Contents
What is leaky gut
The medical name for leaky gut is “intestinal permeability” and it’s neither disease nor symptom—intestinal permeability is a natural biological function. Lining your intestines is a barrier of cells; between each cell is a tight junction that keeps bad things (like toxins and bad bacteria) out of your body and allows good things (nutrients) in. The issue is that this natural permeability can go awry if the barrier becomes faulty. In genetically susceptible people, leaky gut can be triggered by gut bacteria disruptions from poor diet, some medications or gluten. Importantly, experts don’t agree on what is and isn’t a trigger. But once permeability is disrupted, the bad guys “leak” into your bloodstream and trigger inflammation, causing other health issues.
For digestion and absorption purposes, humans have developed a very complicated and highly specialized gastrointestinal system maintained by the mucosal barrier 1). The intestinal epithelium is the largest mucosal surface providing an interface between the external environment and the mammalian host. The intestinal mucosa faces tremendous exterior antigens, including food antigens, commensal bacteria, pathogens, and toxins. Its exquisite anatomical and functional arrangements and the finely-tuned coordination of digestive, absorptive, motility, neuroendocrine, and immunological functions are testimonial of the complexity of the gastrointestinal (GI) system. Also pivotal is the regulation of molecular trafficking between the intestinal lumen and the submucosa via the paracellular space. Thus, a specialized barrier function is required to block the entry of diverse exterior antigens while absorbing nutrients.
Impressively, in the intestine, the front line of this barrier is maintained by only a single layer of specialized epithelial cells that are linked together by tight junction (TJ) proteins. Many other factors aid in support of this barrier including mucins, antimicrobial molecules, immunoglobulins, and cytokines. If any abnormalities occur among these factors, the intestinal permeability may increase, which is termed a “leaky gut” 2). A leaky gut allows the entry of exterior antigens from the gut lumen into the host, which may promote both local and systemic immune responses. Multiple diseases may arise or be exacerbated due to a leaky gut, including autoimmune diseases such as inflammatory bowel disease, celiac disease, autoimmune hepatitis, type 1 diabetes, multiple sclerosis (MS) and systemic lupus erythematosus (SLE) 3), 4), 5), 6), 7). Numerous factors can affect gut permeability, such as various diet-derived compounds, alcohol consumption, and gut microbiota dysbiosis.
The Gut Barrier
A large variety of exogenous substances colonize the gut lumen, such as microorganisms, toxins, and antigens. Without an intact and properly functioning intestinal barrier, these substances can penetrate the tissues beneath the intestinal epithelial lining, diffuse into blood and lymphatic circulations, and disrupt tissue homeostasis. However, there is an efficient multifaceted intestinal barrier system with physical, biochemical, and immunological components that prevents the entry of most pathogens (Figure 1). These components coordinate with each other to prevent uncontrolled translocation of luminal contents into the body. Below is a brief synopsis of the main components comprising the intestinal barrier.
Figure 1. Illustration of host intestinal barriers
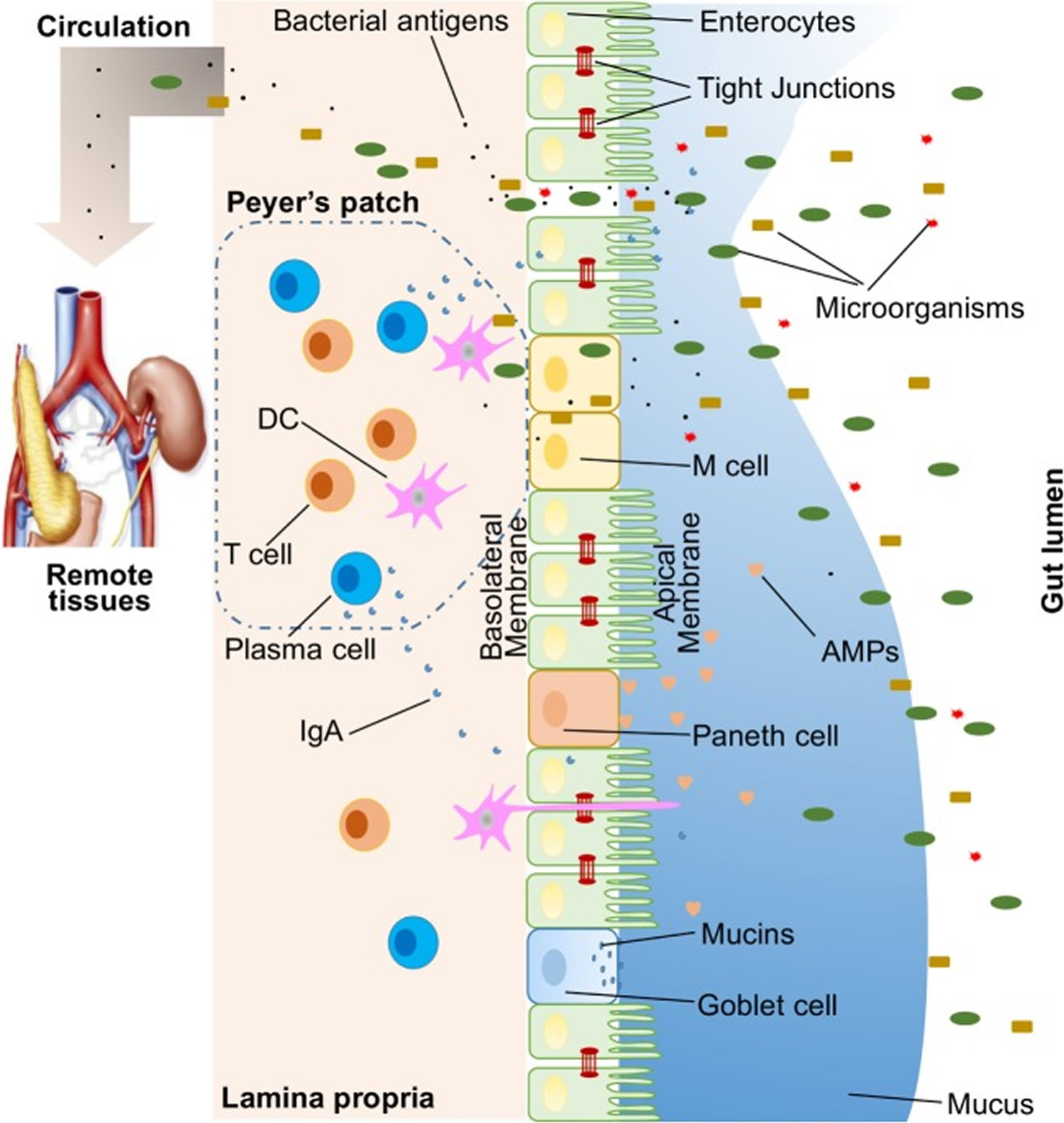
Note: Illustration of host intestinal barriers, including physical barrier (epithelium, tight junctions, mucus, commensal bacteria), biomedical barrier [antimicrobial proteins (AMPs)], and immunological barrier (lymphocytes and IgA). Also shown is the microbial translocation to remote tissues (for example, kidney and pancreas) in the presence of a leaky gut.
[Source 8)]Physical Barrier
In humans, the intestinal epithelium covers as large as 400 m2 of surface area 9). Though only a single layer of cells, the intestinal epithelial cells (IECs) are the mainstay of the intestinal barrier and serve as a physical barrier (Figure 1). There are at least seven types of functional intestinal epithelial cells (IECs)—enterocytes, goblet cells, Paneth cells, microfold cells (M cells), enteroendocrine cells, cup cells, and tuft cells, although the functions of the last two cell populations are not well understood 10). Among all these cell types, enterocytes represent the absolute majority, accounting for at least 90% of crypt cells or villus cells. Enterocytes are absorptive cells and vital for nutrient uptake. However, growing evidence indicates that the functions of enterocytes are not limited to nutrient absorption. For example, enterocytes can control the abundance of Gram-positive bacteria by expressing RegIIIγ, one type of antimicrobial proteins (AMPs) 11). All epithelial cell types originate from Lgr5+ intestinal epithelial stem cells, which reside within the crypts 12). The turnover rate of IECs is high and the cells are renewed every 3–5 days in the mammalian intestine 13), with the exception being the Paneth cells, which have a life span of about 2 months.
The intestinal epithelial cells (IECs) lining is continuous, and the contact between IECs is sealed by tight junctions (TJs) 14). The paracellular pathway, in contrast to transcellular pathway, allows the transport of substances across the gut epithelium through the spaces between intestinal epithelial cells (IECs). A large variety of molecules, mainly proteins, control the plasticity of tight junctions (TJs). More than 40 tight junction (TJ) proteins have been recognized, including occludin, claudins, junctional adhesion molecule A, and tricellulin (17). Under various pathological conditions, paracellular permeability may be increased, resulting in the entry of unwelcome, potentially harmful molecules.
On top of the gut epithelium, there are two layers of mucus, the inner and outer layers, that cover the whole intestinal epithelial lining and provide physical protection to separate luminal microorganisms from the epithelium. Organized by its major component, a highly glycosylated gel-forming mucin MUC2, the mucus contains diverse molecules including IgA as well as enzymes and proteins, such as lactoferrin 15). Goblet cells are the central cell type for the formation of mucus. They not only produce MUC2 mucin but also secret other mucus components such as ZG16, AGR2, FCGBP, CLCA1, and TFF3 16). Colitis would spontaneously develop in Muc2-deficient mice, indicating a critical role for MUC2 in mucosal protection 17). In addition to gel-forming mucins, there is another type of mucin that is in close proximity to epithelial cells, called transmembrane mucins. Enterocytes are the main producers of transmembrane mucins 18).
The gut commensal bacteria have been described as one component of the intestinal physical barrier primarily due to its two major functions 19). The first is to promote resistance to the colonization of harmful or pathogenic bacteria species by competing for nutrients, occupying attachment sites, and releasing antimicrobial substances 20). Additionally, the gut microbiota regulates the digestion and absorption of nutrients to supply energy to epithelial cells, which are a major component of the physical barrier 21). A good example of the direct energy supply is the production of short-chain fatty acids by the gut microbiota, which are used by colonocytes for their development and metabolism 22). Taken together, IECs, the mucus layers, and gut microbial residents serve as the physical barrier to limit the entry of unfriendly luminal contents into host tissues.
Biochemical Barrier
Biochemical molecules with antimicrobial properties exist in the mucus as well as far into the lumen and include bile acids and AMPs 23) (Figure 1). These diverse molecules form a complicated network to reduce the load of colonized bacteria and decrease the chance of contact between luminal antigens and host cells. They are a good supplement to the physical barrier and an essential component of the intestinal barrier function.
The proximal small intestine harbors very few microorganisms 24). But as the distance from the stomach increases, the pH rises and the number of colonized bacteria esculates 25). Facing a large number of microorganisms, which likely outnumber the number of host cells, multiple AMPs are generated to fight against invaders. These AMPs are divided into several types, including α- and β-defensins, C-type lectin, cathelicidin, lysozyme, and intestinal alkaline phosphatase (IAP) 26). Their detailed antimicrobial mechanisms are discussed elsewhere 27). As a major, but not exclusive, producer of AMPs, Paneth cells support and mediate the biochemical barrier function.
Immunological Barrier
Below the intestinal epithelium, there are organized lymphoid follicles, including the Peyer’s patches and isolated lymphoid follicles. Inside the follicles, a variety of immune cells, including B cells, T cells, dendritic cells (DCs), and neutrophils, orchestrate the immune response by presenting antigens, secreting cytokines, and producing antigen-binding antibodies (Figure 1). In the intestinal epithelium where lymphoid follicles are found, M cells are present that transcytose antigens across the intestinal epithelium to the Peyer’s patches underneath 28). In addition, goblet cells present acquired luminal antigens to CD103+ dendritic cells (DCs) in lamina propria in small intestine by forming goblet cell-associated antigen passages (GAPs) 29). Interestingly, spontaneous antigen presentation was also observed in the colon, but only when the mice were raised germ-free (GF), or housed conventionally but with oral antibiotic treatment 30). This suggests that the antigen uptake process and formation of GAPs are regulated by the colonic microbiota 31). In addition, goblet cells and GAPs are capable of sensing invasive pathogens and inhibiting the translocation of pathogenic bacteria into the host immune system 32). Furthermore, intestinal mononuclear phagocytescan sense and sample luminal contents 33). CX3CR1-expressing cells are responsible for this process, and antigen sampling is dependent on structures called transepithelial dendrites (TEDs) 34). The formation of TEDs is regulated by CX3CR1+ macrophages and the expression of CX3CL1 by certain IECs 35).
Another component of the immunological barrier is secretory IgA (SIgA). As the most abundant immunoglobulin in the body, IgA resides primarily on intestinal mucosal surfaces. While some people with selective IgA deficiency appear to be healthy, SIgA is important as it presumably interacts with commensal bacteria to provide protection against pathogens. A unique feature about SIgA is that is structurally resilient in protease-rich environments allowing it to remain functionally active compared to other antibody isotypes on mucosal surfaces 36). In adult humans, about 50 mg/kg of SIgA is produced daily by plasma cells residing in the intestinal lamina propria. Finally, SIgA can be transcytosed through the epithelium and secreted into the gut lumen.
Though not mentioned here, self-modulating factors, such as nerves and diverse cytokines, are also important for maintaining the normal functions of the intestinal barrier.
Mechanisms of Leaky Gut
A large variety of gut barrier disruptors and/or gut microbiota disturbers may potentially result in microbial translocation and subsequent inflammation locally and systemically. These include diet, infections, alcohol consumption, and burn injury.
Diet-Induced Gut Leakiness
Nutrients and food ingredients have been reported to contribute to the maintenance or alterations of gut microbiota and the intestinal barrier function 37). A recent review by De Santis et al. detailed many dietary factors that may modulate the intestinal barrier 38). Some recent publications emphasize the effects of diet-induced alterations of gut microbiota on compromising the gut barrier function. Vitamin D has been recognized as an intestinal permeability protector by inducing the expression of TJ proteins ZO-1 and claudin-1. In vitamin D receptor (VDR)-knockout mice, more severe experimental colitis has been observed, suggesting the protective effect of vitamin D on the mucosal barrier 39). However, another group have recently found that vitamin D receptor deficiency lowers, whereas vitamin D treatment upregulates, the expression of claudin-2, a pore-forming TJ protein, which renders the intestinal epithelium leaky 40). Further analysis confirmed that vitamin D receptor enhanced claudin-2 promoter activity. The exact role of vitamin D and vitamin D receptor on modulating intestinal permeability is therefore unclear and should be investigated carefully in association with gut microbiota. In a recent study by Desai et al., a low-fiber diet consumption was found to trigger the expansion of mucus-degrading bacteria, including Akkermansia muciniphila and Bacteroides caccae 41). As a result, the thickness of mucus is significantly decreased in mice fed with fiber-deficient diets, although the transcription of Muc2 gene was surprisingly heightened, possibly as a compensatory response. The thinner mucus and compromised intestinal barrier function lead to a higher susceptibility to certain colitis-causing pathogens 42). Moreover, a diet high in saturated fat has been shown to greatly decrease Lactobacillus and increase Oscillibacter, and these changes were correlated with significantly increased permeability in the proximal colon 43). Furthermore, studies revealed that the abundance of the Oscillospira genus was negatively correlated with the mRNA expression of barrier-forming TJ protein ZO-1.
Stress-Induced Gut Leakiness
Under certain circumstances, stress-induced alterations of gut microbiota and the impaired intestinal barrier would allow the occurrence of microbial translocation. Burn injury and alcohol consumption are examples of such stress. Burn injury results in increased intestinal permeability, which is mediated by increased activity of myosin light-chain (MLC) kinase 44). It is known that MLC phosphorylation or kinase activation can trigger epithelial TJ opening 45). In burn injury, TJ proteins, including ZO-1, occluding, and claudin-1, are redistributed, which can be reversed by adding an MLC phosphorylation inhibitor. In addition, both humans and mice experiencing burn injury undergo similar alterations of gut microbiota, in particular, with increases of the abundance of bacteria from the Enterobacteriaceae family 46). Importantly, microbial translocation of these Gram-negative aerobic bacteria has been observed. Another research group, using a different burn injury mouse model reported increased colonic permeability together with reduced aerobic and anaerobic bacterial populations in the gut microbiota, particularly those producing butyrate 47). As a consequence, the butyrate level in the stool was significantly decreased in mice with burn injury. Interestingly, when the experimental mice received fecal microbiota transplant, their altered bacterial counts and impaired mucosal barrier function were reversed, suggesting direct involvement of microbiota in causing gut leakiness after burn injury.
Chronic alcohol consumption is responsible for intestinal barrier dysfunction, alterations on both the quality and quantity of gut microbiota, LPS translocation, and alcoholic liver disease (ALD). In both human and mouse, it has been well established that alcohol can disrupt intestinal barrier function, which is closely related to increased tumor necrosis factor (TNF) production from intestinal monocytes/macrophages and enterocytes bearing TNF-receptor 1, followed by downstream activation of MLC kinase 48). Notably, when mice given chronic alcohol also received oral antibiotic treatment, to remove the microbiota, the level of TNF production and intestinal permeability decreased to levels comparable to those in control mice 49). This indicates that the alcohol-induced, TNF-mediated gut leakiness is greatly dependent on gut microbiota. Indeed, though the mechanism is unknown, alcohol administration alters microbiota qualitatively and quantitatively in both human and mouse 50). Bacterial overgrowth has been observed with alcohol consumption, whereas antibiotics can decrease the bacterial load and attenuate ALD 51). Interestingly, probiotic Lactobacillus is significantly suppressed during alcohol consumption 52). Directly supplying Lactobacillus strains or indirect stimulation of Lactobacilli with prebiotics or diets can decrease bacterial overgrowth, restore mucosal integrity of the intestine, and suppress microbial translocation 53). Microbial translocation, especially the translocation of LPS, is involved in ALD development and progression as evidenced by the lack of ALD in mice deficient of TLR4 54). It is worth noting that some bacteria species can produce alcohol, including E. coli and Weissella confusa, and this may be the mechanism by which they compromise the intestinal barrier function 55).
Infections can play a role in regulating the mucosal barrier. A good example is Helicobacter pylori, a Gram-negative bacterium infecting the human stomach 56). H. pylori is known to directly increase epithelial permeability by redistributing TJ protein ZO-1 57). In addition, bacteriophages, which are usually not considered pathogenic to mammals, can have an impact on the leaky gut. When rats were given a bacteriophage cocktail containing phages against Salmonella enterica, disruption of the intestinal barrier integrity was observed 58). The authors speculated that the gut microbiota might have been affected by bacteriophages, but sequencing data were not supplied to support their claims.
Taken together, perturbation of gut microbiota, which may be the consequence of diverse interventions, can lead to increased intestinal permeability and translocation of bacterial components and products. Such microbial translocation can subsequently trigger an abnormal immune response, causing inflammation and/or tissue damage in extraintestinal organs.
Leaky Gut Symptoms
There’s strong evidence that leaky gut can cause food allergies, inflammatory bowel disease (IBD) and celiac disease. There’s good evidence leaky gut may lead to type 1 diabetes or multiple sclerosis, and more limited research links it to nonalcoholic fatty liver disease, nonalcoholic steatohepatitis (NASH) and type 2 diabetes. Other experts believe leaky gut contributes to a wider host of ills, such as seasonal allergies, depression and eczema.
How to heal leaky gut
Considering the contributions of leaky gut and bacterial translocation to inflammation and multiple diseases, reversing gut leakiness appears to be an attractive therapeutic strategy. Prebiotics and probiotics, for example, can be used to reduce intestinal permeability 59). Diverse probiotic species have been uncovered that possess the properties to protect the intestinal barrier through targeting different components of the mucosal barrier system. The human commensal Bacteroides fragilis may serve as such a probiotic 60). In a mouse model, autism spectrum disorder (ASD) has been shown to be accompanied by intestinal barrier dysfunction, gut microbiota dysbiosis, and leakiness of 4-ethylphenylsulfate (4EPS), which originates from the commensal bacteria. When 4EPS was given to wild-type mice, it directly caused behavioral abnormalities similar to ASD mice. Treatment with B. fragilis reduced the translocation of disease-causative 4EPS, and significantly ameliorated the behavior defects. The therapeutic benefit of B. fragilis is believed to be due to its ability to alter microbial composition and enhance intestinal barrier function 61). B. fragilis is also known for its capability to induce the development of Foxp3+ regulatory T cells, a process regulated by another product of B. fragilis, polysaccharide A 62). B. fragilis and polysaccharide A are beneficial against inflammatory diseases, such as colitis and experimental autoimmune encephalomyelitis 63). The application of B. fragilis to prevent the leaky gut and reverse autoimmunity warrants further investigation.
Lactic acid bacteria and Bifidobacteria are known to secrete factors that dampen inflammation by downregulating NF-κB dependent gene expression, IL-8 secretion, and levels of macrophage-attracting chemokines 64). Lactic acid bacteria and Bifidobacteria have also been shown to directly downregulate T effector-mediated inflammatory responses while upregulating anti-inflammatory T regulatory cell expression in mice 65). The exact mechanism of how these gut flora modulate immune responses is still not well understood; in a practical point of view, probiotic candidates with different targets on reversing the leaky gut may synergistically act to attenuate disease as thus may serve as a probiotic cocktail. As probiotics are generally considered safe, it is anticipated that they will become cost-effective treatment options for people with autoimmune diseases in the foreseeable future. This is a very young but exciting field in which much still remains to be learned.
References [ + ]



{kind=link}