Contents
What is phosphatidylserine
Phosphatidylserine (1, 2-diacyl-sn-glycero-3-phospho-L-serine) is the major anionic phospholipid in mammalian cell membranes and phosphatidylserine accounts for 13–15 % of the phospholipids in the human cerebral cortex 1. The brain is enriched in the two aminophospholipids, phosphatidylethanolamine and phosphatidylserine compared with other tissues. In the brain, and particularly in the retina 2, the acyl chains of phosphatidylserine are highly enriched in docosahexaenoic acid (DHA) 3. In human gray matter, docosahexaenoic acid (DHA) accounts for >36% of the fatty acyl species of phosphatidylserine 4. Phosphatidylserine has been shown to play a key role in the functioning of neuron membranes, such as signal transduction, secretory vesicle release and cell-to-cell communication 5. With aging, neural membrane fluidity is compromised due to the increased presence of cholesterol, a low incorporation rate and decreased levels of total polyunsaturated fatty acids, blockages to phospholipid pathways, and increases in free radicals, resulting in oxidative stress 6. At least 25 clinical trials suggested that consumption of phosphatidylserine supplement may reduce the risk of dementia and cognitive dysfunction in the elderly 7, 8. The administration of phosphatidylserine extracted from bovine cortex (bovine cortex-phosphatidylserine) has positive effects on brain function. Bovine cortex-phosphatidylserine was shown to improve learning and memory in age-associated memory impaired subjects 9, to enhance behavioral and cognitive parameters in geriatric patients 10, and to improve cognitive performance of Alzheimer’s disease patients 11. However, improvements lasted only a few months and were seen in people with the least severe symptoms. Initially, phosphatidylserine supplements were derived from the brain cells of cows (bovine cortex-phosphatidylserine). The bovine brain cortex phosphatidylserine was found to be enriched with docosahexaenoic acid (DHA) 12. But because of concerns about mad cow disease, most manufacturers now produce the supplements from soy lecithin, sunflower lecithin or cabbage derivatives. However, soybean-derived phosphatidylserine does not contain docosahexaenoic acid (DHA), and clinical studies have demonstrated inconclusive efficacy results 13. Preliminary studies have shown that plant-based phosphatidylserine supplements may also offer benefits, but more research is needed. However, no modern studies have continued to focus on phosphatidylserine, suggesting its limited effect.
Phosphatidylserine supplement has been approved by the U.S. Food and Drug Administration 14 to treat memory deficit disorders such as Alzheimer’s disease and other forms of dementia, to support healthy cognitive function during aging, and to remediate cognitive deficits as a result of heavy drinking and cigarette smoking. The mechanism by which phosphatidylserine- omega-3 (DHA) exerts its effects is not fully understood, however phosphatidylserine has been found to regulate key proteins in neuronal membranes, including sodium/calcium ATPase 15, protein kinase C 16 and Raf-1 protein kinase 17. Phosphatidylserine was also found to influence neurotransmitter activity, such as the release of acetylcholine, dopamine and noradrenaline 18. In addition, phosphatidylserine-omega-3 was found to significantly increase DHA level in brains of middle-aged rats 19.
Mammalian cell membranes contain >1,000 different phospholipids 20. Phosphatidylcholine is the most abundant phospholipid in mammalian cell membranes, constituting 40–50% of total phospholipids. The second most abundant mammalian membrane phospholipid is phosphatidylethanolamine (PE), which constitutes 20–50% of total phospholipids. In the brain, ∼45% of total phospholipids are phosphatidylethanolamine, whereas in the liver, only ∼20% of total phospholipids are phosphatidylethanolamine. Phosphatidylserine is a quantitatively minor membrane phospholipid that makes up 2–10% of total phospholipids.
In the plasma membrane, phosphatidylserine is localized exclusively in the cytoplasmic leaflet where it forms part of protein docking sites necessary for the activation of several key signaling pathways. These include the Akt, protein kinase C (PKC) and Raf-1 signaling that is known to stimulate neuronal survival, neurite growth and synaptogenesis 21. Modulation of the phosphatidylserine level in the plasma membrane of neurons has significant impact on these signaling processes. The mechanism of phosphatidylserine-mediated activation of these neuronal signaling pathways is illustrated in Figure 2.
Figure 1. Phosphatidylserine
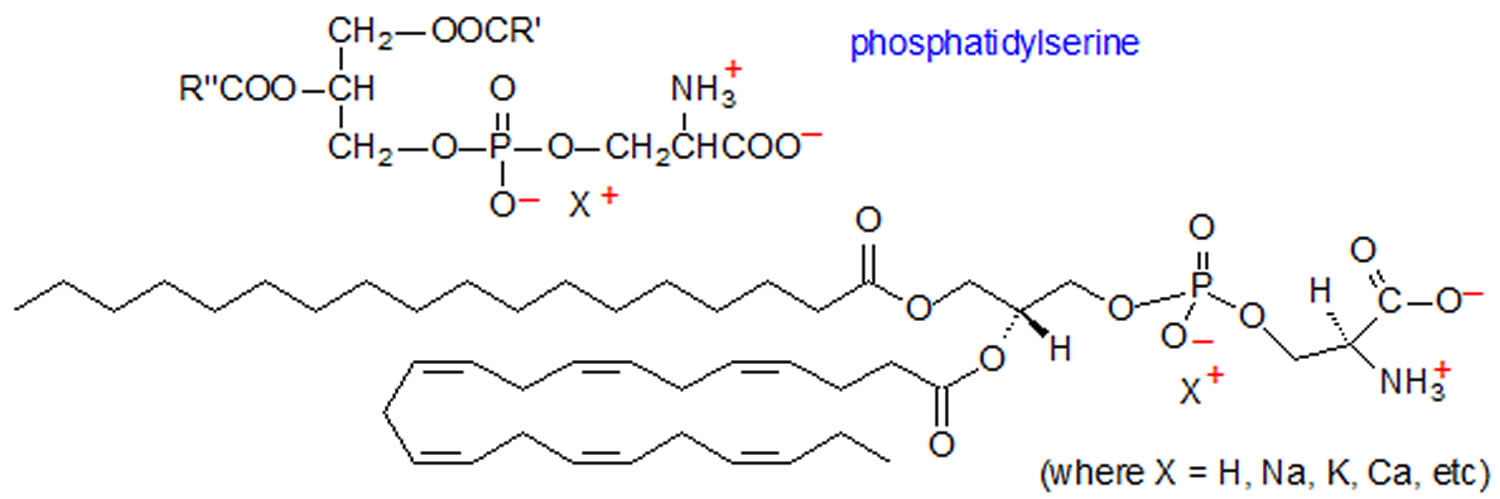
Figure 2. Activation of neuronal signaling pathways facilitated by phosphatidylserine
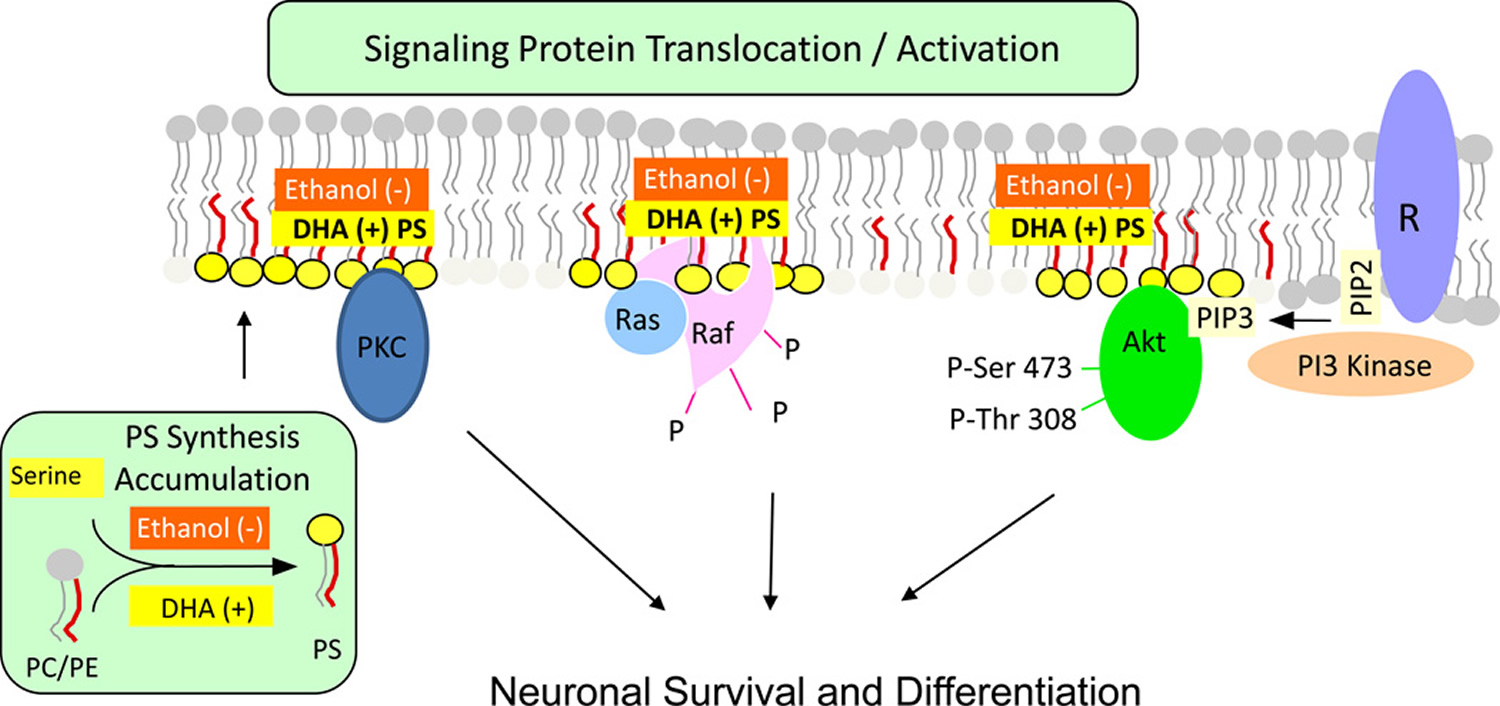
Footnotes: Activation of neuronal signaling pathways facilitated by phosphatidylserine. Activation of Akt, protein kinase C and Raf-1 requires translocation from the cytosol to the cytoplasmic surface of the plasma membrane. Translocation is initiated by specific stimuli, for example, growth factor-dependent PIP3 generation from PIP2 by PI3 kinase in the case of Akt. Binding to the membrane occurs in part through an interaction of these proteins with phosphatidylserine present in anionic domains of the lipid bilayer, activating the signaling pathways leading to neuronal differentiation and survival. DHA facilitates this mechanism by increasing phosphatidylserine production in neurons, while ethanol has the opposite effect because it inhibits the DHA-induced increase in PS production.
Abbreviations: PS =phosphatidylserine; R = receptor; DHA = docosahexaenoic acid; PC = phosphatidylcholine; PE = phosphatidylethanolamine
[Source 22]Phosphatidylserine biological function
Phosphatidylserine, a phospholipid with a negatively charged head-group, is an important constituent of eukaryotic cellular membranes. On the plasma membrane, rather than being evenly distributed, phosphatidylserine is found preferentially in the inner leaflet. Disruption of this asymmetry, leading to the appearance of phosphatidylserine on the surface of the cell, is known to play a central role in both apoptosis and blood clotting.
The best-studied roles of phosphatidylserine involve signalling, not within the intracellular environment, but in an extracellular context such as during apoptosis 23 and during blood clotting. Like most lipids, phosphatidylserine is not evenly distributed throughout all cellular membranes, nor is it always equally distributed between leaflets of a membrane bilayer 24. In healthy cells plasmalemmal phosphatidylserine is exclusively on the inner (cytoplasmic-facing) leaflet due to the action of ATP-dependent aminophospholipid flippases 25. When cells undergo apoptosis (regulated cell death) phosphatidylserine appears on the outside-facing (extracellular) leaflet, signalling phagocytic cells to engulf the dying cell. phosphatidylserine is also exposed exofacially in activated blood platelets, which prompts the binding and activation of a number of clotting factors, including factors V, VIII, X and prothrombin 26.
There is little doubt that, in addition to these extracellular functions, phosphatidylserine plays important roles within the intracellular environment. Indeed, a number of important intracellular proteins require phosphatidylserine for their proper localization and/or activation. Such proteins include the E3 ubiquitin-protein ligase NEDD4, a number of protein kinase C isoforms, several phospholipase C and D isoforms, PTEN ,an important phosphatidylinositol (3,4,5)-trisphosphate phosphatase, dysferlin, a protein important in muscle repair, as well as a number of synaptotagmin isoforms that are important for vesicular trafficking and fusion 27. Additionally, it is known on the whole that phosphatidylserine is important, as mice with a complete loss of ability to synthesize phosphatidylserine are not viable 28, and though yeast are able to survive without phosphatidylserine synthesis, their growth is greatly impaired 29. However, despite the obvious importance of intracellular phosphatidylserine, its distribution, dynamics and function have not been thoroughly investigated.
Cell signaling
Phosphatidylserine(s) are actively held facing the cytosolic (inner) side of the cell membrane by the enzyme flippase. However, when a cell undergoes apoptosis, phosphatidylserine is no longer restricted to the cytosolic side by flippase. Instead scramblase catalyzes the rapid exchange of phosphatidylserine between the two sides. When the phosphatidylserines flip to the extracellular (outer) surface of the cell, they act as a signal for macrophages to engulf the cells 30.
Coagulation
Phosphatidylserine plays a role in blood coagulation (also known as clotting). When circulating platelets encounter the site of an injury, collagen and thrombin -mediated activation causes externalization of phosphatidylserine from the inner membrane layer, where it serves as a pro-coagulant surface 31. This surface acts to orient coagulation proteases, specifically tissue factor and factor VII, facilitating further proteolysis, activation of factor X, and ultimately generating thrombin 31.
In the coagulation disorder Scott syndrome, the mechanism in platelets for transportation of phosphatidylserine from the inner platelet membrane surface to the outer membrane surface is defective 32. It is characterized as a mild bleeding disorder stemming from the patient’s deficiency in thrombin synthesis 33.
Ethanol-induced effects
Findings in cultured neuronal cells and animal models indicate that ethanol can disrupt the beneficial interaction between DHA and phosphatidylserine. Exposure of Neuro 2A cells to 25 mM ethanol decreased the DHA-mediated accumulation of phosphatidylserine, Akt phosphorylation and cell survival as indicated in Figure 2 above. Administration of ethanol to pregnant rats also decreased the DHA and phosphatidylserine content of the fetal hippocampus and increased the number of apoptotic hippocampal cells 34. Likewise, administration of ethanol to rats during the prenatal and developmental period decreased microsomal phosphatidylserine biosynthetic activity without altering phosphatidylserine synthetase expression, and thus the amount of phosphatidylserine, particularly the 18:0, DHA species in the cerebral cortex 35. The ethanol-mediated decrease in the 18:0, DHA-phosphatidylserine molecular species is also consistent with the finding that incubation of brain microsomes with high concentrations of ethanol increased oleoyl-CoA incorporation into phosphatidylserine and diverted polyunsaturated fatty acids into triglycerides 36. Considering the significant roles played by phosphatidylserine in neuronal survival and function discussed above, a reduction in DHA-stimulated synthesis of phosphatidylserine may be one factor that produces the deleterious effect of ethanol on the central nervous system.
Phosphatidylserine foods
Phosphatidylserine is commonly found in common foods such as meat, fish and legumes,. Currently, dietary intakes of phosphatidylserine, from its natural presence in the diet, is estimated to be in the range of 75 – 184 mg/person/day.
Table 1. Phosphatidylserine content in different foods
| Food | Phosphatidylserine Content in mg/100 g |
|---|---|
| Bovine brain | 713 |
| Atlantic mackerel | 480 |
| Chicken heart | 414 |
| Atlantic herring | 360 |
| Eel | 335 |
| Offal (average value) | 305 |
| Pig’s spleen | 239 |
| Pig’s kidney | 218 |
| Tuna | 194 |
| Chicken leg, with skin, without bone | 134 |
| Chicken liver | 123 |
| White beans | 107 |
| Soft-shell clam | 87 |
| Chicken breast, with skin | 85 |
| Mullet | 76 |
| Veal | 72 |
| Beef | 69 |
| Pork | 57 |
| Pig’s liver | 50 |
| Turkey leg, without skin or bone | 50 |
| Turkey breast without skin | 45 |
| Crayfish | 40 |
| Cuttlefish | 31 |
| Atlantic cod | 28 |
| Anchovy | 25 |
| Whole grain barley | 20 |
| European hake | 17 |
| European pilchard (sardine) | 16 |
| Trout | 14 |
| Rice (unpolished) | 3 |
| Carrot | 2 |
| Ewe’s Milk | 2 |
| Cow’s Milk (whole, 3.5% fat) | 1 |
| Potato | 1 |
Phosphatidylserine synthesis in the brain
In mammalian tissues, phosphatidylserine is synthesized from either phosphatidylcholine or phosphatidylethanolamine exclusively by Ca2+-dependent reactions where the head group of the substrate phospholipids is replaced by serine 38, as illustrated in Figure 3. These base-exchange reactions are catalyzed by phosphatidylserine synthases (PSS) and so far two isoforms, PSS1 and PSS2 encoded by two separate genes, Pss1 and Pss2, respectively, have been identified. PSS1 utilizes phosphatidylcholine as its substrate, and PSS2 utilizes phosphatidylethanolamine. These enzymes are localized in the endoplasmic reticulum, particularly enriched in the mitochondria associated membrane regions of the endoplasmic reticulum 39.
Together with testis and kidney, brain is one of the tissues that have high capacity to synthesize phosphatidylserine 40. Also, the expression of phosphatidylserine synthases (PSS) in the brain is among the highest. The serine base exchange enzymatic activities of rat cerebellar homogenates, cerebral cortical homogenates and cerebral cortical membranes were shown to be recovered in the insoluble floating fraction of TritonX-100 extracts, suggesting the localized presence of phosphatidylserine synthases in membrane lipid rafts 41. Although intriguing, the possible contribution of microsomal contamination cannot be excluded. Phosphatidylserine production is increased in cells of neuronal origin by compounds that trigger Ca2+ release, a finding consistent with the fact that phosphatidylserine synthesis is a Ca2+-dependent process 42.
Figure 3. Phosphatidylserine synthesis and metabolism in the brain
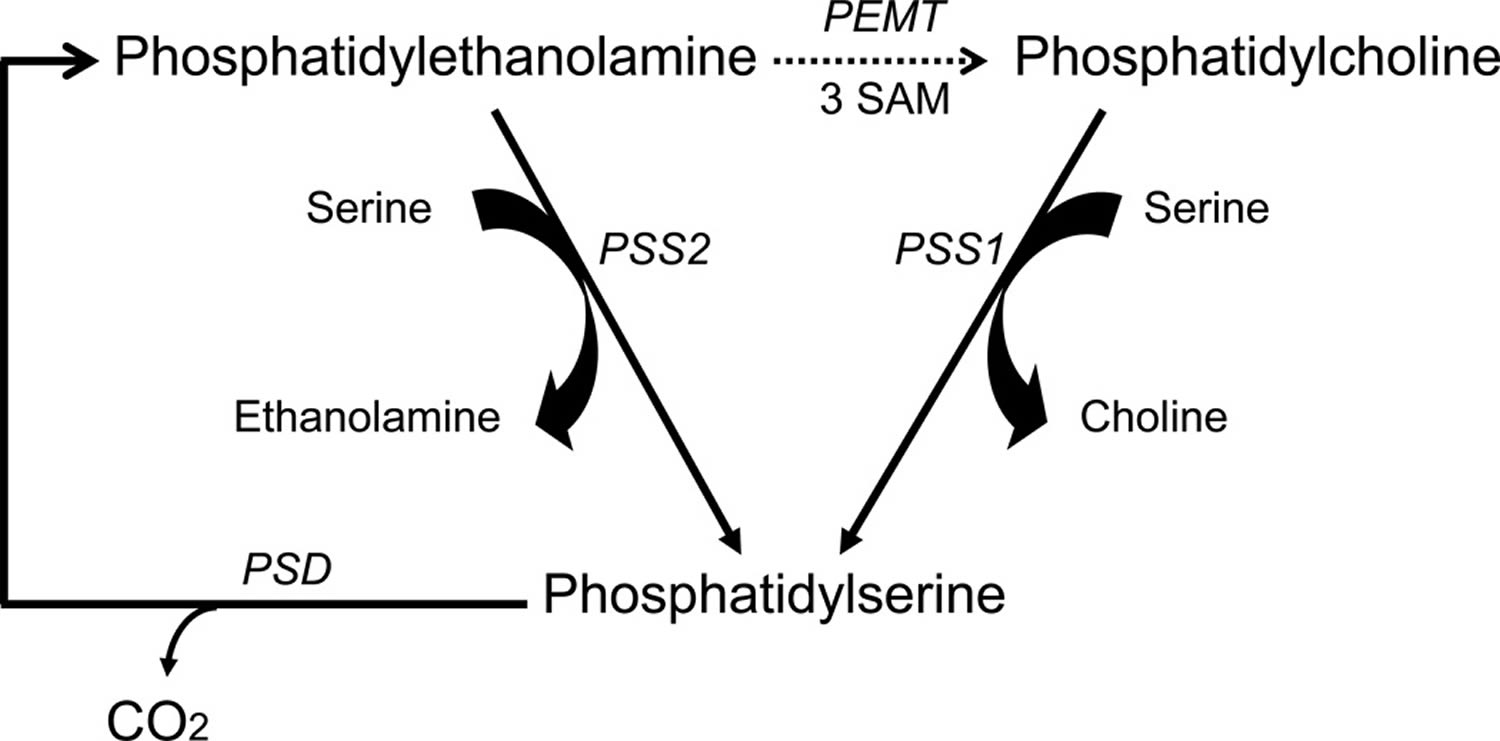
Footnotes: Phosphatidylserine synthesis and metabolism in the brain. Phosphatidylserine is synthesized by replacement of the choline group of phosphatidylcholine by serine in a reaction catalyzed by phosphatidylserine synthase 1, and also by replacement of the ethanolamine group of phosphatidylethanolamine by serine in a reaction catalyzed by phosphatidylserine synthase 2. These synthetic reactions occur in the endoplasmic reticulum. phosphatidylserine is decarboxylated to phosphatidylethanolamine in the mitochondria by phosphatidylserine decarboxylase (PSD). The phosphatidylethanolamine methyltransferase (PEMT) reaction that utilizes S-adenosylmethionine (SAM) to convert phosphatidylethanolamine to phosphatidylcholine is indicated as a dashed arrow because more recent findings have demonstrated that previously reported methylation activity in the brain is quantitatively insignificant 43.
Abbreviations: PC = phosphatidylcholine; PE = phosphatidylethanolamine; PS = phosphatidylserine; PSD = phosphatidylserine decarboxylase; PSS = phosphatidylserine synthases
[Source 22]Composition of brain phosphatidylserine
The phosphatidylserine content in human brain is maintained at the 13–14% level throughout the life 44. The fatty acid composition of phosphatidylserine, and for comparison that of phosphatidylethanolamine and phosphatidylcholine, contained in the gray and white matter of human brain, is shown in Table 2 44. There are substantial differences in the fatty acid composition in gray and white matter phosphatidylserine. Gray matter phosphatidylserine contains considerably more DHA and less 18:1n-9 than white matter. Appreciable differences also occur in the phosphatidylethanolamine fatty acid composition in gray and white matter, whereas comparatively small differences occur in the phosphatidylcholine composition. Docosahexaenoic acid (DHA) accounts for more than one-third of the total fatty acid and 80% of the polyunsaturated fatty acid in gray matter phosphatidylserine. A substantial amount of DHA is present in gray matter phosphatidylethanolamine, but only a relatively small amount is present in phosphatidylcholine. Gray matter phosphatidylserine and phosphatidylethanolamine contain considerably more 18:0 and much less 16:0 than phosphatidylcholine. Arachidonic acid is present primarily in phosphatidylethanolamine, and there is little arachidonic acid in phosphatidylserine. According to one study 45, phosphatidylethanolamine also contains the largest amount of docosapentaenoic acid, an arachidonic acid-derived product. Only trace amounts of linoleic acid (18:2n-6) are present in brain phosphatidylserine, phosphatidylethanolamine and phosphatidylcholine.
Table 2. Fatty acid composition of human brain glycerophosphatides
| Fatty acid | Fatty Acyl Composition (% of total fatty acid in each phospholipid class) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phosphatidylserine | Phosphatidylethanolamine | Phosphatidylcholine | ||||||||||
| Gray matter | White matter | Gray matter | White matter | Gray matter | White matter | |||||||
| Aa | Bb | A | B | A | B | A | B | A | B | A | B | |
| 16:0 | 2.3 | 4.2 | 1.7 | 1.9 | 6.7 | 5.7 | 6.7 | 3.4 | 45.0 | 45.9 | 34.3 | 30.2 |
| 16:1 | 0.3 | 0.3 | 0.4 | 0.3 | 0.4 | 0.4 | 1.4 | 0.5 | 3.1 | 2.4 | 1.0 | 2.3 |
| 18:0 | 25.4 | 45.3 | 35.8 | 44.1 | 26.0 | 28.4 | 9.0 | 9.3 | 9.3 | 11.2 | 13.4 | 12.9 |
| 18:1 | 21.5 | 14.0 | 39.7 | 41.4 | 11.9 | 10.3 | 42.4 | 38.9 | 31.4 | 30.3 | 45.2 | 47.1 |
| 20:4 | 1.6 | 3.0 | 2.0 | 1.2 | 13.8 | 11.2 | 6.4 | 16.5 | 4.1 | 3.6 | 1.3 | 0.8 |
| 22:5n-6 | 5.0 | 0.7 | 4.8 | 0.1 | 14.3 | 1.2 | 13.7 | 0.7 | ndc | trd | nd | 0.2 |
| 22:5n-3 | 3.3 | 0.5 | 0.9 | 0.2 | tr | 1.1 | 0.5 | 0.9 | nd | tr | 0.3 | 0.3 |
| 22:6 | 36.6 | 23.2 | 5.6 | 1.3 | 24.3 | 30.5 | 3.4 | 8.6 | 3.1 | 2.5 | 0.1 | 0.2 |
Interactions between phosphatidylserine and DHA
Brain is enriched with DHA (docosahexaenoic acid), and the phosphatidylserine content in the grey matter of human brain is particularly high as shown in Table 1. Phosphatidylcholine and phosphatidylethanolamine containing DHA are the best substrate for phosphatidylserine biosynthesis 46. As a consequence, the phosphatidylserine level is high in the brain where DHA is abundant. In contrast, DHA-depletion from the brain lowers the phosphatidylserine level 47. For example, dietary depletion of Omega-3 fatty acids can reduce DHA in the brain and increase the Omega-6 counterpart docosapentaenoic acid. Since phospholipids containing docosapentaenoic acid are not as good substrates for phosphatidylserine synthesis as DHA (docosahexaenoic acid)-containing phospholipid species, the brain phosphatidylserine level is reduced 46. It is important to note that modern western diets are excessively abundant in Omega-6 fatty acids compared to Omega-3 fatty acids, and therefore, DHA enrichment in the brain is compromised as indicated by the considerable accumulation of docosapentaenoic acid observed in the postmortem human hippocampus 48. Such a fatty acid profile is comparable to that observed in rodent brains depleted in DHA at least to a moderate extent, and therefore, the phosphatidylserine level is likely not at a maximum in modern human brains although systematic analysis has not yet been performed.
Phospahtidylserine and cognitive function
A decrease of the DHA content in phosphatidylserine has been reported in cognitive impairment. A small reduction in the DHA content of hippocampal phosphatidylserine was observed in 12 month-old senescence-accelerated prone mice that have a shorter life span, learning and memory deficit, and an increase in hippocampal Aβ-peptide content 49. The decrease in DHA was associated with a corresponding increase in the AA content of hippocampal phosphatidylserine. Likewise, the DHA content of phosphatidylserine in the superior temporal and mid-frontal cortex was reduced by 12 and 14 %, respectively, in brain tissues obtained from patients with Alzheimer’s disease 50. However, there is no information as to how a decrease in the DHA content of phosphatidylserine might contribute to the pathogenesis of cognitive impairment, and a decrease in DHA content is not a uniform finding in animal models of cognitive impairment. For example, substantial fatty acyl compositional changes, including reductions in AA, have been observed in brain phosphatidylserine of aged Wistar rats with cognitive deficits, but there is no difference in the DHA content of the phosphatidylserine 51. Therefore, the putative linkage between DHA reductions in phosphatidylserine and cognitive impairment remains open to question.
Effects of dietary phosphatidylserine supplements on cognitive function
Dietary phosphatidylserine supplements are reported to improve cognitive function in experimental animals 52, and a similar result has been obtained recently with krill phosphatidylserine which has a high content of omega-3 fatty acid. Aged rats given daily doses of krill phosphatidylserine orally for 7 days showed improvement in the Morris water maze test. There was less loss of choline acetyltransferase and acetylcholine esterase transporter mRNA in the hippocampus. The neuroprotective activity of 20 mg/kg krill phosphatidylserine was equivalent to that of 50 mg/kg soy phosphatidylserine in these aged rats 53. Normal young rats given 100 mg/kg krill phosphatidylserine orally for 30 days also showed improvement in the Morris water maze test 54.
Likewise, cognitive improvement was reported in humans given oral phosphatidylserine supplements 55, and these findings subsequently were confirmed and extended. Human subjects treated for 42 days with 200 mg of soy-based phosphatidylserine showed a more relaxed state before and after mental stress as measured by electro-encephalography 56, and no adverse effects were evident at this dose given three-times a day for 6 to 12 weeks 57. The ability to recall words increased by 42% in male and female subjects who were older than 60 years and complained of subjective memory loss when they were treated with 300 mg/day of phosphatidylserine containing 37.5 mg of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) 58. Improved verbal immediate recall was also observed in a double-blind, placebo controlled clinical trial in a large group of elderly subjects with memory complaints when treated with a daily dose of 300 mg phosphatidylserine containing DHA and eicosapentaenoic acid in a 3:1 ratio. A subset with relatively good cognitive performance at baseline showed the greatest improvement 59. In a similar double blind study in Japanese subjects between the ages of 50 and 69 years with memory complaints, subjects having low scores at baseline showed greater improvement in delayed verbal recall after treatment for 6 months with soybean phosphatidylserine 60.
Several biochemical responses to phosphatidylserine administration that have been reported in experimental animals could be involved in the mechanism of phosphatidylserine-mediated improvement in cognition. Stimulation of dopamine-dependent adenylate cyclase activity was observed in mouse brain following an intravenous infusion of a sonicated preparation of bovine brain phosphatidylserine 61. Sonicated suspensions of phosphatidylserine injected intravenously also increased calcium-dependent acetylcholine output from the cerebral cortex in urethane anesthetized rats 62. Likewise, intravenous injection of purified bovine brain phosphatidylserine for 8 days attenuated the decrease in acetylcholine release from the parietal cortex in aged rats, possibly by providing more choline for acetylcholine synthesis 63. Furthermore, orally administered krill phosphatidylserine in normal young rats for 30 days produced an increase in neurons positive for brain-derived neurotrophic factor and insulin-like growth factor in the hippocampal CA1 region 54.
A current hypothesis is that these biochemical responses and the resulting cognitive improvements are due to phosphatidylserine mediated effects on neuronal membrane properties 55. However, experimental evidence indicating that orally or intravenously administered phosphatidylserine actually alters neuronal membrane properties is lacking. How the administered phosphatidylserine is transported in the plasma, how much enters the brain, whether it is taken up intact, and whether it is incorporated into neurons or glia are not known. Dietary phospholipids are hydrolyzed during digestion, so orally administered phosphatidylserine most likely is not absorbed intact. Phosphatidylserine preparations are rich in DHA, and DHA supplementation is known to improve hippocampal function 64. Because the administered phosphatidylserine probably undergoes partial or complete hydrolysis, the beneficial effects of phosphatidylserine on cognition, particularly from krill or bovine sources, possibly are produced by DHA released from the phosphatidylserine rather than the intact phosphatidylserine itself. These issues will have to be investigated in order to obtain some mechanistic insight into how dietary or intravenously administered phosphatidylserine supplements function to produce cognitive improvement.
Phosphatidylserine summary
A body of evidence supports the functional significance of phosphatidylserine in the brain. Underlying mechanisms are still unfolding; however, phosphatidylserine facilitates the activation of signaling proteins and receptors that are critical for neuronal survival, differentiation and synaptic neurotransmission. Despite its constitutive nature, membrane phosphatidylserine is often an indispensable participant in signaling events and/or influences the signaling in a concentration-dependent manner. The phosphatidylserine biosynthesis preferentially utilizes DHA-containing phospholipids as substrates. Although membrane phospholipids are under a tight homeostatic regulation, the phosphatidylserine level can be altered according to the DHA status, specifically in the brain. Therefore, diet- or ethanol-induced alteration of the brain DHA level and membrane phosphatidylserine can influence the signaling platform in the membrane and the transmission of the signaling cues. Detailed molecular mechanisms, particularly membrane phosphatidylserine-protein interactions, warrant further investigation in order to obtain more insight into the functional significance of neuronal phosphatidylserine. Such endeavor is likely to generate new targets for controlling physiologic or pathophysiologic processes affecting brain function.
Phosphatidylserine dosage
A typical recommended dose of phosphatidylserine as a dietary supplement is 100 mg three times a day (300 mg/day). In numerous human clinical studies, the safety of phosphatidylserine has been confirmed at daily doses of up to 300 mg for up to 6 months 65. The safety of phosphatidylserine has been proven in human clinical studies including susceptible groups (elderly and children) and healthy individuals.
Phosphatidylserine side effects
Phosphatidylserine has been marketed as a dietary supplement for the past two decades without any adverse effects (except gastrointestinal side effects such as nausea and indigestion). A variety of animal toxicity studies and in vitro mutagenicity/genotoxicity studies corroborate the human clinical safety data. The animal studies did not show any significant toxicity at doses up to approximately 1,000 mg/kg/day 66.
- Distribution and fatty acid composition of phosphoglycerides in normal human brain. Svennerholm L. J Lipid Res. 1968 Sep; 9(5):570-9. http://www.jlr.org/content/9/5/570.long[↩]
- Ford, D. A., J. K. Monda, R. S. Brush, R. E. Anderson, M. J. Richards, and S. J. Fliesler. 2008. Lipidomic analysis of the retina in a rat model of Smith-Lemli-Opitz syndrome: alterations in docosahexaenoic acid content of phospholipid molecular species. J. Neurochem.[↩]
- Kim, H. Y. 2007. Novel metabolism of docosahexaenoic acid in neural cells. J. Biol. Chem. 282: 18661–18665.[↩]
- Kim, H. Y., J. Bigelow, and J. H. Kevala. 2004. Substrate preference in phosphatidylserine biosynthesis for docosahexaenoic acid containing species. Biochemistry. 43: 1030–1036.[↩]
- Metabolism and functions of phosphatidylserine. Vance JE, Steenbergen R. Prog Lipid Res. 2005 Jul; 44(4):207-34.[↩]
- Yehuda S, Rabinovitz S, Carasso RL, Mostofsky DI: The role of polyunsaturated fatty acids in restoring the aging neuronal membrane. Neurobiol Aging 2002;23:843-853.[↩]
- The influence of soy-derived phosphatidylserine on cognition in age-associated memory impairment. Jorissen BL, Brouns F, Van Boxtel MP, Ponds RW, Verhey FR, Jolles J, Riedel WJ. Nutr Neurosci. 2001; 4(2):121-34. https://www.ncbi.nlm.nih.gov/pubmed/11842880/[↩]
- An open trial of plant-source derived phosphatydilserine for treatment of age-related cognitive decline. Schreiber S, Kampf-Sherf O, Gorfine M, Kelly D, Oppenheim Y, Lerer B. Isr J Psychiatry Relat Sci. 2000; 37(4):302-7. https://www.ncbi.nlm.nih.gov/pubmed/11201936/[↩]
- Effects of phosphatidylserine in age-associated memory impairment. Crook TH, Tinklenberg J, Yesavage J, Petrie W, Nunzi MG, Massari DC. Neurology. 1991 May; 41(5):644-9.[↩]
- Cognitive decline in the elderly: a double-blind, placebo-controlled multicenter study on efficacy of phosphatidylserine administration. Cenacchi T, Bertoldin T, Farina C, Fiori MG, Crepaldi G. Aging (Milano). 1993 Apr; 5(2):123-33.[↩]
- Effects of phosphatidylserine in Alzheimer’s disease. Crook T, Petrie W, Wells C, Massari DC. Psychopharmacol Bull. 1992; 28(1):61-6.[↩]
- Kim HY, Akbar M, Kim YS: Phosphatidylserine-dependent neuroprotective signaling promoted by docosahexaenoic acid. Prostaglandins Leukot Essent Fatty Acids 2010;82:165-172.[↩]
- Jorissen BL, Brouns F, Van Boxtel MP, Ponds RW, Verhey FR, Jolles J, Riedel WJ: The influence of soy-derived phosphatidylserine on cognition in age-associated memory impairment. Nutr Neurosci 2001;4:121-134.[↩]
- GRAS Notice 636: Phosphatidylserine derived from Soy. https://www.fda.gov/downloads/food/ingredientspackaginglabeling/gras/noticeinventory/ucm505227.pdf[↩]
- ATPase activity of the sodium pump needs phosphatidylserine. Wheeler KP, Whittam R. Nature. 1970 Jan 31; 225(5231):449-50.[↩]
- Roles of ionic residues of the C1 domain in protein kinase C-alpha activation and the origin of phosphatidylserine specificity. Bittova L, Stahelin RV, Cho W. J Biol Chem. 2001 Feb 9; 276(6):4218-26.[↩]
- Phosphatidylserine and phosphatidylethanolamine in mammalian cells: two metabolically related aminophospholipids. Vance JE. J Lipid Res. 2008 Jul; 49(7):1377-87.[↩]
- A review of phosphatidylserine pharmacological and clinical effects. Is phosphatidylserine a drug for the ageing brain? Pepeu G, Pepeu IM, Amaducci L. Pharmacol Res. 1996 Feb; 33(2):73-80.[↩]
- n-3 phosphatidylserine attenuated scopolamine-induced amnesia in middle-aged rats. Vaisman N, Pelled D. Prog Neuropsychopharmacol Biol Psychiatry. 2009 Aug 31; 33(6):952-9.[↩]
- Thematic Review Series: Glycerolipids. Phosphatidylserine and phosphatidylethanolamine in mammalian cells: two metabolically related aminophospholipids. The Journal of Lipid Research July 2008, 49, 1377-1387. http://www.jlr.org/content/49/7/1377.long[↩]
- Akbar M, Calderon F, Wen Z, Kim HY. Docosahexaenoic acid: a positive modulator of Akt signaling in neuronal survival. Proc Natl Acad Sci USA. 2005;102:10858–10863.[↩]
- Kim H-Y, Huang BX, Spector AA. Phosphatidylserine in the Brain: Metabolism and Function. Progress in lipid research. 2014;0:1-18. doi:10.1016/j.plipres.2014.06.002. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4258547/[↩][↩][↩]
- Fadok, V.A.; de Cathelineau, A.; Daleke, D.L.; Henson, P.M.; Bratton, D.L. Loss of phospholipid asymmetry and surface exposure of phosphatidylserine is required for phagocytosis of apoptotic cells by macrophages and fibroblasts. J. Biol. Chem. 2001, 276, 1071-1077.[↩]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112-124.[↩]
- Daleke, D.L. Phospholipid flippases. J. Biol. Chem. 2007, 282, 821-825.[↩]
- Zwaal, R.F.; Comfurius, P.; Bevers, E.M. Lipid-protein interactions in blood coagulation. Biochim. Biophys. Acta 1998, 1376, 433-453.[↩]
- Cho, W.; Stahelin, R.V. Membrane binding and subcellular targeting of C2 domains. Biochim. Biophys. Acta 2006, 1761, 838-849.[↩]
- Arikketh, D.; Nelson, R.; Vance, J.E. Defining the importance of phosphatidylserine synthase-1 (PSS1): Unexpected viability of PSS1-deficient mice. J. Biol. Chem. 2008, 283, 12888-12897.[↩]
- Atkinson, K.; Fogel, S.; Henry, S.A. Yeast mutant defective in phosphatidylserine synthesis. J. Biol. Chem. 1980, 255, 6653-6661.[↩]
- Birge RB, Boeltz S, Kumar S, et al. Phosphatidylserine is a global immunosuppressive signal in efferocytosis, infectious disease, and cancer. Cell Death and Differentiation. 2016;23(6):962-978. doi:10.1038/cdd.2016.11. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4987730/[↩]
- Exposure of platelet membrane phosphatidylserine regulates blood coagulation. Prog Lipid Res. 2003 Sep;42(5):423-38. https://www.ncbi.nlm.nih.gov/pubmed/12814644[↩][↩]
- Zwaal FA, Comfurius P, Bevers EM. Scott syndrome, a bleeding disorder caused by defective scrambling of membrane phospholipids. Biochem Bioph Acta 2004; 1636:119-128[↩]
- Weiss HJ. Scott syndrome: a disorder of platelet coagulant activity (PCA). Sem Hemat 1994; 31:312-319[↩]
- Ethanol promotes neuronal apoptosis by inhibiting phosphatidylserine accumulation. Akbar M, Baick J, Calderon F, Wen Z, Kim HY. J Neurosci Res. 2006 Feb 15; 83(3):432-40.[↩]
- Inhibition of phosphatidylserine biosynthesis in developing rat brain by maternal exposure to ethanol. Wen Z, Kim HY. J Neurosci Res. 2007 May 15; 85(7):1568-78.[↩]
- Effects of ethanol on the remodeling of neutral lipids and phospholipids in brain mitochondria and microsomes. Carrasco MP, Jiménez-López JM, Segovia JL, Marco C. Neurochem Int. 2007 May; 50(6):858-65.[↩]
- Souci SW, Fachmann E, Kraut H (2008). Food Composition and Nutrition Tables. Medpharm Scientific Publishers Stuttgart.[↩]
- Vance JE. Phosphatidylserine and phosphatidylethanolamine in mammalian cells: two metabolically related aminophospholipids. J Lipid Res. 2008;49:1377–1387.[↩]
- Phospholipid synthesis in a membrane fraction associated with mitochondria. Vance JE. J Biol Chem. 1990 May 5; 265(13):7248-56.[↩]
- Structure and expression of the murine phosphatidylserine synthase-1 gene. Sturbois-Balcerzak B, Stone SJ, Sreenivas A, Vance JE. J Biol Chem. 2001 Mar 16; 276(11):8205-12.[↩]
- Synthesis of phosphatidylserine by base exchange in Triton-insoluble floating fractions from rat cerebellum. Buratta S, Felicetti M, Mozzi R. J Neurochem. 2007 Nov; 103(3):942-51.[↩]
- Modulation of phosphatidylserine synthesis by a muscarinic receptor occupancy in human neuroblastoma cell line LA-N-1. Mikhaevitch IS, Singh IN, Sorrentino G, Massarelli R, Kanfer JN. Biochem J. 1994 Apr 15; 299 ( Pt 2)():375-80.[↩]
- Evidence that the major membrane lipids, except cholesterol, are made in axons of cultured rat sympathetic neurons. Vance JE, Pan D, Campenot RB, Bussière M, Vance DE. J Neurochem. 1994 Jan; 62(1):329-37.[↩]
- Distribution and fatty acid composition of phosphoglycerides in normal human brain. Svennerholm L. J Lipid Res. 1968 Sep; 9(5):570-9.[↩][↩]
- Quantification and fatty acid and fatty aldehyde composition of ethanolamine, choline, and serine glycerophosphatides in human cerebral grey and white matter. O’Brien JS, Fillerup DL, Mead JF. J Lipid Res. 1964 Jul; 5(3):329-38.[↩]
- Substrate preference in phosphatidylserine biosynthesis for docosahexaenoic acid containing species. Kim HY, Bigelow J, Kevala JH. Biochemistry. 2004 Feb 3; 43(4):1030-6.[↩][↩]
- n-3 fatty acid deficiency decreases phosphatidylserine accumulation selectively in neuronal tissues. Hamilton L, Greiner R, Salem N Jr, Kim HY. Lipids. 2000 Aug; 35(8):863-9.[↩]
- Phospholipid profile in the postmortem hippocampus of patients with schizophrenia and bipolar disorder: no changes in docosahexaenoic acid species. Hamazaki K, Choi KH, Kim HY. J Psychiatr Res. 2010 Aug; 44(11):688-93.[↩]
- Petursdottir AL, Farr SA, Morley JE, Banks WA, Skuladottir GV. Lipid peroxidation in brain during aging in the senescence-accelerated mouse (SAM) Neurobiol Aging. 2007;28:1170–1178.[↩]
- Cunnane SC, Schneider JA, Tangney C, Tremblay-Mercier J, Fortier M, Bennett DA, et al. Plasma and brain fatty acid profiles in mild cognitive impairment and Alzheimer’s disease. J Alzheimer’s Dis. 2012;29:691–697 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3409580/[↩]
- Ullman L, Mimouni V, Roxx S, Porsolt R, Poisson JP. Brain and hippocampus fatty acid composition in the phospholipid classes of age-related cognitive deficit rats. Prostagl Leukot Essent Fatty Acid. 2001;64:189–195. https://www.ncbi.nlm.nih.gov/pubmed/11334555[↩]
- Corwin J, Dean RL, III, Bartus RT, Rotrosen J, Watkins DL. Behavioral effects of phosphatidylserine in the aged Fischer 344 rats: Amelioration of passive avoidance deficits without changes in psychomotor task performance. Neurobiol Aging. 1985;6:11–15.[↩]
- Lee B, Sur BJ, Han JJ, Shim I, Her S, Lee HJ, et al. Krill phosphatidylserine improves learning and memory in Morris water maze in aged rats. Prog Neuropsychopharmacol Biol Psych. 2010;34:1085–1093.[↩]
- Park HJ, Shim HS, Kim KS, Han JJ, Kim JS, Yu AR, et al. Enhanced learning and memory of normal young rats by repeated oral administration of Krill phosphatidylserine. Nutr Neurosci. 2012;16:47–53.[↩][↩]
- Delwaide PJ, Gyselynck-Mambourg AM, Hurlet A, Ylieff M. Double blind randomized controlled study of phosphatidylserine in senile demented patients. Acta Neurol Scand. 1986;73:136–140.[↩][↩]
- Baumeister J, Barthel T, Geiss KR, Weiss M. Influence of phosphatidylserine on cognitive performance and cortical activity after induced stress. Nutr Neurosci. 2008;11:103–110.[↩]
- Jorissen BL, Brorens F, Van Boxtel MPJ, Riedel WJ. Safety of soy-derived phosphatidylserine in elderly people. Nutr Neurosci. 2002;5:337–343.[↩]
- Richter Y, Herzog Y, Cohen T, Steinhart Y. The effect of phosphatidylserine-containin omega-3 fatty acids on memory abilities in subjects with subjective memory complaints; a pilot study. Clin Intervent Aging. 2010;5:313–316.[↩]
- Vakhapova V, Cohen T, Richter Y, Herzog Y, Korczyn AD. Phosphatidylserine containing ω-3 fatty acids may improve memory abilities in non-demented elderly with memory complaints: A double-blind placebo-controlled trial. Dement Geriatr Cogn Disord. 2010;29:467–474.[↩]
- Kato-Kataoka A, Sakai M, Ebina R, Nonaka C. Soybean derived phosphatidylserine improves memory function of the elderly Japanese subjects with memory complaints. J Clin Biochem Nutr. 2010;47:246–255 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2966935/[↩]
- Leon A, Benvegnú D, Toffano G, Orlando P, Massari P. Effect of brain cortex phospholipids on adenylate-cyclase activity in mouse brain. J Neurochem. 1978;30:23–26.[↩]
- Casamenti F, Mantovani P, Amaducci L, Papeu G. Effect of phosphatidylserine on acetylcholine output from the cerebral cortex of rats. J Neurochem. 1979;32:529–533.[↩]
- Casamenti F, Scali C, Papeu G. Phosphatidylserine reverses the age-dependent decrease in cortical acetylcholine release: a microdialysis study. Eur J Pharmacol. 1991;194:11–16.[↩]
- Kim HY. Novel metabolism of docosahexaenoic acid in neural cells. J Biol Chem. 2007;282:18661–18665.[↩]
- Hellhammer J, Vogt D, Franz N, Freitas U, Rutenberg D. A soy-based phosphatidylserine/phosphatidic acid complex (PAS) normalizes the stress reactivity of hypothalamus-pituitary-adrenal-axis in chronically stressed male subjects: a randomized, placebo-controlled study. Lipids Health Dis. 2014;13:121. 11 pages.[↩]
- Heywood R, Cozens DD, Richold M. Toxicology of a phosphatidylserine preparation from bovine brain (BC-PS). Clin Trials J. 1987;24:25-32.[↩]

{kind=link}