed/20946891/[↩]
- Martindale W. (1999) Reynolds J., editor. (ed.), The Extra Pharmacopoeia/Martindale, 32nd edition. London: The Pharmaceutical Press.[↩]
- Topical capsaicin (low concentration) for chronic neuropathic pain in adults. Derry S, Moore RA. Cochrane Database Syst Rev. 2012 Sep 12; (9):CD010111. http://onlinelibrary.wiley.com/doi/10.1002/14651858.CD010111/full[↩][↩]
- The paradoxical role of the transient receptor potential vanilloid 1 receptor in inflammation. Alawi K, Keeble J. Pharmacol Ther. 2010 Feb; 125(2):181-95. https://www.ncbi.nlm.nih.gov/pubmed/19896501/[↩]
- Anand P, Bley K. Topical capsaicin for pain management: therapeutic potential and mechanisms of action of the new high-concentration capsaicin 8% patch. BJA: British Journal of Anaesthesia. 2011;107(4):490-502. doi:10.1093/bja/aer260. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3169333/[↩]
- Capsaicin causes protein synthesis inhibition and microtubule disassembly through TRPV1 activities both on the plasma membrane and intracellular membranes. Han P, McDonald HA, Bianchi BR, Kouhen RE, Vos MH, Jarvis MF, Faltynek CR, Moreland RB. Biochem Pharmacol. 2007 May 15; 73(10):1635-45. https://www.ncbi.nlm.nih.gov/pubmed/17274957/[↩]
- Capsaicin and its analogs inhibit the activity of NADH-coenzyme Q oxidoreductase of the mitochondrial respiratory chain. Shimomura Y, Kawada T, Suzuki M. Arch Biochem Biophys. 1989 May 1; 270(2):573-7. https://www.ncbi.nlm.nih.gov/pubmed/2495767/[↩]
- The time course of epidermal nerve fibre regeneration: studies in normal controls and in people with diabetes, with and without neuropathy. Polydefkis M, Hauer P, Sheth S, Sirdofsky M, Griffin JW, McArthur JC. Brain. 2004 Jul; 127(Pt 7):1606-15. https://www.ncbi.nlm.nih.gov/pubmed/15128618/[↩]
- Calcitonin gene-related peptide receptor antagonists for migraine. Fischer MJ. Expert Opin Investig Drugs. 2010 Jul; 19(7):815-23. https://www.ncbi.nlm.nih.gov/pubmed/20482328/[↩]
- Pharmacology and treatment of neuropathic pains. Jensen TS, Madsen CS, Finnerup NB. Curr Opin Neurol. 2009 Oct; 22(5):467-74. https://www.ncbi.nlm.nih.gov/pubmed/19741531/[↩]
- European Medicines Agency (2013) Summary of Product Characteristics: Qutenza 179 mg cutaneous patch. http://www.ema.europa.eu/docs/en_GB/document_library/EPAR_-_Product_Information/human/000909/WC500040453.pdf[↩][↩]
- The pharmacological challenge to tame the transient receptor potential vanilloid-1 (TRPV1) nocisensor. Holzer P. Br J Pharmacol. 2008 Dec; 155(8):1145-62. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2607216/[↩]
- Reynolds 1999. Martindale: the extra pharmacopoeia. 2nd Edition Reynolds JEF, editor. Royal Pharmaceutical Society; London: 1999.[↩]
- Turnbull A. Tincture of capsaicin as a remedy for chilblains and toothache. Dublin Free Press. 1850;1:95–6.[↩]
- Derry S, Lloyd R, Moore RA, McQuay HJ. Topical capsaicin for chronic neuropathic pain in adults. The Cochrane database of systematic reviews. 2009;(4):CD007393. doi:10.1002/14651858.CD007393.pub2. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4161117/[↩]
- Hempenstall K, Nurmikko TJ, Johnson RW, A’Hern RP, Rice AS. Analgesic Therapy in Postherpetic Neuralgia: A Quantitative Systematic Review. Woolf CJ, ed. PLoS Medicine. 2005;2(7):e164. doi:10.1371/journal.pmed.0020164. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1181872/[↩]
- Altman R, Barkin RL. Topical therapy for osteoarthritis: clinical and pharmacologic perspectives. Postgrad Med. 2009;121:139–47. https://www.ncbi.nlm.nih.gov/pubmed/19332972[↩]
- Capsaicin 8% Topical Patch (Qutenza)—A Review of the Evidence. Journal of Pain & Palliative Care Pharmacotherapy Vol. 25 , Iss. 1,2011. http://www.tandfonline.com/doi/full/10.3109/15360288.2010.547561[↩]
- Pharmacokinetic analysis of capsaicin after topical administration of a high-concentration capsaicin patch to patients with peripheral neuropathic pain. Babbar S, Marier JF, Mouksassi MS, Beliveau M, Vanhove GF, Chanda S, Bley K. Ther Drug Monit. 2009 Aug; 31(4):502-10. https://www.ncbi.nlm.nih.gov/pubmed/19494795/[↩][↩][↩]
- Pharmacokinetic and the effect of capsaicin in Capsicum frutescens on decreasing plasma glucose level. Chaiyasit K, Khovidhunkit W, Wittayalertpanya S. J Med Assoc Thai. 2009 Jan; 92(1):108-13. https://www.ncbi.nlm.nih.gov/pubmed/19260251/[↩]
- Mozsik G. Capsaicin as new orally applicable gastroprotective and therapeutic drug alone or in combination with nonsteroidal anti-inflammatory drugs in healthy human subjects and in patients. Prog Drug Res 2014;68:209–58. https://www.ncbi.nlm.nih.gov/pubmed/24941671[↩][↩][↩]
- Sandor B, Papp J, Mozsik G et al. Orally given gastroprotective capsaicin does not modify aspirin-induced platelet aggregation in healthy male volunteers (human phase I examination). Acta Physiol Hung 2014;101:429–37. https://www.ncbi.nlm.nih.gov/pubmed/25532954[↩][↩][↩]
- Satyanarayana MN. Capsaicin and gastric ulcers. Crit Rev Food Sci Nutr 2006;46:275–328. https://www.ncbi.nlm.nih.gov/pubmed/16621751[↩]
- Pabalan N, Jarjanazi H, Ozcelik H. The impact of capsaicin intake on risk of developing gastric cancers: a meta-analysis. J Gastrointest Cancer 2014;45:334–41. https://www.ncbi.nlm.nih.gov/pubmed/24756832[↩]
- Bley K, Boorman G, Mohammad B et al. A comprehensive review of the carcinogenic and anticarcinogenic potential of capsaicin. Toxicol Pathol 2012;40:847–73. https://www.ncbi.nlm.nih.gov/pubmed/22563012[↩][↩][↩]
- Gaddy JA, Radin JN, Loh JT, et al. High Dietary Salt Intake Exacerbates Helicobacter pylori-Induced Gastric Carcinogenesis. Blanke SR, ed. Infection and Immunity. 2013;81(6):2258-2267. doi:10.1128/IAI.01271-12. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3676043/[↩]
- Vanilloid receptors–do they have a role in whole body metabolism? Evidence from TRPV1. Zsombok A. J Diabetes Complications. 2013 May-Jun; 27(3):287-92. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3633635/[↩]
- Gastrointestinal absorption and metabolism of capsaicin and dihydrocapsaicin in rats. Kawada T, Suzuki T, Takahashi M, Iwai K. Toxicol Appl Pharmacol. 1984 Mar 15; 72(3):449-56. https://www.ncbi.nlm.nih.gov/pubmed/6710495[↩]
- Vyklicky L, Novakova-Tousova K, Benedikt J et al. Calcium-dependent desensitization of vanilloid receptor TRPV1: a mechanism possibly involved in analgesia induced by topical application of capsaicin. Physiol Res 2008;57(Suppl 3):S59–68. https://www.ncbi.nlm.nih.gov/pubmed/18481914[↩]
- Kissin I. Vanilloid-induced conduction analgesia: selective, dose-dependent, long-lasting, with a low level of potential neurotoxicity. Anesth Analg 2008;107:271–81. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2497455/[↩]
- Jones VM, Moore KA, Peterson DM. Capsaicin 8% topical patch (Qutenza)—a review of the evidence. J Pain Palliat Care Pharmacother 2011;25:32–41. https://www.ncbi.nlm.nih.gov/pubmed/21426216[↩]
- Gunthorpe MJ, Szallasi A. Peripheral TRPV1 receptors as targets for drug development: new molecules and mechanisms. Curr Pharm Des 2008;14:32–41. https://www.ncbi.nlm.nih.gov/pubmed/18220816[↩]
- Czikora A, Rutkai I, Pasztor ET et al. Different desensitization patterns for sensory and vascular TRPV1 populations in the rat: expression, localization and functional consequences. PLoS ONE 2013;8:e78184. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3826751/[↩]
- Signal transduction of eNOS activation. Fleming I, Busse R. Cardiovasc Res. 1999 Aug 15; 43(3):532-41. https://www.ncbi.nlm.nih.gov/pubmed/10690325/[↩]
- Fluid shear stress stimulates phosphorylation-dependent nuclear export of HDAC5 and mediates expression of KLF2 and eNOS. Wang W, Ha CH, Jhun BS, Wong C, Jain MK, Jin ZG. Blood. 2010 Apr 8; 115(14):2971-9. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2854437/[↩]
- Integration of flow-dependent endothelial phenotypes by Kruppel-like factor 2. Parmar KM, Larman HB, Dai G, Zhang Y, Wang ET, Moorthy SN, Kratz JR, Lin Z, Jain MK, Gimbrone MA Jr, García-Cardeña G. J Clin Invest. 2006 Jan; 116(1):49-58. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1307560/[↩]
- Transient receptor potential vanilloid 1 activation by dietary capsaicin promotes urinary sodium excretion by inhibiting epithelial sodium channel α subunit-mediated sodium reabsorption. Li L, Wang F, Wei X, Liang Y, Cui Y, Gao F, Zhong J, Pu Y, Zhao Y, Yan Z, Arendshorst WJ, Nilius B, Chen J, Liu D, Zhu Z. Hypertension. 2014 Aug; 64(2):397-404. http://hyper.ahajournals.org/content/64/2/397.long[↩][↩]
- Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Zygmunt PM, Petersson J, Andersson DA, Chuang H, Sørgård M, Di Marzo V, Julius D, Högestätt ED. Nature. 1999 Jul 29; 400(6743):452-7. https://www.ncbi.nlm.nih.gov/pubmed/10440374/[↩]
- Structure-activity relationships of vanilloid receptor agonists for arteriolar TRPV1. Czikora Á, Lizanecz E, Bakó P, Rutkai I, Ruzsnavszky F, Magyar J, Pórszász R, Kark T, Facskó A, Papp Z, Édes I, Tóth A. Br J Pharmacol. 2012 Mar; 165(6):1801-1812. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3372831/[↩]
- Capsaicin, a spicy component of hot peppers, modulates adipokine gene expression and protein release from obese-mouse adipose tissues and isolated adipocytes, and suppresses the inflammatory responses of adipose tissue macrophages. Kang JH, Kim CS, Han IS, Kawada T, Yu R. FEBS Lett. 2007 Sep 18; 581(23):4389-96. https://www.ncbi.nlm.nih.gov/pubmed/17719033/[↩]
- Transient receptor potential vanilloid 1 activation enhances gut glucagon-like peptide-1 secretion and improves glucose homeostasis. Wang P, Yan Z, Zhong J, Chen J, Ni Y, Li L, Ma L, Zhao Z, Liu D, Zhu Z. Diabetes. 2012 Aug; 61(8):2155-65. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3402317/[↩]
- Non-pungent capsaicin analogs (capsinoids) increase metabolic rate and enhance thermogenesis via gastrointestinal TRPV1 in mice. Kawabata F, Inoue N, Masamoto Y, Matsumura S, Kimura W, Kadowaki M, Higashi T, Tominaga M, Inoue K, Fushiki T. Biosci Biotechnol Biochem. 2009 Dec; 73(12):2690-7. https://www.jstage.jst.go.jp/article/bbb/73/12/73_90555/_article[↩]
- Could capsaicinoids help to support weight management? A systematic review and meta-analysis of energy intake data. Whiting S, Derbyshire EJ, Tiwari B. Appetite. 2014 Feb; 73():183-8. https://www.ncbi.nlm.nih.gov/pubmed/24246368/[↩]
- Li L, Chen J, Ni Y et al. TRPV1 activation prevents nonalcoholic fatty liver through UCP2 upregulation in mice. Pflugers Arch 2012;463:727–32. https://www.ncbi.nlm.nih.gov/pubmed/22395410[↩]
- Planavila A, Rodriguez-Calvo R, Jove M et al. Peroxisome proliferator-activated receptor beta/delta activation inhibits hypertrophy in neonatal rat cardiomyocytes. Cardiovasc Res 2005;65:832–41. https://www.ncbi.nlm.nih.gov/pubmed/15721863[↩]
- Gao F, Liang Y, Wang X, et al. TRPV1 Activation Attenuates High-Salt Diet-Induced Cardiac Hypertrophy and Fibrosis through PPAR-δ Upregulation. PPAR Research. 2014;2014:491963. doi:10.1155/2014/491963. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4131514/[↩][↩]
- Echtay KS, Brand MD. 4-hydroxy-2-nonenal and uncoupling proteins: an approach for regulation of mitochondrial ROS production. Redox Rep 2007;12:26–9. https://www.ncbi.nlm.nih.gov/pubmed/17263904[↩]
- Tian XY, Wong WT, Xu A et al. Uncoupling protein-2 protects endothelial function in diet-induced obese mice. Circ Res 2012;110:1211–16. http://circres.ahajournals.org/content/110/9/1211.long[↩]
- Sun J, Pu Y, Wang P, et al. TRPV1-mediated UCP2 upregulation ameliorates hyperglycemia-induced endothelial dysfunction. Cardiovascular Diabetology. 2013;12:69. doi:10.1186/1475-2840-12-69. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3644255/[↩][↩]
- Serviddio G, Bellanti F, Tamborra R et al. Uncoupling protein-2 (UCP2) induces mitochondrial proton leak and increases susceptibility of non-alcoholic steatohepatitis (NASH) liver to ischaemia-reperfusion injury. Gut 2008;57:957–65. https://www.ncbi.nlm.nih.gov/pubmed/18308829[↩]
- Li Q, Li L, Wang F et al. Dietary capsaicin prevents nonalcoholic fatty liver disease through transient receptor potential vanilloid 1-mediated peroxisome proliferator-activated receptor delta activation. Pflugers Arch 2013;465:1303–16. https://www.ncbi.nlm.nih.gov/pubmed/23605066[↩][↩][↩]
- McCarty MF. High mitochondrial redox potential may promote induction and activation of UCP2 in hepatocytes during hepatothermic therapy. Med Hypotheses 2005;64:1216–19. https://www.ncbi.nlm.nih.gov/pubmed/15823721[↩]
- Ono K, Tsukamoto-Yasui M, Hara-Kimura Y et al. Intragastric administration of capsiate, a transient receptor potential channel agonist, triggers thermogenic sympathetic responses. J Appl Physiol (1985) 2011;110:789–98. http://jap.physiology.org/content/110/3/789.long[↩]
- Whiting S, Derbyshire EJ, Tiwari B. Could capsaicinoids help to support weight management? A systematic review and meta-analysis of energy intake data. Appetite 2014;73:183–8. https://www.ncbi.nlm.nih.gov/pubmed/24246368[↩]
- Shirai Y, Ueno S, Nakayama A et al. Studies of the toxicological potential of capsinoids, XII: pharmacokinetic study of capsinoid-containing CH-19 Sweet extract in rats. Int J Toxicol 2010;29(2 Suppl):15S–21S. https://www.ncbi.nlm.nih.gov/pubmed/20388820[↩][↩]
- Iida T, Moriyama T, Kobata K et al. TRPV1 activation and induction of nociceptive response by a non-pungent capsaicin-like compound, capsiate. Neuropharmacology 2003;44:958–67. https://www.ncbi.nlm.nih.gov/pubmed/12726827[↩]
- Snitker S, Fujishima Y, Shen H et al. Effects of novel capsinoid treatment on fatness and energy metabolism in humans: possible pharmacogenetic implications. Am J Clin Nutr 2009;89:45–50. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3151435/[↩]
- Yoneshiro T, Aita S, Kawai Y et al. Nonpungent capsaicin analogs (capsinoids) increase energy expenditure through the activation of brown adipose tissue in humans. Am J Clin Nutr 2012;95:845–50. http://ajcn.nutrition.org/content/95/4/845.long[↩]
- Yoneshiro T, Saito M. Transient receptor potential activated brown fat thermogenesis as a target of food ingredients for obesity management. Curr Opin Clin Nutr Metab Care 2013;16:625–31. https://www.ncbi.nlm.nih.gov/pubmed/24100669[↩]
- Ludy M-J, Moore GE, Mattes RD. The Effects of Capsaicin and Capsiate on Energy Balance: Critical Review and Meta-analyses of Studies in Humans. Chemical Senses. 2012;37(2):103-121. doi:10.1093/chemse/bjr100. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3257466[↩]
- Effect of capsaicin on substrate oxidation and weight maintenance after modest body-weight loss in human subjects. Lejeune MP, Kovacs EM, Westerterp-Plantenga MS. Br J Nutr. 2003 Sep; 90(3):651-59. https://www.ncbi.nlm.nih.gov/pubmed/13129472/[↩]
- Tolerability of NGX-4010, a capsaicin 8% patch for peripheral neuropathic pain. Peppin JF, Majors K, Webster LR, Simpson DM, Tobias JK, Vanhove GF. J Pain Res. 2011; 4():385-92. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3255992/[↩]
- Ching LC, Kou YR, Shyue SK et al. Molecular mechanisms of activation of endothelial nitric oxide synthase mediated by transient receptor potential vanilloid type 1. Cardiovasc Res 2011;91:492–501. https://www.ncbi.nlm.nih.gov/pubmed/21493704[↩]
- Xu X, Wang P, Zhao Z et al. Activation of transient receptor potential vanilloid 1 by dietary capsaicin delays the onset of stroke in stroke-prone spontaneously hypertensive rats. Stroke 2011;42:3245–51. http://stroke.ahajournals.org/content/42/11/3245.long[↩]
- Fragasso G, Palloshi A, Piatti PM et al. Nitric-oxide mediated effects of transdermal capsaicin patches on the ischemic threshold in patients with stable coronary disease. J Cardiovasc Pharmacol 2004;44:340–7. https://www.ncbi.nlm.nih.gov/pubmed/15475832[↩]
- Li L, Wang F, Wei X et al. Transient receptor potential vanilloid 1 activation by dietary capsaicin promotes urinary sodium excretion by inhibiting epithelial sodium channel alpha subunit-mediated sodium reabsorption. Hypertension 2014;64:397–404. http://hyper.ahajournals.org/content/64/2/397.long[↩]
- Chen J, Li L, Li Y, et al. Activation of TRPV1 channel by dietary capsaicin improves visceral fat remodeling through connexin43-mediated Ca2+ Influx. Cardiovascular Diabetology. 2015;14:22. doi:10.1186/s12933-015-0183-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4340344/[↩]
- Snitker S, Fujishima Y, Shen H, et al. Effects of novel capsinoid treatment on fatness and energy metabolism in humans: possible pharmacogenetic implications. The American Journal of Clinical Nutrition. 2009;89(1):45-50. doi:10.3945/ajcn.2008.26561. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3151435/[↩]
- McCarty MF, DiNicolantonio JJ, O’Keefe JH. Capsaicin may have important potential for promoting vascular and metabolic health. Open Heart. 2015;2(1):e000262. doi:10.1136/openhrt-2015-000262. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4477151/[↩]


{kind=link}