Contents
- What is Serotonin
- Serotonin Synthesis and Metabolism
- The synthesis of serotonin can increase markedly under conditions requiring a continuous supply of the neurotransmitter
- As with other biogenic amine transmitters, serotonin is stored primarily in vesicles and released by an exocytotic mechanism
- The activity of serotonin in the synapse is terminated primarily by its re-uptake into serotonergic terminals
- Serotonin breakdown (metabolism)
- Serotonin function
- Serotonin and depression
- How to increase serotonin
- Serotonin foods
- Selective Serotonin Reuptake Inhibitors (SSRIs)
- Serotonin syndrome
- What does serotonin syndrome feel like?
- How long does serotonin syndrome last?
- Serotonin syndrome long term effects
- Serotonin syndrome causes
- Risk factors for serotonin syndrome
- Serotonin syndrome prevention
- Serotonin syndrome signs and symptoms
- Serotonin syndrome complications
- Serotonin syndrome diagnosis
- Serotonin syndrome treatment
- Serotonin syndrome prognosis
- Serotonin Synthesis and Metabolism
What is Serotonin
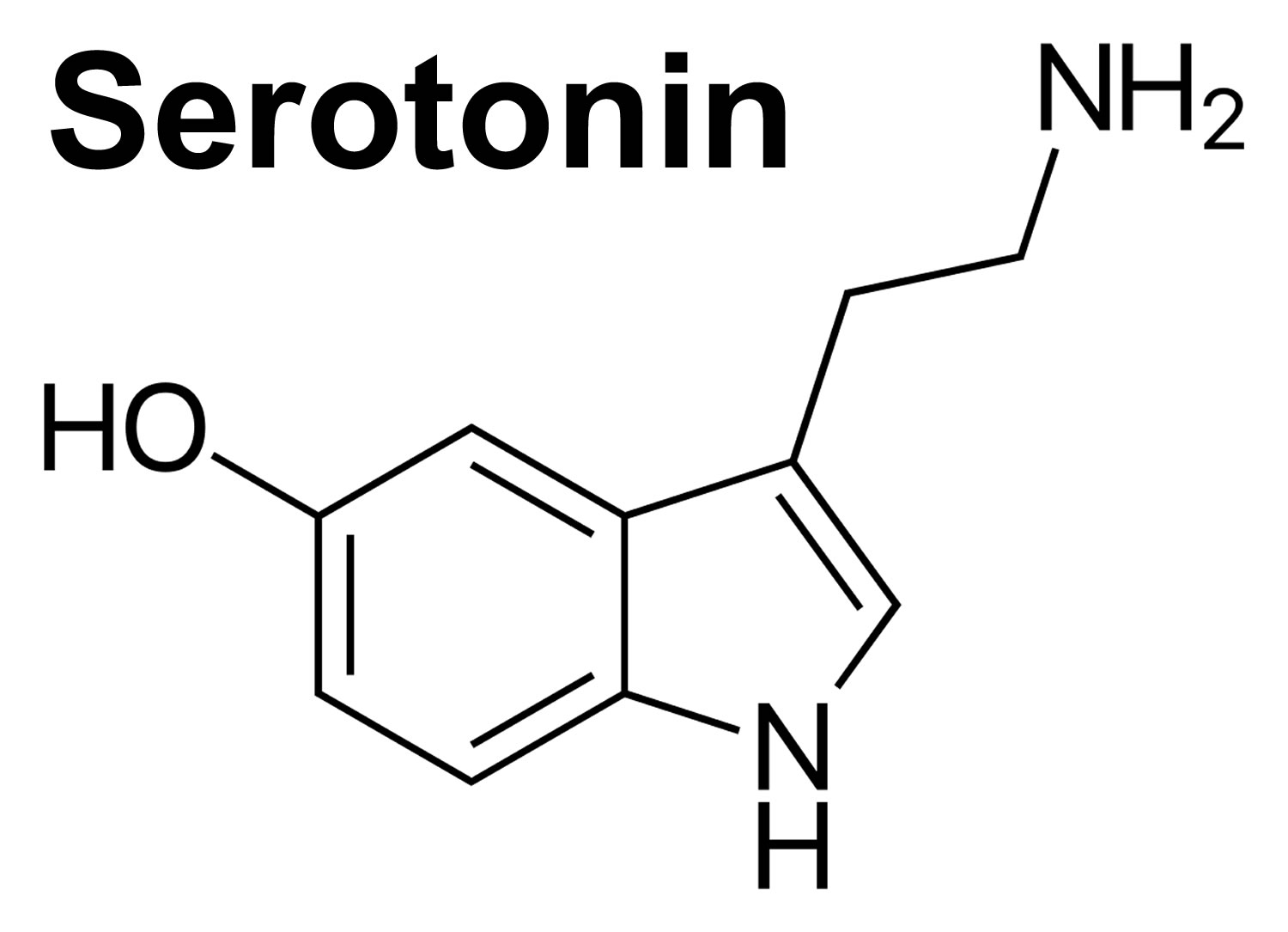
Serotonin also known as 5-hydroxytryptamine or 5-HT, is one of the a biochemical messengers and regulators, synthesized from the essential amino acid L-Tryptophan that nerve cells in the brain use to communicate 1, 2, 3. Serotonin forms from the hydroxylation (i.e., the addition of -OH [hydroxyl] group) and decarboxylation of the tryptophan amino acid. The highest serotonin concentration is in the gastrointestinal tract’s enterochromaffin cells, with small amounts in the central nervous system (brain and spinal cord) and platelets 1. Serotonin induces changes in the cell by its action on the serotonergic receptors, which are coupled to different G proteins mediating intracellular changes 2. Serotonin activates the intracellular signaling cascade through its 15 receptors, which are classified into 7 families (HTR1 to HTR7) based on structural, pharmacological, and signal transduction properties 4. With the exception of HTR3, a ligand-gated cation channel, all other subtypes are G protein-coupled receptors that activate various intracellular signaling pathways 5, 6. Serotonin receptors are widely distributed in the brain and various peripheral organs. These observations suggest that serotonin is not only an important neurotransmitter in the central nervous system (brain and spinal cord), but also may exert its effects through its receptors specifically expressed in different peripheral tissues 7. During development, several serotonin receptor subtypes emerge in the developing brain prior to enzymes for serotonin synthesis, suggesting an extra-embryonic source of serotonin in the early stages of neurodevelopment 8.
Serotonin can also act in a receptor-independent manner by covalently binding to glutamine residues of various extracellular, cytoplasmic or nuclear proteins, in a process known as serotonylation 9. Serotonylation has been implicated in the regulation of platelet functions 10, 11, insulin secretion 12 and gene expression regulation 13.
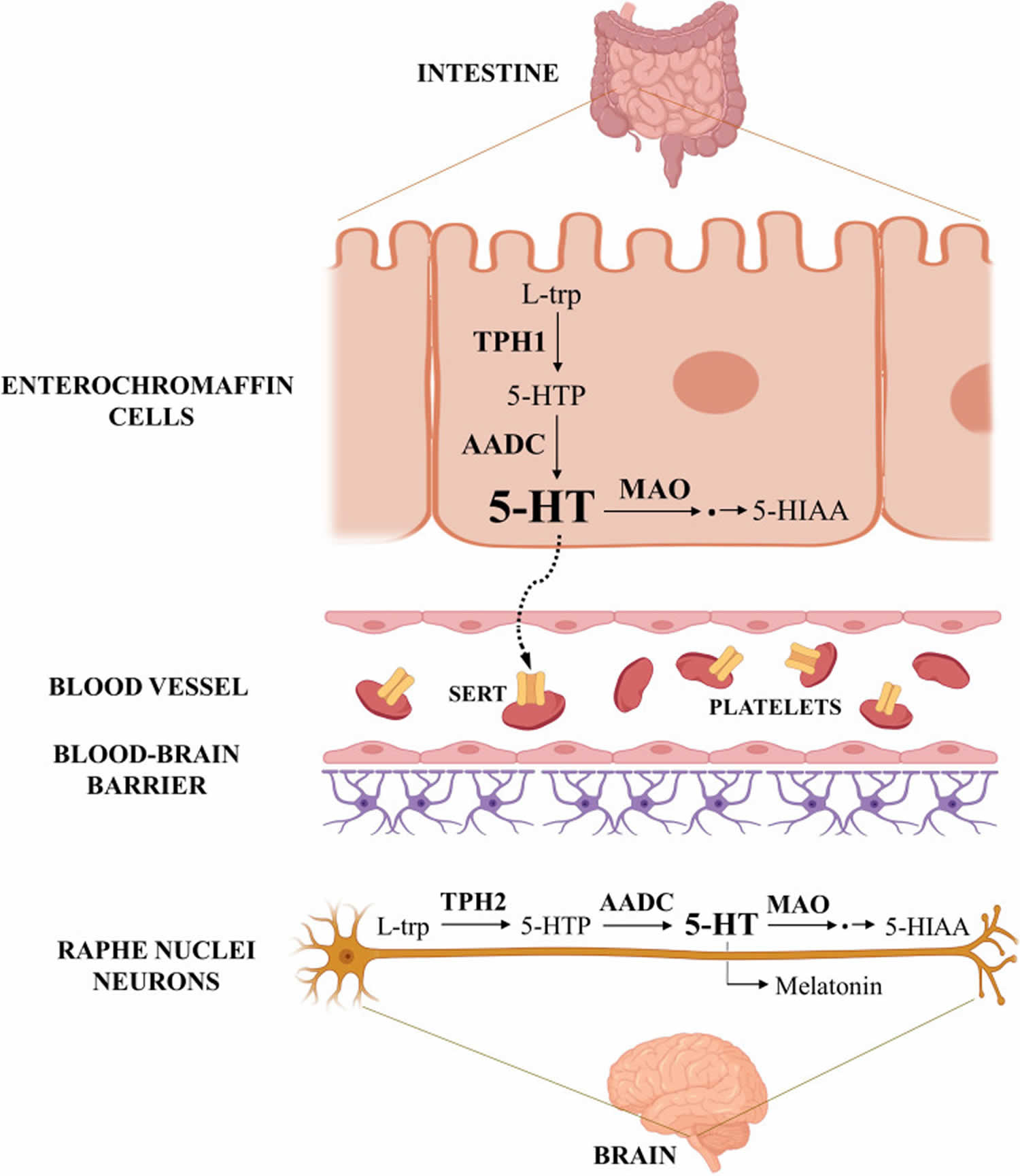
Serotonin (5-hydroxytryptamine [5-HT]) was originally called enteramine, first isolated serendipitously by Vittorio Erspmarer in 1935 during his attempt to purify extract from enterochromaffin cells of the intestine and was shown to induce intestinal contractions 14, 15. In the 1940s, the same substance was identified as a “tone-modifier” present in serum and therefore named serotonin in 1948 16, 17. Only about 1% to 2% of total amount of serotonin is produced by serotonergic neurons in the brain, whereas 90% of serotonin is detected to be secreted from the enterochromaffin cells of gastrointestinal tract (GIT) 18. Some of the intestine-derived serotonin is secreted into the blood, where it is rapidly internalized in platelets and only a tiny fraction (< 1%) remains free in plasma 19. There are other niches of local serotonin synthesis in the human body, including serotonergic neurons, pinealocytes, pulmonary artery endothelial cells, mammary epithelial cells, mast cells, pancreatic beta cells, adipocytes, hepatocytes, osteoclasts, melanocytes, keratinocytes, and fibroblasts 20, 21, 22.
Today, serotonin (5-hydroxytryptamine [5-HT]), a monoamine neurotransmitter, is best known for its neurotransmitter role in the brain (helping transmit nerve impulses), but serotonin acts also as a hormone and autocrine/paracrine messenger in various other organs 23, 24. Serotonin (5-hydroxytryptamine [5-HT]) modulates brain functions such as mood, emotion, cognition, wake-sleep cycle, appetite, sexual behavior, pain perception, and response to stress 25. In addition, serotonin (5-hydroxytryptamine [5-HT]) regulates and fine tunes numerous other physiological processes, including stopping of bleeding (hemostasis) and constrict blood vessels, gastrointestinal functions, immune response, reproductive functions, bone remodeling, and energy balance 23, 26, 7. Moreover, Serotonin contributes to regulating organ development and regeneration, by controlling basic cellular processes such as proliferation, differentiation, and migration 27, 28. In the developing fetal brain, Serotonin acts as a neurotrophic factor for various neuronal populations and also regulates the maturation of its own neurons 29, 30. Less well known are the antioxidant effects of serotonin in scavenging reactive oxygen species (ROS) and inhibiting lipid peroxidation 31.
Serotonin is metabolized by the liver and its metabolites, primarily 5-HIAA (5-hydroxyindoleacetic acid), are eliminated in the urine. In healthy volunteers, the transient reduction of serotonergic neurotransmission by means of dietary depletion of tryptophan has often been associated with mood-lowering 32. Serotonin deficiency causes disorders such as depression, obsessive-compulsive disorder (OCD), phobias, posttraumatic stress disorder (PTSD), epilepsy, and generalized anxiety disorder (GAD). Modifying serotonin levels is one way that antidepressant and anti-anxiety medications are thought to work and help people feel better. selective serotonin reuptake inhibitors, whose mechanism of action is believed to rely on the enhancement of serotonergic function in the brain, are effective antidepressants 33. In vivo measurements of various components of serotonin neurotransmission in patients with major depressive disorder and their relatives all support the theory of an absolute or relative deficit in cortico-limbic pathways believed to regulate the expression of mood 34. Serotonin has three different modes of action in the nervous system: as neurotransmitter, acting locally at synaptic boutons; upon diffusion at a distance from its release sites, producing paracrine (also called volume) effects, and by circulating in the blood stream, producing hormonal effects. The three modes can affect a single neuronal circuit 35. However, the precise nature of serotonin’s role in the brain is largely unknown.
In humans serotonin is found primarily in the central nervous system (brain and spinal cord), gastrointestinal tract’s enterochromaffin cells, special cells in the bronchial tubes (lungs) and blood platelets. More than 90% of serotonin in the blood is found in the platelets. Serotonin mediates several important physiological functions including neurotransmission, gastrointestinal motility, hemostasis, and cardiovascular integrity. Multiple receptor families (serotonin receptors the serotonin receptor 1AR and serotonin receptor 2AR) explain the broad physiological actions and distribution of this biochemical mediator 36. The biological function of serotonin plays several roles in the human body, such as influencing learning, memory, happiness, and reward, as well as physiological processes such as sleep regulation, behavior, and appetite 37. Serotonin also plays a significant role in the pharmacodynamic activities of antidepressants, such as selective serotonin reuptake inhibitors (SSRIs) and serotonin and norepinephrine reuptake inhibitors (SNRIs), which are a type of antidepressant medication that works by increasing serotonin (and norepinephrine) levels in the brain, which is thought to help improve mood and other symptoms of depression and anxiety.
A significantly increased level of serotonin in a person with carcinoid syndrome symptoms is suggestive but not diagnostic of a carcinoid tumor. In order to diagnose the condition, the tumor itself must be located, biopsied, and examined by a pathologist. The healthcare practitioner will frequently follow an abnormal test result with an order for an imaging scan to help locate any tumor(s) that may be present. Someone may still have a carcinoid tumor even if the concentrations of serotonin and 5-HIAA (5-hydroxyindoleacetic acid) are normal. Some carcinoid tumors do not produce serotonin or only produce it intermittently. A person with no symptoms and normal levels of serotonin and 5-HIAA (5-hydroxyindoleacetic acid) is unlikely to have a serotonin-secreting carcinoid tumor.
Serotonin is produced as needed by the nervous system, mainly the brain, but also by carcinoid tumors are slow-growing masses that can form in the gastrointestinal tract (especially in the appendix) and in the lungs, although they may affect other organs as well. They are one of several types of tumors that arise from cells in the neuroendocrine system. These cells, which secrete hormones in response to signals from the nervous system, are found in organs throughout the body. The serotonin produced by carcinoid tumors may cause flushing of the face, diarrhea, a rapid heart rate, and wheezing, especially when the tumor has spread to the liver. This group of signs and symptoms is referred to as the carcinoid syndrome. According to the American Cancer Society, there are about 8,000 gastrointestinal and 4,000 lung carcinoid tumors diagnosed each year in the United States. Many more of these tumors may exist, but most remain small and do not cause any symptoms. When carcinoid tumors are discovered in asymptomatic patients during surgical procedures performed for other reasons, they are called “incidental” tumors. A small percentage of these tumors may eventually grow large enough to cause obstructions in the intestines or bronchial tubes of the lungs.
Serotonin in the nervous system acts as a local transmitter at synapses, and as a paracrine or hormonal modulator of circuits upon diffusion, allowing a wide variety of “state-dependent” behavioral responses to different stimuli 35. Such is the case of aggressive behavior and rhythmic motor patterns , including those responsible for feeding. In vertebrates, which display a wider and much more sophisticated behavioral repertoire, serotonin also modulates sleep, the arousal state, sexual behavior, and others 38.
Serotonin-containing neuronal cell bodies are restricted to discrete clusters or groups of cells located along the midline of the brainstem. Their axons, however, innervate nearly every area of the CNS (Figures 1 and 2). In 1964, Dahlstrom and Fuxe 39, using the Falck-Hillarp technique of histofluorescence, observed that the majority of serotonergic soma are found in cell body groups, which previously had been designated as the raphe nuclei. This earlier description of the raphe nuclei was based on cell body structural characteristics and organization. Dahlstrom and Fuxe described nine groups of serotonin-containing cell bodies, which they designated B1 through B9, and which correspond for the most part with the raphe nuclei 39 (Table 1). Some serotonergic neuronal cell bodies, however, are found outside the raphe nuclei, and not all of the cell bodies in the raphe nuclei are serotonergic. In most of the raphe nuclei, the majority of neurons are nonserotonergic. For example, the dorsal raphe contains the largest number of serotonergic neurons; however, only 40 to 50% of the cell bodies in the dorsal raphe are serotonergic 40.
Figure 1. Serotonergic pathway in the brain
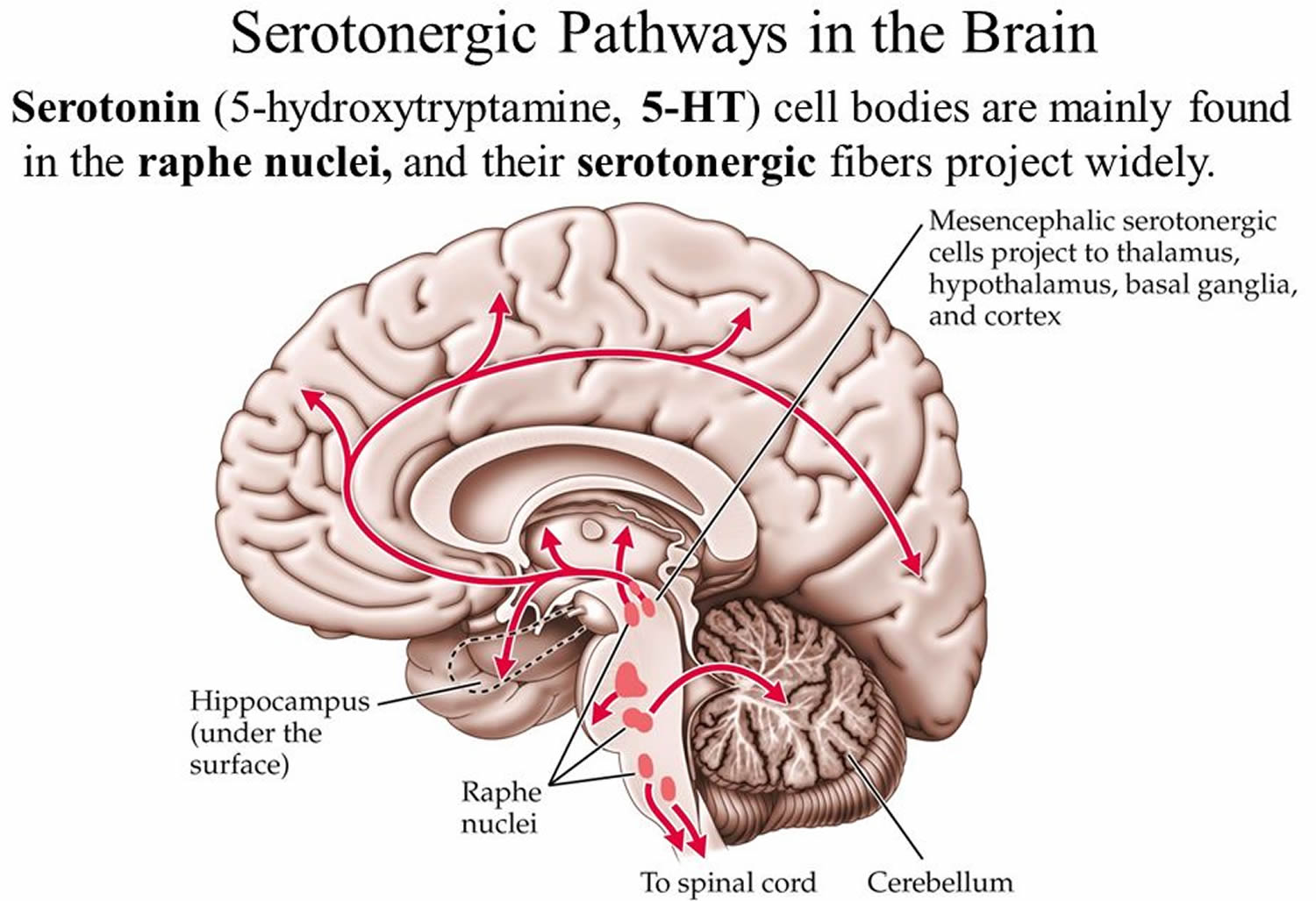
Figure 2. Location of the serotonergic cell body in the brain

Footnotes: Schematic drawing depicting the location of the serotonergic cell body groups in a sagittal section of the rat central nervous system and their major projections.
Abbreviations: OT= olfactory tuberculum; Sept = septum; C. Put = nucleus caudate-putamen; G. Pal = globus pallidus; T = thalamus; H = habenula; S. Nigra = substantia nigra.
[Source 40]Table 1. Classification of Serotonergic Cell Body Groups According to Dahlstrom and Fuxe and Corresponding Anatomical Structure
| Groups of serotonin-containing cell bodies | Anatomical structure |
|---|---|
| B1 | Raphe pallidus nucleus, caudal ventrolateral medulla |
| B2 | Raphe obscurus nucleus |
| B3 | Raphe magnus nucleus, rostral ventrolateral medulla, lateral paragigantocellular reticular nucleus |
| B4 | Raphe obscurus nucleus, dorsolateral part |
| B5 | Median raphe nucleus, caudal part |
| B6 | Dorsal raphe nucleus, caudal part |
| B7 | Dorsal raphe nucleus principal, rostral part |
| B8 | Median raphe nucleus, rostral main part; caudal linear nucleus; nucleus pontis oralis |
| B9 | Nucleus pontis oralis, supralemniscal region |
Serotonin Synthesis and Metabolism
Serotonin (5-hydroxytryptamine [5-HT]) biosynthesis uses the essential amino acid L-tryptophan as the precursor and involves 2 enzymatic steps (Figure 3). The first step, conversion of L-tryptophan to 5-hydroxytryptophan (5-HTP), is catalyzed by rate-limiting tryptophan hydroxylase (TPH) that hydroxylate (the addition of -OH [hydroxyl] group) and subsequent decarboxylation of 5-hydroxytryptophan (5-HTP) to serotonin, is catalyzed by aromatic amino acid decarboxylase (AAAD). Conversion of L-tryptophan to 5-hydroxytryptophan (5-HTP) tryptophan hydroxylase (TPH) is the rate-limiting step in the synthesis of serotonin 41, 42. There are 2 tryptophan hydroxylase (TPH) isoforms, tryptophan hydroxylase 1 (Tph1) and tryptophan hydroxylase 2 (Tph2), encoded by genes located on human chromosome 11 and 12, respectively 43. The two tryptophan hydroxylase (TPH) isoforms have different tissue expression – tryptophan hydroxylase 1 (Tph1) (peripheral) is abundant in enterochromaffin cells of the intestine and in other peripheral tissues, whereas tryptophan hydroxylase 2 (Tph2) (neuronal) is found mainly in serotonergic neurons of the brain 44. Although mainly expressed in the peripheral tissues (e.g., gut, skin, pineal and gland), tryptophan hydroxylase 1 (Tph1) is also reported to be present in the brain and spinal cord 45. Similarly, tryptophan hydroxylase 2 (Tph2) is also detected in the aorta-gonad-mesonephros region of zebrafish and mouse embryos by RNA sequencing and immunofluorescence assay 46, 47. 5-hydroxytryptophan (5-HTP) is subsequently converted into serotonin through the decarboxylation process mediated by aromatic amino acid decarboxylase (AAAD) 41, 47. Other than converting 5-hydroxytryptophan (5-HTP) into serotonin, aromatic amino acid decarboxylase (AAAD) can also participate in other decarboxylation reactions, such as converting L-dopa into dopamine in dopaminergic, noradrenergic and adrenergic neurons 48.
In the pineal gland, serotonin is converted enzymatically to melatonin. Once the biological function of serotonin is accomplished, the metabolism of serotonin would be carried out by the outer mitochondrial membrane enzyme monoamine oxidase A (MAO-A) into 5-hydroxyindole acetic acid (5-HIAA). 5-hydroxyindole acetic acid (5-HIAA) is the metabolite of serotonin without any biological activity 49, 50, which is excreted out of the body by the kidney. The metabolism process of serotonin is mainly processed in the liver.
Not all cells that contain serotonin (5-hydroxytryptamine [5-HT]) synthesize it. For example, platelets do not synthesize serotonin (5-HT); rather, they accumulate serotonin (5-hydroxytryptamine [5-HT]) from plasma by an active-transport mechanism found on the platelet membrane 40. Certain brain cells do synthesize serotonin. The synthesis and primary metabolic pathways of serotonin are shown in Figure 3 below. The initial step in the synthesis of serotonin is the facilitated transport of the amino acid L-tryptophan from blood into brain. The primary source of tryptophan is dietary protein (see Tryptophan food sources below). Certain other neutral amino acids, such as phenylalanine, leucine and methionine, are transported into brain by the same carrier. The entry of tryptophan into brain is related not only to its concentration in blood but is also a function of its concentration in relation to the concentrations of other neutral amino acids such as phenylalanine, leucine and methionine. Consequently, lowering the dietary intake of tryptophan while raising intake of the amino acids (such as phenylalanine, leucine and methionine) that tryptophan competes with for transport into brain lowers the content of serotonin in brain and changes certain behaviors associated with serotonin function. This strategy for lowering the brain content of serotonin has been used clinically to evaluate the importance of brain serotonin in the mechanism of action of psychotherapeutic drugs 51.
According to some evidence, tryptophan, which increases brain serotonin in humans as in experimental animals 52, is an effective antidepressant in mild-to-moderate depression 53. Further, in healthy people with high trait irritability, it increases agreeableness, decreases quarrelsomeness and improves mood 54.
Interestingly, some researchers stated that purified tryptophan and foods containing tryptophan have different effects on brain serotonin. Although purified tryptophan increases brain serotonin, foods containing tryptophan do not 55. This is because tryptophan is transported into the brain by a transport system that is active toward all the large neutral amino acids and tryptophan is the least abundant amino acid in protein. There is competition between the various amino acids for the transport system, so after the ingestion of a meal containing protein, the rise in the plasma level of the other large neutral amino acids will prevent the rise in plasma tryptophan from increasing brain tryptophan. The idea, common in popular culture, that a high-protein food such as turkey will raise brain tryptophan and serotonin is, unfortunately, false 56. Another popular myth that is widespread on the internet is that bananas improve mood because of their serotonin content. Although it is true that bananas contain serotonin, it does not cross the blood–brain barrier 56. α-Lactalbumin, a minor constituent of milk, is one protein that contains relatively more tryptophan than most proteins. Acute ingestion of α-lactalbumin by humans can improve mood and cognition in some circumstances, presumably owing to increased serotonin 57.
Serotonergic neurons contain the enzyme l-tryptophan-5-monooxygenase, more commonly termed tryptophan hydroxylase, which converts tryptophan to 5-hydroxytryptophan (5-HTP) (Figure 3). Tryptophan hydroxylase enzyme is synthesized in serotonergic cell bodies of the raphe nuclei and is found only in cells that synthesize serotonin; its distribution in brain is similar to that of serotonin itself. Tryptophan hydroxylase enzyme requires both molecular oxygen and a reduced pteridine cofactor, such as l-erythro-tetrahydrobiopterin (BH4), for activity. In the enzymatic reaction, one atom of oxygen is used to form 5-HTP (5-hydroxytryptophan) and the other is reduced to water. The pteridine cofactor donates electrons, and the unstable quinonoid dihydrobiopterin that results is regenerated immediately to the tetrahydrobiopterin form by a NADPH-linked enzymatic reaction. If the concentration of tryptophan in serotonergic neurons is assumed to be comparable to that in whole brain, the enzyme would not be saturated with substrate and the formation of serotonin (5-HT) in brain would be expected to rise as the brain concentration of tryptophan increases. This occurs specifically in response to raising the dietary intake of tryptophan. However, the relationship among tryptophan availability, total tissue serotonin (5-HT) concentration and serotonin (5-HT) release is not fully understood.
The other enzyme involved in the synthesis of serotonin, aromatic l-amino acid decarboxylase (AADC), is a soluble pyridoxal-5′-phosphate-dependent enzyme which converts 5-hydroxytryptophan (5-HTP) to serotonin (5-HT) (Figure 3). It has been demonstrated that administration of pyridoxine increases the rate of synthesis of serotonin (5-HT) in monkey brain, as revealed using position emission tomography 40. This presumably reflects a regulatory effect of pyridoxine on aromatic l-amino acid decarboxylase (AADC) activity and raises the interesting issue of the use of pyridoxine supplementation in situations associated with serotonin deficiency.
Aromatic l-amino acid decarboxylase (AADC) is present not only in serotonergic neurons but also in catecholaminergic neurons, where it converts 3,4-dihydroxyphenylalanine (DOPA) to dopamine. However, different pH optima or concentrations of substrate or cofactor are required for optimal activity of the enzyme in brain homogenates when using either 5-hydroxytryptophan (5-HTP) or DOPA (3,4-dihydroxyphenylalanine) as the substrate. cDNAs encoding AADC have been cloned from various species. The encoded protein contains 480 amino acids and has a molecular weight of 54,000 but it appears to exist as a dimer. Characterization of the protein expressed in cells transfected with the cDNA shows that it decarboxylates either DOPA (3,4-dihydroxyphenylalanine) or 5-hydroxytryptophan (5-HTP). Also, in situ hybridization of the mRNA for the enzyme revealed its presence both in serotonergic cells in the dorsal raphe nucleus and in catecholaminergic cells in brain regions containing catecholaminergic nerve cell soma 58. Taken together, these results support the idea that the enzymatic decarboxylation of both DOPA and 5-hydroxytryptophan (5-HTP) is catalyzed by the same enzyme.
Because the decarboxylase enzyme is not saturated with 5-hydroxytryptophan (5-HTP) under physiological conditions, that is, the concentration of 5-hydroxytryptophan (5-HTP) is much less than the Km of 10 μM, it is possible to raise the content of serotonin (5-HT) in brain not only by increasing the dietary intake of tryptophan but also by raising the intake of 5-hydroxytryptophan (5-HTP). This procedure, though, results in the formation of serotonin (5-HT) in cells that would not normally contain it, such as catecholaminergic neurons, because of the nonselective nature of aromatic l-amino acid decarboxylase (AADC).
The initial hydroxylation of tryptophan, rather than the decarboxylation of 5-hydroxytryptophan (5-HTP), appears to be the rate-limiting step in serotonin synthesis. Evidence in support of this view includes the fact that 5-hydroxytryptophan (5-HTP) is found only in trace amounts in brain, presumably because it is decarboxylated about as rapidly as it is formed. As might be expected if the hydroxylation reaction is rate-limiting, inhibition of this reaction results in a marked depletion of the content of serotonin (5-HT) in brain. The enzyme inhibitor most widely used in experiments is parachlorophenylalanine. In vivo, parachlorophenylalanine irreversibly inhibits tryptophan hydroxylase, presumably by incorporating itself into the enzyme to produce an inactive protein. This results in a long-lasting reduction of serotonin (5-HT) levels. Recovery of enzyme activity and serotonin (5-HT) biosynthesis requires the synthesis of new enzyme. Marked increases in levels of mRNA for tryptophan hydroxylase are found in the raphe nuclei 1 to 3 days after administration of parachlorophenylalanine 59.
Figure 3. Serotonin biosynthesis and metabolism
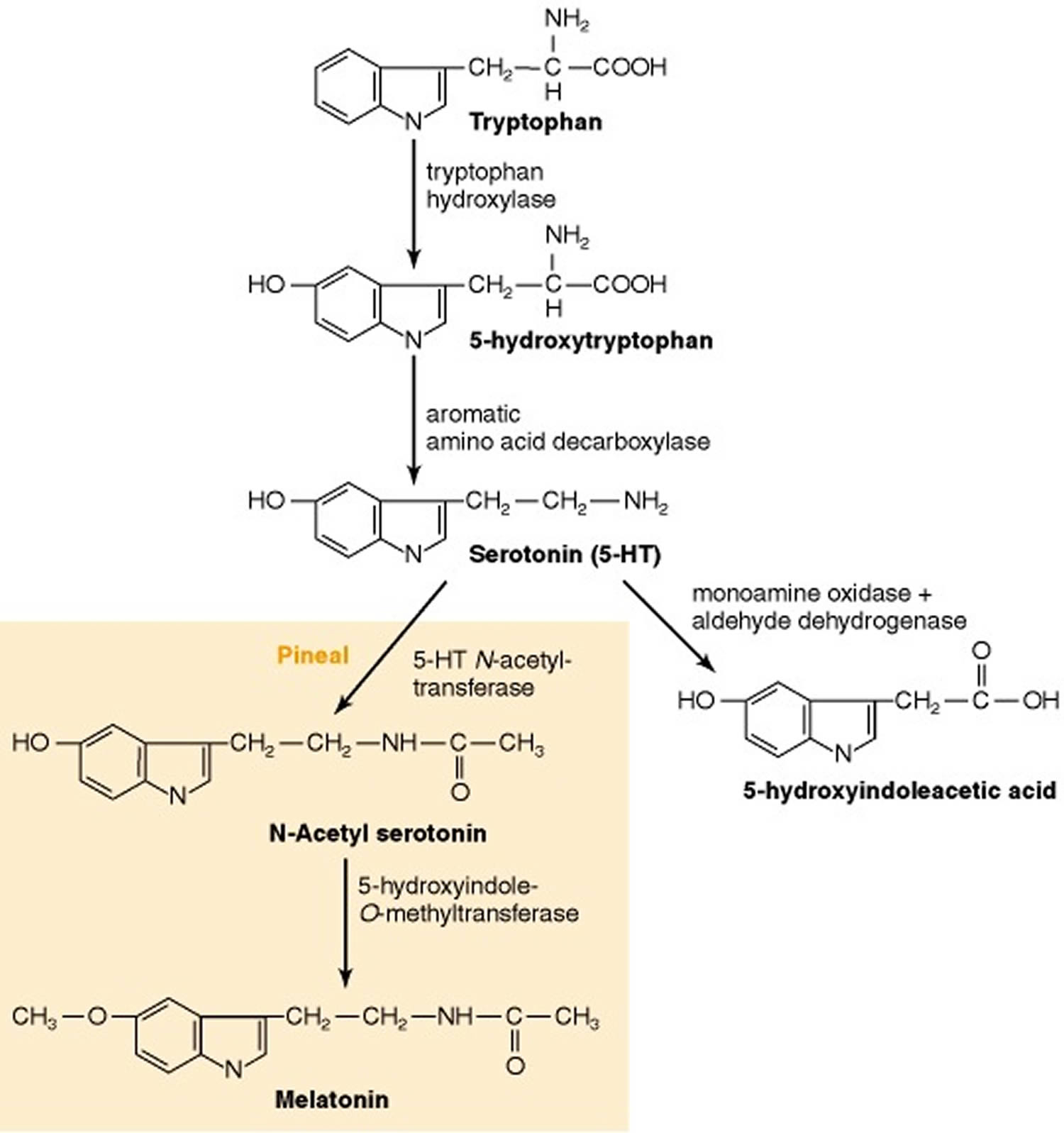
Footnotes: Synthesis and metabolism process of serotonin. The figure shows the serotonin pathways in the gastrointestinal tract and in the central nervous system (brain and spinal cord), the main sites of its synthesis in the human body. Serotonin synthesized in the enterochromaffin cells of the intestine is released into the portal circulation, taken up into platelets, and distributed to various other organs. In animals, serotonin is synthesized from amino acids L-tryptophan. Under the hydroxylation (the addition of -OH [hydroxyl] group) of tryptophan hydroxylase, L-tryptophan is converted into 5-hydroxytryptophan (5-HTP), which is subsequently catalyzed into serotonin by aromatic amino acid decarboxylase (AAAD). Tryptophan hydroxylase 1 (Tph1) and Tryptophan hydroxylase 2 (Tph2) are two forms of tryptophan hydroxylase. In the pineal gland, serotonin is converted enzymatically to melatonin. Once the biological function of serotonin is accomplished, the metabolism of serotonin would be carried out by the outer mitochondrial membrane enzyme monoamine oxidase A (MAO-A) into 5-hydroxyindole acetic acid (5-HIAA). 5-hydroxyindole acetic acid (5-HIAA) is the metabolite of serotonin without any biological activity 49, 50, which is excreted out of the body by the kidney. The metabolism process of serotonin is mainly processed in the liver.
[Source 40 ]The synthesis of serotonin can increase markedly under conditions requiring a continuous supply of the neurotransmitter
Plasticity is an important concept in neurobiology. In general, this refers to the ability of neuronal systems to conform to either short- or long-term demands placed upon their activity or function. One of the processes contributing to neuronal plasticity is the ability to increase the rate of neurotransmitter synthesis and release in response to increased neuronal activity. Serotonergic neurons have this capability; the synthesis of serotonin from tryptophan is increased in a frequency-dependent manner in response to electrical stimulation of serotonergic soma 60. The increase in synthesis results from the enhanced conversion of tryptophan to 5-hydroxytryptophan (5-HTP) and has an absolute dependence on extracellular Ca2+. It is likely that the increased synthesis results in part from alterations in the kinetic properties of tryptophan hydroxylase, perhaps due to calcium-dependent phosphorylation of the enzyme. The enzyme can be phosphorylated directly by the action of calmodulin-dependent protein kinase II; an activator protein appears to be required for this interaction. In the presence of the activator, tryptophan hydroxylase also may be a substrate for cAMP-dependent protein kinase (PKA). The increased activity of tryptophan hydroxylase does not result from the removal of enzyme inhibition caused by either serotonin or 5-hydroxytryptophan (5-HTP).
Short-term requirements for increases in the synthesis of serotonin can be met by processes that change the kinetic properties of tryptophan hydroxylase, such as phosphorylation, without necessitating the synthesis of more molecules of tryptophan hydroxylase. By contrast, situations requiring long-term increases in the synthesis and release of serotonin result in the synthesis of tryptophan hydroxylase protein. For example, partial but substantial destruction of >60% of central serotonergic neurons results in an increase in the synthesis of serotonin in residual terminals. The increase in synthesis initially results from activation of existing tryptophan hydroxylase molecules, but the increased synthesis of serotonin seen weeks after the lesion results from more tryptophan hydroxylase being present in the residual terminals. An increase in tryptophan hydroxylase mRNA has been reported in residual raphe serotonergic neurons after partial lesioning, consistent with the idea of an increase in the synthesis of tryptophan hydroxylase molecules in residual neurons.
As with other biogenic amine transmitters, serotonin is stored primarily in vesicles and released by an exocytotic mechanism
Peripheral sources of monoamine-containing cells have been utilized to study the properties of storage vesicles, such as chromaffin cells of the adrenal medulla for catecholamines and parafollicular cells of the thyroid gland for serotonin. In some respects, the vesicles that store serotonin (5-HT) resemble those that store catecholamines. For example, drugs such as reserpine and tetrabenazine, which inhibit the activity of the transporter localized to the vesicular membrane, deplete the brain content of serotonin as well as of catecholamines. Storage of serotonin in vesicles requires its active transport from the cytoplasm. The vesicular transporter uses the electrochemical gradient generated by a vesicular H+-ATPase to drive transport, such that a cytoplasmic amine is exchanged for a luminal proton; that is, uptake of serotonin (5-HT) is coupled to efflux of H+ (Figure 4).
Vesicles storing 5-HT exhibit some differences from those storing catecholamines. In contrast to catecholamine-containing vesicles, there is virtually no ATP in serotonin vesicles. Also, serotonergic synaptic vesicles, but not chromaffin granules, contain a specific protein that binds serotonin (5-HT) with high affinity. This serotonin-binding protein is present in serotonergic cells derived ontogenetically from the neuroectoderm 61. It binds serotonin (5-HT) with high affinity in the presence of Fe2+. There are three isoforms of serotonin-binding protein. The 45-kDa isoform appears to be packaged in secretory vesicles along with serotonin (5-HT), which probably accounts for the observation that newly taken up [3H]serotonin (5-HT) is rapidly complexed with this isoform in brain in situ. This isoform is secreted along with serotonin (5-HT) by a calcium-dependent process.
There is considerable evidence that the release of serotonin (5-HT) occurs by exocytosis, that is, by the discharge from the cell of the entire contents of individual storage vesicles. First, 5-HT is ionized sufficiently at physiological pH so that it does not cross plasma membranes by simple diffusion. Second, most intraneuronal 5-HT is contained in storage vesicles and other contents of the vesicle, including serotonin-binding protein (SPB), are released together with serotonin. By contrast, cytosolic proteins do not accompany electrical stimulation-elicited release of 5-HT. Third, the depolarization-induced release of serotonin (5-HT) occurs by a calcium-dependent process; indeed, it appears that the influx of Ca2+ with or without membrane depolarization can increase the release of serotonin (5-HT). Ca2+ has been reported to stimulate the fusion of vesicular membranes with the plasma membrane.
The rate of serotonin release is dependent on the firing rate of serotonergic soma in the raphe nuclei. Numerous studies have revealed that an increase in raphe cell firing enhances the release of serotonin (5-HT) in terminal fields. The opposite effect is observed when raphe cell firing decreases. This means that drugs that change the firing rate of serotonergic soma modify the release of serotonin as well. An important target for such drugs is the somatodendritic autoreceptor, which, as discussed later, is the 5-HT1A receptor subtype (see Figure 5). Administration of 5-HT1A agonists, such as 8-hydroxy-2-(di-n-propylamino)-tetralin (8-OH-DPAT), into the dorsal raphe nucleus slows the rate of firing of serotonergic soma. Using the technique of in vivo microdialysis, application of 8-OH-DPAT in the dorsal raphe nucleus decreases the release of 5-HT in the striatum. Depending on the species, serotonergic autoreceptors in terminal fields appear to be either the 5-HT1B or the 5-HT1D subtype. Administration of agonists of these receptors into areas receiving serotonergic innervation decreases the synthesis and release of 5-HT measured in vitro or in situ, using the technique of microdialysis. However, in contrast to the activation of somatodendritic autoreceptors, such effects are not due to decreases in the firing rate of serotonergic soma.
Figure 4. Serotonin release and storage
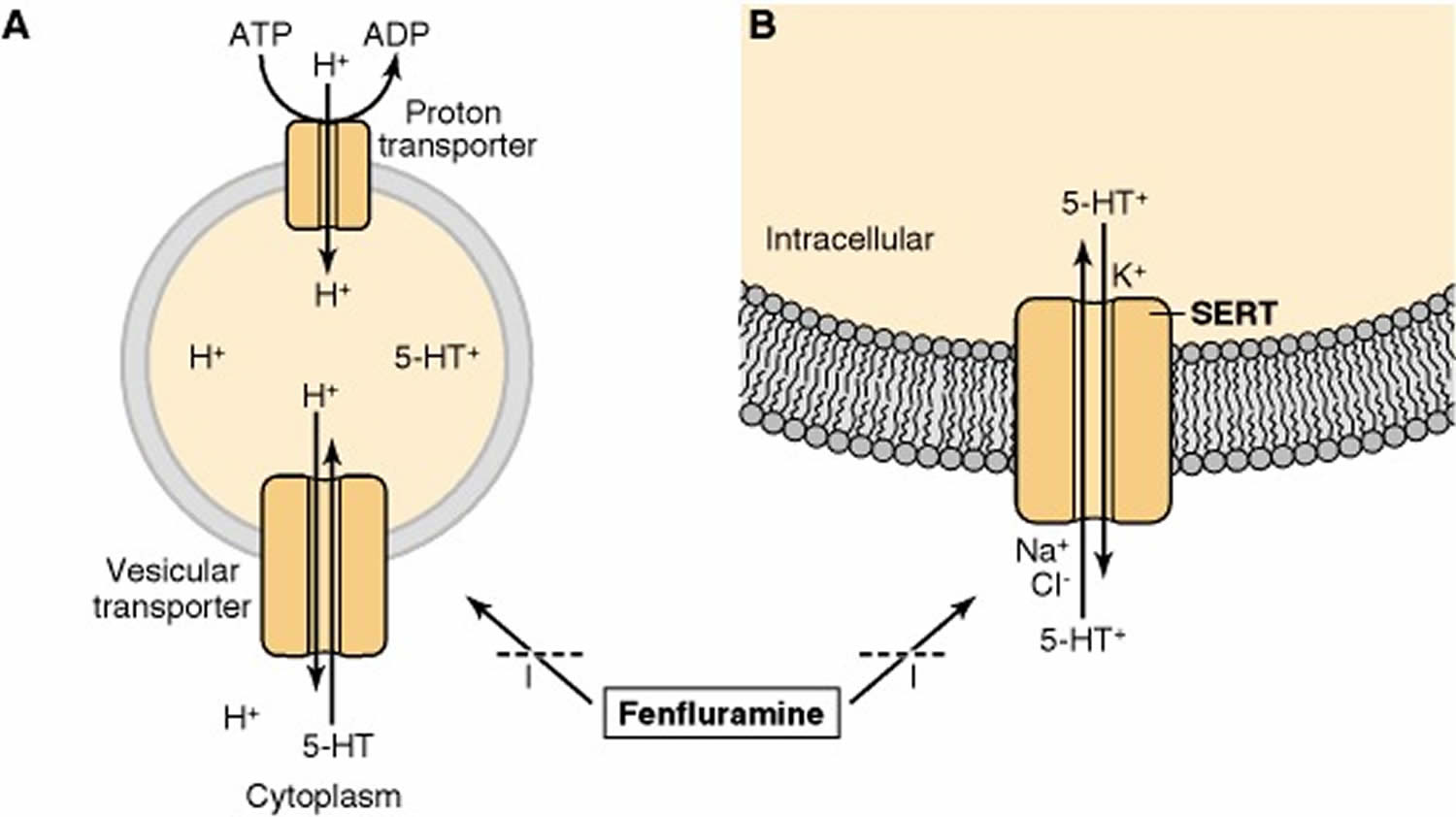
Footnote: The substituted amphetamine fenfluramine inhibits the transport of serotonin (5-HT) by both (A) the vesicular transporter and (B) the serotonin transporter (SERT). Substituted amphetamines, such as fenfluramine and 3,4-methylenedioxymethamphetamine (MDMA), stimulate the release of serotonin (5-HT) from serotonergic terminals. These drugs block the vesicular transporter and disrupt the proton gradient across the vesicle membrane. The increase in intracellular serotonin (5-HT) favors the release of serotonin (5-HT) by the reverse action of the SERT. These drugs also act as substrates for the SERT so as to inhibit the transport of serotonin (5-HT) into cells.
[Source 40]Figure 5. Effects of psychoactive drugs on serotonergic neurotransmission
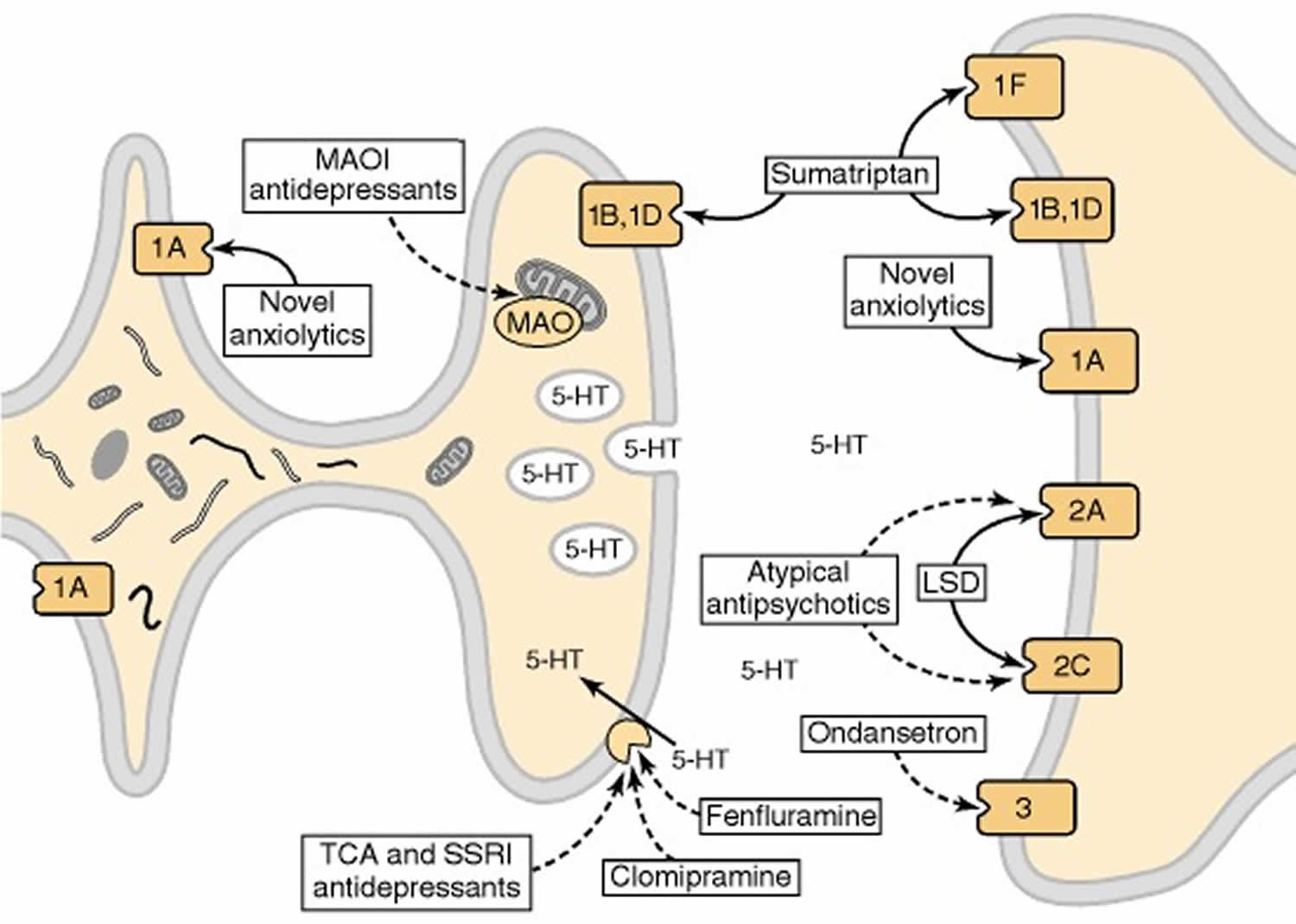
Footnote: Drugs that act as agonists are indicated by solid-line arrows, whereas antagonists or inhibitors are shown with broken-line arrows. The serotonin receptor 1A receptor (5-HT1AR) acts as both the somatodendritic autoreceptor and a postsynaptic receptor. Anxiolytic drugs, such as buspirone, are agonists at this receptor. In terminal fields, the autoreceptor is either the serotonin receptor 1B receptor (5-HT1B) or serotonin receptor 1D receptor (5-HT1D) subtype; these receptors also function as postsynaptic receptors. The antimigraine drug sumatriptan is an agonist at these receptors as well as at the 5-HT1F receptor. Hallucinogenic drugs, such as LSD, are agonists at serotonin receptor 2A receptor (5-HT2A) and serotonin receptor 2C receptor (5-HT2C) receptors, whereas atypical antipsychotic drugs, such as clozapine and olanzapine, are antagonists. The 5-HT3 receptor, a ligand-gated ion channel, is blocked by drugs effective in the treatment of chemotherapy-induced nausea and emesis, such as ondansetron. Another important target for psychotherapeutic drugs is the serotonin transporter, which is blocked by drugs effective in the treatment of depression or obsessive-compulsive disorder, such as clomipramine. The enzyme responsible for the catabolism of serotonin, monoamine oxidase (MAO), is inhibited by another class of antidepressants. MAOI (monoamine oxidase inhibitor); TCA (tricyclic antidepressant); SSRI (selective serotonin reuptake inhibitor).
[Source 40]The activity of serotonin in the synapse is terminated primarily by its re-uptake into serotonergic terminals
Synaptic effects of many amino acid and monoaminergic neurotransmitters, including serotonin, are terminated by binding of these molecules to specific transporter proteins. The serotonin transporter (SERT) is located on serotonergic neurons. Evidence for this comes from studies showing that the selective lesioning of serotonergic neurons in brain markedly reduces both the high-affinity uptake of [3H]5-HT in areas of brain receiving serotonergic innervation and the specific binding of radioligands to the serotonin transporter. Activity of the serotonin transporter (SERT) regulates the concentration of serotonin in the synapse, thereby influencing synaptic transmission.
The uptake system for serotonin is saturable and of high affinity, with a Km value for serotonin of approximately 0.2 to 0.5 μM. Uptake of serotonin is an active process that is temperature-dependent and has an absolute requirement for external Na+ and C1−; it is inhibited by metabolic inhibitors as well as by inhibitors of Na/K ATPase activity. From these and other data, it has been inferred that the energy requirement for serotonin uptake is not used directly to transport serotonin but rather is necessary to maintain the gradient of Na+ across the plasma membrane, upon which serotonin uptake is dependent. The current model of transport has one Na+, one C1− and one protonated serotonin binding to the transporter extracellularly prior to translocation to form a quaternary complex that subsequently undergoes a conformational change to release the neurotransmitter and the ions into the cytoplasm. The conformational change may involve the “opening” of a pore formed by some portion of the transmembrane domains of the serotonin transporter (SERT). In the cytoplasm, K+ associates with the serotonin transporter (SERT) to promote reorientation of the unloaded carrier for another transport cycle (Figure 4).
Serotonin breakdown (metabolism)
The primary metabolic (catabolic) pathway for serotonin (5-hydroxytryptamine [5-HT]) is oxidative deamination by the outer mitochondrial membrane enzyme monoamine oxidase (MAO). Monoamine oxidase (MAO) converts serotonin (5-hydroxytryptamine [5-HT]) to 5-hydroxyindoleacetaldehyde, and this product is oxidized by an NAD+-dependent aldehyde dehydrogenase to form 5-hydroxyindoleacetic acid (5-HIAA) (Figure 3), which can be excreted by the kidneys. The intermediate acetaldehyde also can be reduced by an NADH-dependent aldehyde reductase to form the alcohol 5-hydroxytryptophol. Whether oxidation or reduction takes place depends on the ratio of NAD+ to NADH in the tissue. In brain, 5-hydroxyindoleacetic acid (5-HIAA) is the primary metabolite of serotonin. 5-hydroxyindole acetic acid (5-HIAA) is the metabolite of serotonin without any biological activity 49, 50, which is excreted out of the body by the kidney. The metabolism process of serotonin is mainly processed in the liver.
There are at least two isoenzymes of MAO (monoamine oxidase), referred to as MAO-A (monoamine oxidase type A) and MAO-B (monoamine oxidase type B), are encoded by distinct genes on the human X chromosome 62. These MAO (monoamine oxidase) isoenzymes are integral flavoproteins of outer mitochondrial membranes in neurons, glia and other cells. Both monoamine oxidase (MAO) isoenzymes catalyze oxidative deamination of serotonin and various other endogenous and dietary monoamines, with MAO-A (monoamine oxidase type A) having preferential affinity for serotonin (5-hydroxytryptamine [5-HT]) over other substrates 63. Serotonin (5-hydroxytryptamine [5-HT]) can also be converted into melatonin via two enzymatic steps. Evidence for the existence of isoenzymes was based initially on differing substrate specificities and sensitivities to inhibitors of MAO. For example, both serotonin and norepinephrine are metabolized preferentially by MAO-A (monoamine oxidase type A). Selective inhibitors of each form of the enzyme monoamine oxidase (MAO) exist: clorgyline or moclobemide for type A and deprenyl for type B. Definitive proof of the existence of these two forms of monoamine oxidase (MAO) comes from the cloning of cDNAs encoding subunits of MAO-A and MAO-B from human liver 64. The deduced amino acid sequences of MAO-A and MAO-B show about 70% homology and have masses of 59.7 and 58.8 kDa, respectively. When each cDNA was cloned into an expression vector and transfected independently into a cell line, the activity of the proteins expressed resembled that of the endogenous enzymes from human brain, such that the expressed MAO-A preferred serotonin as a substrate and was inhibited preferentially by clorgyline. From such data it was inferred that the functional differences between these two enzymes exist in their primary structures.
The use of transgenic mice permits the selective elimination, or “knockout,” of either MAO type 65. In the brains of mice deficient in MAO-A, the content of serotonin is elevated markedly for about 12 days after birth and then slowly declines, reaching values comparable to those in normal mice after about 7 months. In MAO-A-deficient mice, the selective inhibitor of MAO-B, deprenyl, had a greater effect on serotonin metabolism than it did in normal mice. Such observations indicate that, in the absence of MAO-A, MAO-B can metabolize serotonin in vivo. However, mice lacking the MAO-B isoenzyme do not have elevated levels of serotonin in brain. Of interest are the aggressive behaviors exhibited by mice deficient in MAO-A, consistent with a postulated role of serotonergic neurons in human aggressive behaviors.
Serotonin function
The function of brain serotonin remains an enigma 66. There have been several attempts to identify a unifying function of serotonin 67, 68. Most researchers acknowledge that the function of the serotonin system remains ‘elusive’ 69 and ‘a puzzle’ 70 and it is argued that this may be due to the special diversity and complexity of the serotonin system with its many receptor subtypes 71, extensive innervation of the brain and paracrine style of transmission 72. The notion that serotonin is an enigma among neuromodulators [said to be ‘involved in everything but responsible for nothing’ 73] and it is argued that the riddle of serotonin can only be solved by focusing on its individual receptor subtypes 66.
Accordingly, given the inherent complexity of the serotonin system, one strategy for understanding its functioning is to focus on a select number of receptor subtypes that have been particularly well characterized. From this foundation, one might then consider whether other serotonin receptor subtypes can be incorporated into the associated model, or whether one or more additional models are required to cover the full range of functions associated with brain serotonin transmission. Broadly consistent with prior theories 74, Carhart-Harris and Nutt 66 believed that a key function of brain serotonin (5-HT) is to moderate anxiety and stress, and promote patience and coping 75 via (postsynaptic) 5-HT1AR (serotonin receptor 1AR) signalling. Moreover, Carhart-Harris and Nutt 66 also extend on this by proposing that a second major function of brain serotonin is to open a window of plasticity for greater adaptation 76, mediated in large part by 5-HT2AR (serotonin receptor 2AR) signalling. This bipartite model is consistent with a ‘flexible coping’ model of brain serotonin function, in which postsynaptic 5-HT1ARs mediate so-called ‘passive coping’ (i.e. tolerating but not necessarily dealing with a source of psychological pain) and 5-HT2ARs mediate ‘active coping’ (actively dealing with a source of psychological pain by changing one’s relationship to it) 77.
The brain serotonin (5-hydroxytryptamine) system is thought to play an essential role in the control of anxiety-, fear- and panic-like responses in rodents 78. In humans, several lines of evidence link alterations in serotonin (5-hydroxytryptamine) signaling to panic attacks through a defensive behavioral system activated by acute threats 79. Panic attacks represent abrupt surges of intense fear or extreme discomfort that reach a peak within minutes. Unexpected panic attacks with at least one of the attacks followed by persistent concern or worry about additional panic attacks or their consequences define panic disorder 80. Maladaptive changes in behavior related to the attack, such as agoraphobia, describe anticipatory anxiety and/or marked fear about apparently threatening situations.
Serotonin (5-HT) may influence anxiety-, fear- and panic-like responses within the basolateral complex of the amygdala. In humans, alteration in the Tryptophan hydroxylase 2 gene (TPH2) were associated with alterations in amygdala responsiveness to anxiety-related stimuli 81 as well as anxiety 82 and panic disorders 83. This has been linked recently to alterations in the GABAergic system 84. In mice, pharmacological depletion of serotonin (5-HT) in the basolateral complex of the amygdala reduces anxiety and interferes with fear conditioning 85. Furthermore, stress-induced enhancement of fear memory is dependent on serotonin (5-HT) action within the basolateral complex of the amygdala 86.
Exaggerated fear accompanied by a panic-like state and escape behavior resulting from lifelong absence of serotonin synthesis involves dysfunction of the amygdalo-dorsal raphe circuitry controlling fear-related behavioral responses 87. This is due to alterations in GABAergic transmission prohibiting increased activity of the basolateral nucleus in aversive inescapable contexts. Increased escape responses reflect characteristic syndromal dimensions of panic disorder and agoraphobia. Thus, constitutive lack of serotonin synthesis influences the risk for anxiety- and stressor-related disorders including panic disorder and comorbid agoraphobia through the absence of GABAergic-dependent compensatory mechanisms in the basolateral complex of the amygdala.
Bowel Function
Most of the body’s serotonin is in the gastrointestinal tract, regulating bowel function and movements 1. The human digestive tract is composed of several layers of enterochromaffin cells. These cells sense food in the stomach and release serotonin as a response. Increased serotonin levels in the gut cause digestive processes to increase speed, often due to digesting toxins or noxious substances. It also plays a part in reducing appetite while eating 2.
Mood
Serotonin plays a significant role in the nervous system and largely affects mood. In the brain, serotonin changes mood, anxiety, and happiness by increasing nerve stimulation and electrical impulses 1. Drugs like ecstasy and LSD increase the levels of serotonin in the brain to produce effects like increased appetite, increased sexual drive, euphoria, and even hallucinations 88.
Blood clotting
When serotonin is released into the blood, it is often absorbed by platelets rather than remaining free serotonin 1. The effect serotonin has on the platelets is similar to those produced by the interaction of platelet factor 2 and platelets 89. Serotonin accelerates the metabolism of fibrinogen to fibrin. This action causes platelet aggregation, leading to vasoconstriction, resulting in a reduction in the blood flow and an increase in clot formation; this is 1 of the earliest defined functions of serotonin 90.
Serotonin has also been identified to play important roles in megakaryocytopoiesis (the cellular developmental process by which megakaryocytes, the precursor cells that produce platelets, develop from hematopoietic stem cells) 7. Although most of serotonin is stored in the platelets, a small quantity of serotonin can also be stored in the megakaryocytes. Megakaryocytes are the progenitors of platelets and the only cells to take up serotonin in the bone marrow 91. Serotonin receptors have been reported to be expressed on the surface of most megakaryocytes. The in vitro study shows that serotonin can enhance megakaryocyte colony formation ability and the mitogenic effect of serotonin on megakaryocytopoiesis may be mediated through the serotonin receptor subtype 2A (Htr2 receptor) 92. A recent study has demonstrated that serotonin receptor subtype 2b (Htr2b receptor) is expressed in megakaryocytes and that serotonin mediated by serotonin receptor subtype 2b (Htr2b receptor) can enhance the proliferation and inhibit the apoptosis of megakaryocytes. This study also shows that serotonin can activate ERK signaling and affect F-actin reorganization to promote megakaryopoiesis and proplatelet formation 93.
Nausea
When serotonin is released into the gut faster than it can be digested, it is reabsorbed into the bloodstream. In the bloodstream, serotonin can interact with 5-HT3 receptors, activating chemoreceptor trigger zones. The activation of these sites causes stimulation of the brain to cause expulsion of the substance eaten; this is perceived as nausea 94.
Bone Density
Several research studies have shown that serotonin may have links to a decrease in bone density, but the relationship lacks sufficient proof 1. This correlation has been hypothesized by an early study that measured the changes in mice that lacked brain serotonin. The researchers found that these mice have severe osteopenia, while mice lacking intestinal serotonin have regular bone density. Humans with increased blood serotonin levels have been linked to increased or regular bone density. The belief is that the 5-HT1B receptor is the link between blood serotonin and bone density 95.
Sexual Function
Serotonin has been linked to increased sexual drive but decreased sexual function; this can be rather detrimental to patients who have received a prescription for selective serotonin reuptake inhibitors (SSRIs) 1. As selective serotonin reuptake inhibitors (SSRIs) drugs are commonly given to patients suffering from depression, they can often exacerbate the underlying problem due to sexual tension. The main cause of serotonin’s role in decreased sexual function is increasing retrograde ejaculation and erectile dysfunction 1. The cause of these symptoms is the overstimulation of the 5-HT2C receptors, which promote erection and inhibit ejaculation, and the under-stimulation of the 5-HT1A receptor, which has inverse functions 96.
Serotonin and depression
The “serotonin hypothesis” of clinical depression is almost 50 years old 97. At its simplest, the hypothesis proposes that diminished activity of serotonin pathways plays a causal role in the pathophysiology of depression. This notion was based on the depressogenic effects of amine depleting agents such as reserpine, as well as the actions of antidepressant drugs such as monoamine oxidase inhibitors and tricyclic antidepressants, discovered by clinical serendipity, but later found in animal experimental studies to potentiate the effects of serotonin and other monoamines at the synapse 98.
The best evidence that serotonin plays a role in the pathophysiology of depression comes from studies of “tryptophan depletion”, where an acute dietary manipulation is employed to produce a transient lowering in brain serotonin activity through diminishing availability of its precursor amino acid, tryptophan 97. In healthy participants with no risk factors for depression, tryptophan depletion does not produce clinically significant changes in mood; however, recovered depressed patients free of medication can show brief, clinically relevant, depressive symptomatology 99. Interestingly, the same is true of recovered depressed patients undergoing catecholamine depletion with alpha-methyl-para-tyrosine 100.
Overall, this evidence suggests that impairing serotonin function can cause clinical depression in some circumstances, but is neither necessary nor sufficient. In addition, the depressogenic effects of tryptophan depletion are much more apparent in people who have experienced prior episodes of depression than in those simply at high risk of illness, for example by virtue of a strong family history 101. This suggests that low serotonin function may compromise mechanisms involved in maintaining recovery from depression rather than having a primary effect to lower mood in all vulnerable people.
These findings also hint at a role for diminished tryptophan availability in triggering depression, particularly in people with a previous history of illness. Interestingly, lower plasma levels of tryptophan are one of the few reasonably robust findings in patients with more severe forms of depression 102 and, more recently, have been linked to peripheral inflammation and consequent induction of the tryptophan metabolizing enzyme indoleamine 2,3-dioxygenase 103. Inflammation could therefore produce depression in vulnerable individuals by lowering plasma tryptophan and diminishing brain serotonin activity. Conceivably, such an effect could explain the diminished efficacy of SSRIs in depressed patients with high levels of inflammatory biomarkers 104.
Serotonin and antidepressant action
Undoubtedly, a major reason for the continuing interest in serotonin and depression is the fact that SSRIs are useful antidepressant drugs for some patients. Elegant basic studies have revealed intriguing molecular and cellular consequences of repeated SSRI administration in animals, for example increases in hippocampal cell proliferation and enhanced expression of neuroplasticity related proteins such as brain derived neurotrophic factor (BDNF) 105. However, linking such changes to resolution of the clinical depressive syndrome is challenging. More pertinent in this respect are neuropsychological studies which show that, in both healthy participants and depressed patients, administration of SSRIs leads to positive shifts in the way the brain appraises emotionally-valenced information. This effect occurs very early in treatment, prior to clinical antidepressant effects, and appears to be mediated via serotonergic innervation to limbic circuitry, particularly the amygdala 106.
This work gives a new insight into how serotonin pathways may influence mood in depressed patients, that is by altering the way the brain appraises emotionally-laden information at an implicit level. Unlike mood, emotions are relatively short-lived, automatic responses to internal or external stimuli, and in depressed patients emotional responses are reliably negatively biased 107. Thus, from this viewpoint, increasing serotonin activity in depressed people does not influence subjective mood directly but, rather, as a secondary consequence of positive shifts in automatic emotional responses.
Over time, it is suggested, this positive biasing of automatic processing would, in an appropriate interpersonal environment, lead to changes in the strategic processing associated with conscious emotional experience. This psychological process is likely to involve re-learning a range of emotional associations, which might account for the gradual onset of clinical antidepressant activity 106. In addition, the notion that “re-learning” is involved in subjective improvement in depression sits well with the finding noted above that antidepressants such as SSRIs promote synaptic plasticity, an effect classically associated with learning 108.
How to increase serotonin
Perreau-Linck and colleagues 109 provides an initial lead about one possible strategy for raising brain serotonin. Using positron emission tomography (PET), they obtained a measure of serotonin synthesis in the brains of healthy participants who underwent positive, negative and neutral mood inductions. Reported levels of happiness were positively correlated and reported levels of sadness were negatively correlated with serotonin synthesis in the right anterior cingulate cortex 109. The idea that alterations in thought, either self-induced or due to psychotherapy, can alter brain metabolism is not new. Numerous studies have demonstrated changes in blood flow in such circumstances. However, reports related to specific transmitters are much less common. In one recent study, meditation was reported to increase release of dopamine 110. The study by Perreau-Linck and colleagues 111 is the first to report that self-induced changes in mood can influence serotonin synthesis. This raises the possibility that the interaction between serotonin synthesis and mood may be 2-way, with serotonin influencing mood and mood influencing serotonin. Obviously, more work is needed to answer questions in this area. For example, is the improvement in mood associated with psychotherapy accompanied by increases in serotonin synthesis? If more precise information is obtained about the mental states that increase serotonin synthesis, will this help to enhance therapy techniques?
Exposure to bright light is a second possible approach to increasing serotonin without drugs. Bright light is, of course, a standard treatment for seasonal affective disorder (SAD) or seasonal depression, but a few studies also suggest that it is an effective treatment for nonseasonal depression 112 and also reduces depressed mood in women with premenstrual dysphoric disorder 113 and in pregnant women suffering from depression (antepartum depression) 114. The evidence relating these effects to serotonin is indirect. In human postmortem brain, serotonin levels are higher in those who died in summer than in those who died in winter 115. A similar conclusion came from a study on healthy volunteers, in which serotonin synthesis was assessed by measurements of the serotonin metabolite 5-hydroxyindoleacetic acid (5-HIAA) in the venous outflow from the brain 116. There was also a positive correlation between serotonin synthesis and the hours of sunlight on the day the measurements were made, independent of season. In rats, serotonin is highest during the light part of the light–dark cycle, and this state is driven by the photic cycle rather than the circadian rhythm 117. The existence of a retinoraphe tract may help explain why, in experimental animals, neuronal firing rates, c-fos expression and the serotonin content in the raphe nuclei are responsive to retinal light exposure 118. In humans, there is certainly an interaction between bright light and the serotonin system. The mood-lowering effect of acute tryptophan depletion in healthy women is completely blocked by carrying out the study in bright light (3000 lux) instead of dim light 119.
Relatively few generations ago, most of the world population was involved in agriculture and was outdoors for much of the day. This would have resulted in high levels of bright light exposure even in winter. Even on a cloudy day, the light outside can be greater than 1000 lux, a level never normally achieved indoors. In a recent study carried out at around latitude 45° North, daily exposure to light greater than 1000 lux averaged about 30 minutes in winter and only about 90 minutes in summer 120 among people working at least 30 hours weekly; weekends were included. In this group, summer bright light exposure was probably considerably less than the winter exposure of our agricultural ancestors. Today we may be living in a bright light–deprived society. A large literature exists on the beneficial effect of bright light exposure in healthy individuals 56. Lamps designed for the treatment of seasonal affective disorder (SAD), which provide more lux than is ever achieved by normal indoor lighting, are readily available, although incorporating their use into a daily routine may be a challenge for some. “Light cafes” pioneered in Scandinavia have come to the United Kingdom 121 and an Austrian village that receives no sunshine in the winter because of its surrounding mountains is building a series of giant mirrors to reflect sunlight into the valley 122. Better use of daylight in buildings is an issue that architects are increasingly aware of. Working indoors does not have to be associated with suboptimal exposure to bright light.
A third strategy that may raise brain serotonin is exercise. A comprehensive review of the relation between exercise and mood concluded that antidepressant and anxiolytic effects have been clearly demonstrated 123. In the United Kingdom the National Institute for Health and Clinical Excellence (NICE), which works on behalf of the National Health Service and makes recommendations on treatments according to the best available evidence, has published a guide on the treatment of depression 124. The guidance recommends treating mild clinical depression with various strategies, including exercise rather than antidepressants, because the risk–benefit ratio is poor for antidepressant use in patients with mild depression. Exercise improves mood in subclinical populations as well as in patients. The most consistent effect is seen when regular exercisers undertake aerobic exercise at a level with which they are familiar 125. In support of the National Institute for Health and Clinical Excellence recommendations, the National Institute of Mental Health in the United States funded clinical trial called Treatment with Exercise Augmentation for Depression (TREAD) study found “symptoms improve with exercise treatment for depression, and this change correlates well with overall outcome” 126. And “changes in motivation and energy were larger, as measured by effect size, than changes in anhedonia” 126. Anhedonia is the inability to feel pleasure in normally pleasurable activities.
Several lines of research suggest that exercise increases brain serotonin function in the human brain. Post and colleagues 127 measured biogenic amine metabolites in cerebrospinal fluid (CSF) of patients with depression before and after they increased their physical activity to simulate mania. Physical activity increased 5-hydroxyindoleacetic acid (5-HIAA), but it is not clear that this was due to increased serotonin turnover or to mixing of CSF from higher regions, which contain higher levels of 5-hydroxyindoleacetic acid (5-HIAA), with lumbar CSF (or to a combination of both mechanisms). Nonetheless, this finding stimulated many animal studies on the effects of exercise. For example, Chaouloff and colleagues 128 showed that exercise increased tryptophan and 5-hydroxyindoleacetic acid (5-HIAA) in rat ventricles. More recent studies using intracerebral dialysis have shown that exercise increases extracellular serotonin and 5-HIAA in various brain areas, including the hippocampus and cortex 129. Two different mechanisms may be involved in this effect. As reviewed by Jacobs and Fornal 130, motor activity increases the firing rates of serotonin neurons, and this results in increased release and synthesis of serotonin 131. In addition, there is an increase in the brain of the serotonin precursor tryptophan that persists after exercise 132.
The largest body of work in humans looking at the effect of exercise on tryptophan availability to the brain is concerned with the hypothesis that fatigue during exercise is associated with elevated brain tryptophan and serotonin synthesis. A large body of evidence supports the idea that exercise, including exercise to fatigue, is associated with an increase in plasma tryptophan and a decrease in the plasma level of the branched chain amino acids (BCAAs) leucine, isoleucine and valine 133. The branched chain amino acids (BCAAs) inhibit tryptophan transport into the brain 134. Because of the increase in plasma tryptophan and decrease in BCAA, there is a substantial increase in tryptophan availability to the brain. Tryptophan is an effective mild hypnotic 53, a fact that stimulated the hypothesis that it may be involved in fatigue. It is notable that several clinical trials of branched chain amino acid (BCAA) investigated whether it was possible to counter fatigue by lowering brain tryptophan, with results that provided little support for the hypothesis. Furthermore, exercise results in an increase in the plasma ratio of tryptophan to the branched chain amino acids (BCAAs) before the onset of fatigue 133. The conclusion of these studies is that, in humans, a rise in precursor availability should increase serotonin synthesis during and after exercise and that this is not related to fatigue, although it may be related to improved mood. Whether motor activity increases the firing rate of serotonin neurons in humans, as in animals, is not known. However, it is clear that aerobic exercise can improve mood.
As with exposure to bright light, there has been a large change in the level of vigorous physical exercise experienced since humans were hunter-gatherers or engaged primarily in agriculture. Lambert 135 argued that the decline in vigorous physical exercise and in particular, in effort-based rewards may contribute to the high level of depression in today’s society. The effect of exercise on serotonin suggests that the exercise itself, not the rewards that stem from exercise, may be important. If trials of exercise to prevent depression are successful, then prevention of depression can be added to the numerous other benefits of exercise.
The fourth factor that could play a role in raising brain serotonin is diet. According to some evidence, tryptophan, which increases brain serotonin in humans as in experimental animals 52, is an effective antidepressant in mild-to-moderate depression 53. Further, in healthy people with high trait irritability, it increases agreeableness, decreases quarrelsomeness and improves mood 54. Treating tryptophan as a drug is reasonable because, first, there is normally no situation in which purified tryptophan is needed for dietary reasons, and second, purified tryptophan and foods containing tryptophan have different effects on brain serotonin. Although purified tryptophan increases brain serotonin, foods containing tryptophan do not 55. This is because tryptophan is transported into the brain by a transport system that is active toward all the large neutral amino acids and tryptophan is the least abundant amino acid in protein. There is competition between the various amino acids for the transport system, so after the ingestion of a meal containing protein, the rise in the plasma level of the other large neutral amino acids will prevent the rise in plasma tryptophan from increasing brain tryptophan. The idea, common in popular culture, that a high-protein food such as turkey will raise brain tryptophan and serotonin is, unfortunately, false 56. Another popular myth that is widespread on the internet is that bananas improve mood because of their serotonin content. Although it is true that bananas contain serotonin, it does not cross the blood–brain barrier 56.
α-Lactalbumin, a minor constituent of milk, is one protein that contains relatively more tryptophan than most proteins. Acute ingestion of α-lactalbumin by humans can improve mood and cognition in some circumstances, presumably owing to increased serotonin 57. Enhancing the tryptophan content of the diet chronically with α-lactalbumin is probably not practical. However, increasing the tryptophan content of the diet relative to that of the other amino acids is something that possibly occurred in the past and could occur again in the future. Kerem and colleagues 136 studied the tryptophan content of both wild chickpeas and the domesticated chickpeas that were bred from them in the Near East in neolithic times. The mean protein content (per mg dry seed) was similar for 73 cultivars and 15 wild varieties. In the cultivated group, however, the tryptophan content was almost twice that of the wild seeds. Interestingly, the greater part of the increase was due to an increase in the free tryptophan content (i.e., not part of the protein). In cultivated chickpeas, almost two-thirds of the tryptophan was in the free form. Kerem and colleagues 136 argue that there was probably selection for seeds with a higher tryptophan content. This is plausible, given another example of an early strategy to increase the available tryptophan content of an important food source. Pellagra is a disorder caused by niacin (vitamin B3) deficiency, usually owing to poverty and a diet relying heavily on corn (maize), which has a low level of niacin and its precursor tryptophan. Cultures in the Americas that relied greatly on corn used alkali during its processing (e.g., boiling the corn in lime when making tortillas). This enhanced the nutritional quality of the corn by increasing the bioavailability of both niacin and tryptophan, a practice that prevented pellagra 137. The Europeans transported corn around the world but did not transport the traditional alkali-processing methods, thereby causing epidemics of pellagra in past centuries. Breeding corn with a higher tryptophan content was shown in the 1980s to prevent pellagra 138; presumably, it also raised brain serotonin. In a recent issue of Nature Biotechnology, Morris and Sands 139 argue that plant breeders should be focusing more on nutrition than on yield. They ask, “Could consumption of tryptophan-rich foods play a role in reducing the prevalence of depression and aggression in society?” Cross-national studies have reported a positive association between corn consumption and homicide rates 140 and a negative association between dietary tryptophan and suicide rates 141. Although the idea behind such studies is interesting, any causal attribution must remain speculative, given the possible confounders. Nonetheless, the possibility that the mental health of a population could be improved by increasing the dietary intake of tryptophan relative to the dietary intake of other amino acids remains an interesting idea that should be explored.
Serotonin foods
The following fruits and nuts had a high serotonin concentration expressed in µg/g weight:
- Butternuts 398 ± 90 µg/g
- Black walnuts 304 ± 46 µg/g
- Shagbark hickory nuts 143 ± 23 µg/g
- English walnuts 87 ± 20 µg/g
- Mockemut hickory nuts 67 ± 13 µg/g
- Plantain 30.3 ± 7.5 µg/g
- Pecans 29 ± 4 µg/g
- Sweet pignuts 25 ± 8 µg/g
- Pineapple 17.0 ± 5.1 µg/g
- Banana 15.0 ± 2.4 µg/g
- Kiwi fruit 5.8 ± 0.9 µg/g
- Plums 4.7 ± 0.8 µg/g
- Tomatoes 3.2 ± 0.6 µg/g
Table 2. Foods high in tryptophan (ordered from highest to low)
| Description | Tryptophan (g) Value Per 100 gram of food source |
|---|---|
| Egg, white, dried, stabilized, glucose reduced | 1.43 |
| Egg, white, dried, powder, stabilized, glucose reduced | 1.27 |
| Egg, white, dried, flakes, stabilized, glucose reduced | 1.18 |
| Soy protein isolate | 1.12 |
| Soy protein isolate, potassium type | 1.12 |
| Seeds, sesame flour, low-fat | 1.1 |
| Egg, white, dried | 1 |
| Seaweed, spirulina, dried | 0.93 |
| Seeds, sesame flour, partially defatted | 0.88 |
| Soy protein concentrate, produced by alcohol extraction | 0.83 |
| Soy protein concentrate, produced by acid wash | 0.83 |
| Whale, beluga, meat, dried (Alaska Native) | 0.8 |
| Egg, whole, dried | 0.78 |
| Egg, whole, dried, stabilized, glucose reduced | 0.77 |
| Winged beans, mature seeds, raw | 0.76 |
| Seeds, cottonseed flour, low fat (glandless) | 0.75 |
| Tofu, dried-frozen (koyadofu) | 0.75 |
| Tofu, dried-frozen (koyadofu), prepared with calcium sulfate | 0.75 |
| Seeds, cottonseed meal, partially defatted (glandless) | 0.74 |
| Seeds, sunflower seed flour, partially defatted | 0.73 |
| Beverages, Protein powder soy based | 0.72 |
| Fish, cod, Atlantic, dried and salted | 0.7 |
| Soy flour, defatted | 0.68 |
| Seeds, sesame flour, high-fat | 0.67 |
| Soy meal, defatted, raw | 0.65 |
| Pork, fresh, variety meats and by-products, pancreas, cooked, braised | 0.62 |
| Seeds, cottonseed flour, partially defatted (glandless) | 0.62 |
| Mollusks, whelk, unspecified, cooked, moist heat | 0.62 |
| Soybeans, mature seeds, raw | 0.59 |
| Seeds, pumpkin and squash seed kernels, dried | 0.58 |
| Soybeans, mature seeds, dry roasted | 0.57 |
| Meat extender | 0.57 |
| Seeds, pumpkin and squash seed kernels, roasted, without salt | 0.57 |
| Seeds, pumpkin and squash seed kernels, roasted, with salt added | 0.57 |
| Cheese, parmesan, shredded | 0.56 |
| Cheese, mozzarella, low moisture, part-skim | 0.55 |
| Cheese, cheddar (Includes foods for USDA’s Food Distribution Program) | 0.55 |
| Game meat, elk, cooked, roasted | 0.55 |
| Leavening agents, yeast, baker’s, active dry | 0.54 |
| Parsley, freeze-dried | 0.52 |
| Cheese, mozzarella, whole milk | 0.52 |
| Soybeans, mature seeds, roasted, salted | 0.51 |
| Soybeans, mature seeds, roasted, no salt added | 0.51 |
| Milk, dry, nonfat, regular, without added vitamin A and vitamin D | 0.51 |
| Milk, dry, nonfat, regular, with added vitamin A and vitamin D | 0.51 |
| Peanut flour, defatted | 0.51 |
| Soy flour, full-fat, roasted | 0.51 |
| Soy flour, full-fat, raw | 0.5 |
| Milk, dry, nonfat, calcium reduced | 0.5 |
| Milk, dry, nonfat, instant, with added vitamin A and vitamin D | 0.49 |
| Milk, dry, nonfat, instant, without added vitamin A and vitamin D | 0.49 |
| Seeds, cottonseed kernels, roasted (glandless) | 0.49 |
| Milk, buttermilk, dried | 0.48 |
| Cheese, parmesan, hard | 0.48 |
| Spices, parsley, dried | 0.47 |
| Pork, cured, bacon, cooked, microwaved | 0.46 |
| Game meat, caribou, cooked, roasted | 0.46 |
| Seeds, chia seeds, dried | 0.44 |
| Game meat, rabbit, wild, cooked, stewed | 0.44 |
| Cheese, romano | 0.43 |
| Cheese, gruyere | 0.42 |
| Lamb, shoulder, arm, separable lean only, trimmed to 1/4″ fat, choice, cooked, braised | 0.41 |
| T.G.I. FRIDAY’S, classic sirloin steak (10 oz) | 0.41 |
| Game meat, elk, raw | 0.41 |
| CRACKER BARREL, grilled sirloin steak | 0.41 |
| Pork, ground, 96% lean / 4% fat, cooked, pan-broiled | 0.41 |
| Beef, round, top round roast, boneless, separable lean only, trimmed to 0″ fat, select, cooked, roasted | 0.41 |
| Pork, fresh, variety meats and by-products, pancreas, raw | 0.41 |
| Pork, cured, bacon, pre-sliced, cooked, pan-fried | 0.41 |
| Beef, round, eye of round roast, boneless, separable lean only, trimmed to 0″ fat, select, cooked, roasted | 0.41 |
| Beef, round, top round, separable lean only, trimmed to 0″ fat, choice, cooked, braised | 0.41 |
| Beef, round, top round, separable lean only, trimmed to 0″ fat, select, cooked, braised | 0.41 |
| Chicken, broiler or fryers, breast, skinless, boneless, meat only, cooked, braised | 0.4 |
| Goose, domesticated, meat only, cooked, roasted | 0.4 |
| Seeds, safflower seed meal, partially defatted | 0.4 |
| Game meat, goat, cooked, roasted | 0.4 |
| Beef, round, top round steak, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, grilled | 0.4 |
| Cheese, swiss | 0.4 |
| Game meat, rabbit, domesticated, composite of cuts, cooked, stewed | 0.4 |
| Beef, loin, top sirloin filet, boneless, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 0.4 |
| Duck, young duckling, domesticated, White Pekin, leg, meat only, bone in, cooked without skin, braised | 0.4 |
| Beef, plate steak, boneless, inside skirt, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 0.4 |
| Beef, round, top round, separable lean and fat, trimmed to 0″ fat, choice, cooked, braised | 0.4 |
| Beef, round, top round, separable lean and fat, trimmed to 0″ fat, select, cooked, braised | 0.4 |
| Lamb, Australian, imported, fresh, shoulder, arm, separable lean only, trimmed to 1/8″ fat, cooked, braised | 0.4 |
| Cereals ready-to-eat, wheat germ, toasted, plain | 0.4 |
| Lamb, New Zealand, imported, frozen, shoulder, whole (arm and blade), separable lean only, cooked, braised | 0.4 |
| Seeds, sesame butter, paste | 0.4 |
| Pork, ground, 96% lean / 4% fat, cooked, crumbles | 0.39 |
| Beef, round, top round steak, boneless, separable lean only, trimmed to 0″ fat, choice, cooked, grilled | 0.39 |
| Lamb, cubed for stew or kabob (leg and shoulder), separable lean only, trimmed to 1/4″ fat, cooked, braised | 0.39 |
| Seeds, sesame butter, tahini, from unroasted kernels (non-chemically removed seed coat) | 0.39 |
| Restaurant, family style, sirloin steak | 0.39 |
| Spices, fenugreek seed | 0.39 |
| Egg, yolk, dried | 0.39 |
| Chicken, broilers or fryers, breast, meat only, cooked, fried | 0.39 |
| Seeds, watermelon seed kernels, dried | 0.39 |
| Seeds, sesame butter, tahini, from raw and stone ground kernels | 0.39 |
| Beef, top loin filet, boneless, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 0.39 |
| Seeds, sesame seeds, whole, dried | 0.39 |
| Beef, round, top round, separable lean and fat, trimmed to 1/8″ fat, select, cooked, braised | 0.39 |
| Beef, loin, top loin steak, boneless, lip-on, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 0.39 |
| Chicken, stewing, light meat, meat only, cooked, stewed | 0.39 |
| Pork, fresh, loin, tenderloin, separable lean only, cooked, broiled | 0.39 |
| Beef, loin, top loin steak, boneless, lip off, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 0.39 |
| Chicken, broiler or fryers, breast, skinless, boneless, meat only, cooked, grilled | 0.39 |
| Beef, round, top round, separable lean and fat, trimmed to 1/8″ fat, all grades, cooked, braised | 0.39 |
| Beef, chuck, mock tender steak, boneless, separable lean only, trimmed to 0″ fat, choice, cooked, braised | 0.39 |
| Beef, round, eye of round steak, boneless, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 0.39 |
| Game meat, rabbit, domesticated, composite of cuts, cooked, roasted | 0.38 |
| Cheese, parmesan, grated | 0.38 |
| Chicken, broilers or fryers, light meat, meat only, cooked, fried | 0.38 |
| Lamb, shoulder, whole (arm and blade), separable lean only, trimmed to 1/4″ fat, choice, cooked, braised | 0.38 |
| Beef, loin, top sirloin petite roast, boneless, separable lean only, trimmed to 0″ fat, select, cooked, roasted | 0.38 |
| Beef, round, top round, separable lean and fat, trimmed to 1/8″ fat, choice, cooked, braised | 0.38 |
| Beef, chuck, mock tender steak, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, braised | 0.38 |
| Beef, rib, back ribs, bone-in, separable lean only, trimmed to 0″ fat, select, cooked, braised | 0.38 |
| Beef, round, top round roast, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, roasted | 0.38 |
| Duck, young duckling, domesticated, White Pekin, breast, meat only, boneless, cooked without skin, broiled | 0.38 |
| Game meat, boar, wild, cooked, roasted | 0.38 |
| DENNY’S, top sirloin steak | 0.38 |
| Lamb, shoulder, blade, separable lean only, trimmed to 1/4″ fat, choice, cooked, braised | 0.38 |
| Beef, chuck, mock tender steak, boneless, separable lean only, trimmed to 0″ fat, select, cooked, braised | 0.38 |
| Beef, rib eye steak, boneless, lip-on, separable lean only, trimmed to 1/8″ fat, select, cooked, grilled | 0.38 |
| Beef, rib eye steak, boneless, lip off, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 0.38 |
| Beef, shank crosscuts, separable lean only, trimmed to 1/4″ fat, choice, cooked, simmered | 0.38 |
| Beef, round, eye of round roast, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, roasted | 0.38 |
| Pork, fresh, loin, tenderloin, separable lean and fat, cooked, broiled | 0.38 |
| Snacks, soy chips or crisps, salted | 0.38 |
| Fish, roe, mixed species, cooked, dry heat | 0.38 |
| Beef, rib eye roast, boneless, lip-on, separable lean only, trimmed to 1/8″ fat, select, cooked, roasted | 0.38 |
| Beef, loin, top sirloin filet, boneless, separable lean only, trimmed to 0″ fat, all grades, cooked, grilled | 0.38 |
| Pork, fresh, leg (ham), whole, separable lean only, cooked, roasted | 0.37 |
| Pork, fresh, loin, center rib (chops), boneless, separable lean only, cooked, broiled | 0.37 |
| Pork, fresh, composite of trimmed retail cuts (loin and shoulder blade), separable lean only, cooked | 0.37 |
| Beef, plate steak, boneless, outside skirt, separable lean only, trimmed to 0″ fat, select, cooked, grilled | 0.37 |
| Chicken, broilers or fryers, giblets, cooked, fried | 0.37 |
| Beef, chuck for stew, separable lean and fat, choice, cooked, braised | 0.37 |
| Turkey, retail parts, wing, meat only, cooked, roasted | 0.37 |
| Chicken, broiler or fryers, breast, skinless, boneless, meat only, with added solution, cooked, grilled | 0.37 |
| Ham and cheese spread | 0.37 |
| Seeds, sesame butter, tahini, from roasted and toasted kernels (most common type) | 0.37 |
| Veal, leg (top round), separable lean only, cooked, braised | 0.37 |
| Beef, chuck for stew, separable lean and fat, all grades, cooked, braised | 0.37 |
| Milk, dry, whole, with added vitamin D | 0.37 |
| Milk, dry, whole, without added vitamin D | 0.37 |
| Seeds, sesame seeds, whole, roasted and toasted | 0.37 |
| Seeds, sesame seed kernels, toasted, without salt added (decorticated) | 0.37 |
| Seeds, sesame meal, partially defatted | 0.37 |
| Seeds, sesame seed kernels, toasted, with salt added (decorticated) | 0.37 |
Table 3. L-tryptophan found in common foods
| L-tryptophan*(mg) | Sum of CAAs** (mg) | Ratio | |
|---|---|---|---|
| Turkey, Skinless, Boneless, Light Meat (per pound, raw) | 410 | 9525 | 0.043 |
| Chicken, Skinless, Boneless, Light Meat (per pound, raw) | 238 | 5122 | 0.046 |
| Turkey, Skinless, Boneless, Dark Meat (per pound, raw) | 303 | 7036 | 0.043 |
| Chicken, Skinless, Boneless, Dark Meat (per pound, raw) | 256 | 5492 | 0.047 |
| Whole Milk (per quart) | 732 | 8989 | 0.081 |
| 2% Milk (per quart) | 551 | 12516 | 0.044 |
| Wheat Bread (per slice) | 19 | 317 | 0.06 |
| White Bread (per slice) | 22 | 439 | 0.05 |
| Semisweet Chocolate (per ounce) | 18 | 294 | 0.061 |
| Sweet Chocolate (per ounce) | 16 | 270 | 0.059 |
| Canned Tuna (per ounce) | 472 | 10591 | 0.045 |
| Cheddar Cheese (per ounce) | 91 | 2298 | 0.04 |
| Peanuts (per ounce) | 65 | 1574 | 0.041 |
| Oats for Oatmeal (per cup) | 147 | 2617 | 0.056 |
| Dried Prune (one) | 2 | 27 | 0.074 |
| Banana (one medium) | 11 | 237 | 0.046 |
| Apple (one medium) | 2 | 70 | 0.029 |
Footnotes: The L-tryptophan/competing amino acid (CAA) ratio represents the relative availability of plasma L-tryptophan for crossing the blood-brain barrier and is thought to be the best indicator of brain serotonin synthesis.
*e.g. The recommended daily allowance for a 79 kg (175 lb) adult is 278 to 476 mg.
**CAAs = Isoleucine, Leucine, Phenylalanine, Tyrosine, and Valine, the five large neutral amino acids typically included in the tryptophan/competing amino acid (CAA) ratio.
Selective Serotonin Reuptake Inhibitors (SSRIs)
SSRIs is short for selective serotonin reuptake inhibitors, which are a class of antidepressant medications that works by increasing serotonin levels in your brain, that are commonly prescribed to treat depression, anxiety disorders, obsessive-compulsive disorder (OCD), panic disorders, and eating disorder bulimia nervosa and other mental health conditions 144, 145. Selective serotonin reuptake inhibitors (SSRIs) work by blocking the reabsorption (reuptake) of serotonin, a neurotransmitter (a chemical messenger) in your brain, which increases the amount of serotonin available to nerve cells (neurons) in the brain which is associated with their antidepressant effect. Serotonin also known as 5-hydroxytryptamine or 5-HT, is one of the a biochemical messengers and regulators, synthesized from the essential amino acid L-Tryptophan that nerve cells (neurons) in your brain use to communicate 1, 2, 3. Raising serotonin levels can help regulate mood, appetite, digestion, sleep, and many other bodily functions. Selective serotonin reuptake inhibitors (SSRIs) include drugs such as citalopram, escitalopram, fluoxetine, paroxetine, and sertraline.
SSRIs drugs include:
- Citalopram (Celexa)
- Escitalopram (Lexapro)
- Fluoxetine (Prozac®, Prozac Weekly®, Rapuflux®, Selfemra®, and Sarafem®)
- Paroxetine (Paxil®, Aropax®, Brisdelle®, Pexeva® and Seroxat®)
- Sertraline (Zoloft)
- Vilazodone (Viibryd)
SSRIs common uses:
- Major depressive disorder
- Anxiety disorders, including generalized anxiety disorder (GAD) and panic disorder
- Severe phobias such as agoraphobia and social anxiety disorder (social phobia)
- Obsessive-compulsive disorder (OCD)
- Post-traumatic stress disorder (PTSD)
- Bulimia nervosa
- Bipolar depression 146
SSRIs can sometimes be used to treat other conditions, such as:
SSRIs are only available orally and come in multiple forms, including tablets, capsules, or liquid suspension/solution. There are currently no parenteral (IV, IM, SubQ), rectal, or other forms of SSRIs. SSRI administration is typically once-daily medication in the morning or nighttime. Except for vilazodone, SSRIs may be taken without regard to food. Take vilazodone with food 147.
SSRIs are usually taken in tablet form. When they’re prescribed, you’ll start on the lowest possible dose thought necessary to improve your symptoms. SSRIs usually need to be taken for 2 to 4 weeks before the benefit is felt. You may experience mild side effects early on, but it’s important that you don’t stop taking the medicine. These effects will usually wear off quickly.
If you take an SSRI for 4 to 6 weeks without feeling any benefit, speak to your doctor or mental health specialist. Your doctor or mental health specialist may recommend increasing your dose or trying an alternative antidepressant. This is because SSRIs differ in how well they block serotonin reuptake and in how quickly they break down and are cleared from the body.
An SSRI that is best for you depends on several factors, such as your symptoms and any other health conditions you may have.
A course of treatment usually continues for at least 6 months after you feel better, although longer courses are sometimes recommended and some people with recurrent problems may be advised to take them indefinitely.
SSRIs also need to be used with caution if you have certain underlying health problems, including diabetes, epilepsy and kidney disease.
Some SSRIs can react unpredictably with other medicines, including some over-the-counter painkillers and herbal remedies, such as St John’s wort. Always read the information leaflet that comes with your SSRI medicine to check if there are any medicines you need to avoid.
SSRIs are generally considered safe and effective, but they can cause side effects, including:
- Upset stomach, vomiting or diarrhea.
- Insomnia
- Headaches
- Sexual dysfunction
- Sweating
- Sleepiness or trouble sleeping.
- Dry mouth.
- Nervousness, anxiety or restlessness.
- Shakiness.
- Sexual problems, such as lessened sexual desire, trouble reaching orgasm or trouble getting and keeping an erection.
- Changes in appetite, leading to weight loss or weight gain.
Taking your SSRI with food may lessen the risk of an upset stomach. Also, so long as your SSRI medicien doesn’t keep you from sleeping, taking it at bedtime may lessen an upset stomach.
SSRIs are generally safe for most people. But some can cause safety issues. For example, citalopram can cause dangerous irregular heart rhythms if the dose is too high. The U.S. Food and Drug Administration (FDA) and the manufacturer recommend that the dose should be no more than 40 milligrams (mg) a day, but no more than 20 mg of citalopram a day for people over age 60.
Before you take an SSRI, talk with your doctor about:
- Interactions with other medicines and supplements. When taking an antidepressant, tell your doctor about any other prescription or nonprescription medicines, herbs, or other supplements you’re taking. Some antidepressants can interfere with the effectiveness of other medicines. Some antidepressants can cause dangerous reactions when combined with certain medicines or herbal supplements. For example, SSRIs may raise your risk of bleeding. The risk is higher when you also take other medicines that raise the risk of bleeding, for example, a nonsteroidal anti-inflammatory drug, such as aspirin or ibuprofen (Advil, Motrin IB, others), or warfarin (Jantoven) and other blood thinners.
- Serotonin syndrome. Rarely, an antidepressant can cause high levels of serotonin to build up in your body. Serotonin syndrome most often occurs when two medicines that raise the level of serotonin are combined. These include, for example, other antidepressants, some pain or headache medicines, and the herbal supplement St. John’s wort. Symptoms of serotonin syndrome include anxiety, being nervous or jittery, high fever, sweating, confusion, shaking, restlessness, lack of coordination, major changes in blood pressure, and a fast heartbeat. Get medical help right away if you have any of these symptoms.
- Antidepressants and pregnancy. Talk to your doctor about the risks and benefits of using specific antidepressants. Some antidepressants may harm your baby if you take them during pregnancy or while you’re breastfeeding. If you’re taking an antidepressant and you’re thinking about getting pregnant, talk with your doctor about the possible risks. Don’t stop taking your medicine without talking with your doctor first, as stopping puts you at risk.
- Suicide risk and antidepressants. Most antidepressants are generally safe. But the U.S. Food and Drug Administration (FDA) requires that all antidepressants carry boxed warnings, the strictest warnings for prescriptions. In some cases, children, teenagers and young adults under 25 may have an increase in suicidal thoughts or behavior when taking antidepressants. This may be more likely in the first few weeks after starting or when the dose is changed. Anyone taking an antidepressant should be watched closely for worsening depression or unusual behavior. If you or someone you know has suicidal thoughts when taking an antidepressant, call your doctor right away or get emergency help. Keep in mind that depression that’s not treated is a more concerning risk of suicide. And antidepressants may lessen suicide risk in the long run by improving mood for many people.
- Stopping treatment with SSRIs. SSRIs aren’t habit-forming. However, stopping antidepressant treatment suddenly or missing several doses can cause withdrawal-like symptoms. This is sometimes called discontinuation syndrome. Work with your doctor to slowly and safely lower your dose.
- SSRI withdrawal-like symptoms can include:
- Restlessness and anxiety.
- Nausea.
- Dizziness.
- Feeling sluggish or sleepy.
- Flu-like symptoms, such as chills, sweating and muscle aches.
- SSRI withdrawal-like symptoms can include:
Finding the right antidepressant
Different people may react differently to the same antidepressant. For example, one medicine may work better — or not as well — for you than for another person. Or you may have more, or fewer, side effects from taking a specific antidepressant than someone else does.
Traits passed down in your family play a role in how antidepressants affect you. If a relative responded well to a particular antidepressant, tell your doctor. This may be a good medicine for you to try first.
In some cases, results of special blood tests, where available, may offer clues about how your body may respond to a particular antidepressant. But other factors can affect your response to medicine. For example, people with a history of bipolar disorder typically aren’t given SSRIs for depression. That’s because SSRIs may worsen their symptoms.
When choosing an antidepressant, your doctor considers your symptoms, any health conditions you may have, other medicines you take and what has worked for you in the past.
It may take several weeks or more before an antidepressant is fully effective and for early side effects to ease up. Your doctor may recommend some dose changes or different antidepressants. With patience, you and your doctor can find a medicine that works well for you.
How selective serotonin reuptake inhibitor (SSRI) drugs work
The therapeutic actions of selective serotonin reuptake inhibitors (SSRIs) have their basis on increasing serotonin that researchers hypothesize as the cause of depression in the monoamine hypothesis 144. Serotonin is a neurotransmitter (a messenger chemical that carries signals between nerve cells in the brain). It’s thought to have a good influence on mood, emotion and sleep. After carrying a message, serotonin is usually reabsorbed by the nerve cells (known as “reuptake”). Selective serotonin reuptake inhibitors (SSRIs) work by blocking (“inhibiting”) the reuptake of serotonin in the brain, thereby increasing serotonin activity, meaning more serotonin is available to pass further messages between nearby nerve cells. SSRIs inhibit the serotonin transporter (SERT) at the presynaptic axon terminal. By inhibiting serotonin transporter (SERT), an increased amount of serotonin (5-hydroxytryptamine or 5HT) remains in the synaptic cleft and can stimulate postsynaptic receptors for a more extended period 148, 149, 150, 147. However, it would be too simplistic to say that depression and related mental health conditions are caused by low serotonin levels, but a rise in serotonin levels can improve symptoms and make people more responsive to other types of treatment, such as Cognitive Behavioral Therapy (CBT).
Unlike other classes of antidepressants, SSRIs have little effect on other neurotransmitters, such as dopamine or norepinephrine. SSRIs also have relatively fewer side effects than tricyclic antidepressants (TCAs) and monoamine oxidase inhibitors (MAOIs) due to fewer effects on adrenergic, cholinergic, and histaminergic receptors.
Selective serotonin reuptake inhibitor (SSRI) drugs uses
Selective serotonin reuptake inhibitors (SSRIs) are a widely used type of antidepressant (medicine used to treat clinical depression). SSRIs are usually the first choice medicine for depression because they generally have fewer side effects than most other types of antidepressant 144. SSRIs are mainly prescribed to treat depression, particularly persistent or severe cases, and are often used in combination with a talking therapy such as cognitive behavioral therapy (CBT).
Selective serotonin reuptake inhibitors (SSRIs) are approved for use in both adult and children 151, 152
SSRIs in use in the United States are:
- Fluoxetine
- Sertraline
- Paroxetine
- Fluvoxamine
- Citalopram
- Escitalopram
- Vilazodone
SSRIs currently with FDA labeled indications to treat the following conditions 144:
- Major depressive disorder
- Generalized anxiety disorder (GAD)
- Bulimia nervosa
- Bipolar depression 146
- Obsessive-compulsive disorder (OCD)
- Panic disorder
- Premenstrual dysphoric disorder (PMDD)
- Treatment-resistant depression
- Post-traumatic stress disorder (PTSD)
- Social anxiety disorder (social phobia)
- Severe phobias such as agoraphobia
Other off-label uses of SSRI include but are not limited to 153, 144:
- Binge eating disorder
- Body dysmorphic disorder
- Fibromyalgia
- Premature ejaculation
- Paraphilias: Paraphilia is an experience of recurring or intense sexual arousal to atypical objects, situations, fantasies, behaviors and/or targets (eg, children, corpses, animals) 154. Paraphilia has also been defined as a sexual interest in anything other than a legally consenting human partner. A total of eight Paraphilias are listed in the DSM V and include pedophilia, exhibitionism, voyeurism, sexual sadism, sexual masochism, frotteurism, fetishism, and transvestic fetishism 155. Although not innately pathological, a paraphilic disorder can evolve if paraphilia invokes harm, distress, or functional impairment on the lives of the affected individual or others.
- Pedophilia is any sexual activity with a prepubescent child, where the offender/patient is at least sixteen years of age, and the victim is at least five years younger 156.
- Exhibitionism is the exposure of an individual’s genitalia to unsuspecting strangers for sexual satisfaction 157
- Voyeurism is the viewing of an unsuspecting person engaging in disrobing or sexual activity 158.
- Sexual sadism is when sexual arousal is gained from inflicting mental or physical suffering on a nonconsenting person 159.
- Sexual masochism is the derivation of sexual arousal from being the recipient of physical or mental abuse and/or humiliation 160.
- Frotteurism is the touching of or rubbing against a nonconsenting person 161.
- Fetishism is the use of nonliving objects, most commonly shoes and undergarments, for sexual pleasure 162.
- Transvestic fetishism is the derivation of sexual arousal from cross-dressing or dressing in clothes of the opposite sex 163.
- Autism spectrum disorder (ASD) is a developmental disability that causes a complex range of behavioural symptoms.
- Raynaud phenomenon also called Raynaud’s disease is a condition that causes blood vessels in the extremities, particularly fingers and toes, to narrow in response to cold or stress, leading to reduced blood flow and a characteristic color change (white, then blue, then red)
- Vasomotor symptoms (hot flushes) associated with menopause.
SSRI Contraindications
SSRIs are contraindicated with the concurrent use of monoamine oxidase inhibitors (MAOIs), linezolid, and other medications that increase serotonin levels and could put patients at risk for life-threatening serotonin syndrome. Do not use SSRI if you have used an monoamine oxidase (MAO) inhibitor in the past 14 days. A dangerous drug interaction could occur. Monoamine oxidase (MAO) inhibitors include isocarboxazid (Marplan), linezolid (Zyvox), methylene blue, rasagiline, phenelzine (Nardil), selegiline (Emsam, Zelapar), and tranylcypromine (Parnate). Wait at least 14 days after stopping an monoamine oxidase (MAO) inhibitor before you take SSRI.
Paroxetine is contraindicated in pregnancy and is classified as category D/X due to its teratogenic effects in causing cardiovascular defects, specifically heart malformations if prescribed in the first trimester 164.
Pregnancy and SSRI
SSRIs can be prescribed if you’re pregnant, breastfeeding or under 18, but this depends if your doctor thinks that the benefits outweigh the risks. Talk to your doctor about what’s best for you or your child.
Babies of women who are taking an SSRI at the time of delivery may have some difficulties in the first few days of life. Reported symptoms include jitteriness, increased muscle tone, irritability, constant crying, changes in sleeping patterns, tremors, difficulty eating, and problems with breathing. Not every baby will experience these symptoms. For the SSRI medications, it is estimated that 10-30% of babies will be affected.
Some babies with symptoms of withdrawal may need to spend time in the neonatal intensive care unit (NICU) to receive additional care. However, in most cases the symptoms are mild and go away within two weeks. Reassuringly, there does not seem to be a dose-response relationship, which means that women who need a higher amount of medication to manage their anxiety are not expected to have babies who are at a higher risk for withdrawal.
SSRI side effects
Most people will only experience a few mild side effects when taking SSRIs. These can be troublesome at first, but they’ll generally improve with time 165, 166.
Common side effects of SSRIs can include 167, 168:
- feeling agitated, shaky or anxious
- diarrhea and feeling or being sick
- gastrointestinal distress
- dizziness
- blurred vision
- headache
- loss of libido (reduced sex drive)
- difficulty achieving orgasm during sex or masturbation
- in men, difficulty obtaining or maintaining an erection (erectile dysfunction)
- sleep disturbances
- weight changes
- xerostomia
SSRIs also have the potential to prolong the QT interval, which can lead to fatal arrhythmia, torsade de pointes. Citalopram has correlations with a longer QT duration, or QT prolongation than the other medications in this class 169, 170. Coagulopathy (bleeding disorder) also correlates with SSRI use 171. Although infrequent, as with all medications that increase serotonin activity, it is important to be aware of the risk of serotonin syndrome, particularly when prescribing multiple medications that may have serotonergic effects.
In 2004, the FDA issued a black box warning for SSRIs and other antidepressant medications due to a possible increased risk of suicidality among pediatric and young adult (up to age 25) populations 172, 173. The risk and benefits of initiating SSRI therapy on acutely suicidal patients must be weighed, keeping in mind that depression itself is a large risk factor for the risk of suicide (usually indicated by suicidal ideation or intent) and requires treatment.
SSRIs are brokendown (metabolized) by and have effects on the cytochrome P450 system. Fluoxetine, paroxetine, sertraline, citalopram, and escitalopram are inhibitors of CYP2D6. Fluoxetine and fluvoxamine are inhibitors of CYP2C19. Fluvoxamine is an inhibitor of CYP1A2 147.
You’ll usually need to see your doctor every few weeks when you first start taking SSRIs to discuss how well the medicine is working. You can also contact your doctor at any point if you experience any troublesome or persistent side effects.
SSRI Overdose
Symptoms of SSRI (selective serotonin reuptake inhibitors) overdose may include the following:
- dizziness
- drowsiness
- sweating
- agitation, hallucinations, fever, sweating, confusion, fast heartbeat, shivering, severe muscle stiffness or twitching, loss of coordination, nausea, vomiting, or diarrhea
- nausea
- vomiting
- uncontrollable shaking of a part of the body
- drowsiness
- fast, irregular, or pounding heartbeat
- memory loss
- confusion
- seizures
- coma (loss of consciousness)
- fast breathing
- bluish color around mouth, fingers, or fingernails
- muscle pain
- dark-colored urine.
Out of all the SSRIs, citalopram and escitalopram are more likely to cause overdose due to differences in their structures. Citalopram and escitalopram have an increased risk of heart toxicity due to QT prolongation, which can progress to serious arrhythmias such as Torsades de pointes, a type of very fast heart rhythm (tachycardia) that starts in your heart’s lower chambers (ventricles).
Serotonin syndrome
Serotonin syndrome also called serotonin toxicity is a potentially life threatening condition due to excessive serotonin in your body 174, 175. Serotonin syndrome most often occurs when two medicines that affect the body’s level of serotonin are taken together at the same time. The medicines cause too much serotonin to be released or to remain in the brain area. Serotonin syndrome (serotonin toxicity) can occur when you increase the dose of such a drug or add a new drug to your regimen. Certain illegal drugs and dietary supplements also are associated with serotonin syndrome. For example, you can develop serotonin syndrome if you take migraine medicines called triptans together with antidepressants called selective serotonin reuptake inhibitors (SSRIs), and selective serotonin/norepinephrine reuptake inhibitors (SSNRIs).
Serotonin syndrome symptoms and their severity vary from person to person. They can be mild to severe to even fatal.
If you’re taking a medication that affects serotonin and experience any of the following symptoms, call your doctor or visit an urgent or emergency care facility right away.
Mild symptoms
- Nervousness.
- Nausea, vomiting.
- Diarrhea.
- Dilated pupils.
- Tremor.
Moderate symptoms
- Agitation, restlessness.
- Muscle twitching, involuntary muscle contractions, muscle spasms, muscle rigidity.
- Sweating, shivering.
- Abnormal (side-to-side) eye movements.
Severe symptoms
- Confusion, disorientation, delirium.
- Rapid heart rate.
- High blood pressure.
- High body temperature (greater than 101.3° Fahrenheit [38.5° Celsius]).
- Seizures.
- Abnormal heartbeat.
- Passing out, fainting.
Symptoms usually begin within a few hours of taking a new medication that affects serotonin levels or increasing the dose of a drug you’re already taking. Nearly all people will experience symptoms within 24 hours of starting, adding or increasing the dosage of a serotonergic medication or product.
Serotonin syndrome generally doesn’t cause any problems once serotonin levels are back to their original levels.
Without treatment, serotonin symptoms can cause:
- Seizures.
- Trouble breathing.
- Kidney failure. The breakdown products of muscle (from spasms) are released into your blood, which is filtered through your kidneys. If left untreated, these products can cause severe kidney damage.
- Coma.
- Death.
If left untreated, severe serotonin syndrome can lead to unconsciousness and death.
There aren’t any tests to diagnose serotonin syndrome symptoms. Your doctor usually makes the diagnosis based on the results of your physical exam, review of your symptoms and history of medications you take that affect serotonin levels.
You can assist your doctor by telling them all the products you take, including prescription medicines, over-the-counter medicines, supplements, herbal products and illegal drugs. Be honest. Don’t worry. Your doctor is here to help you, not judge you.
To make sure your symptoms are caused by serotonin syndrome and not due to another cause, your doctor may use tests to:
- Measure levels of any drugs you’re using
- Check for signs of infection
- Check body functions that may be affected by serotonin syndrome
A number of conditions can cause symptoms similar to those of serotonin syndrome. Minor symptoms can be caused by several conditions. Moderate and severe symptoms similar to those of serotonin syndrome could be caused by:
- A serious reaction to certain medications, such as some anesthetics, antipsychotic drugs and other agents known to produce these severe reactions
- An overdose of illicit drugs, antidepressant medications or other medications that increase serotonin levels
- Damage associated with illicit drug use
- Severe alcohol withdrawal
Your doctor may order additional tests to rule out other causes of your symptoms. Tests may include:
- Blood and urine tests
- Chest X-ray
- Computerized Tomography (CT) scan
- Spinal tap (lumbar puncture)
Treatment of serotonin syndrome depends on the severity of your symptoms.
- If your symptoms are minor, a visit to the doctor and stopping the medication causing the problem may be enough.
- If you have symptoms that concern your doctor, you may need to go to the hospital. Your doctor may have you stay in the hospital for several hours to make sure your symptoms are improving.
- If you have severe serotonin syndrome, you’ll need intensive treatment in a hospital.
Depending on your symptoms, you may receive the following treatments:
- Muscle relaxants. Benzodiazepines, such as diazepam (Valium, Diastat) or lorazepam (Ativan), can help control agitation, seizures and muscle stiffness.
- Serotonin antagonists. If other treatments aren’t working, serotonin antagonists such as the medicine cyproheptadine may help. These medicines work by blocking certain serotonin receptors, lessening the activity that causes the symptoms.
- Oxygen and intravenous (IV) fluids. Breathing oxygen through a mask helps maintain oxygen levels in your blood, and IV fluids are used to treat dehydration and fever.
- Drugs that control heart rate and blood pressure. These may include esmolol (Brevibloc) or nitroprusside (Nitropress) to reduce a high heart rate or high blood pressure. If your blood pressure is too low, your doctor may give you phenylephrine (Vazculep) or epinephrine (Adrenalin, Epipen, others).
- A breathing tube and machine and medication to paralyze your muscles. You may need this treatment if you have a high fever (106° Fahrenheit [41.1° Celsius]).
Milder forms of serotonin syndrome usually go away within 24 to 72 hours of stopping medications that increase serotonin. You may need to take medications to block the effects of serotonin already in your system.
However, symptoms of serotonin syndrome caused by some antidepressants could take several weeks to go away completely. These medications remain in your system longer than do other medications that can cause serotonin syndrome.
Citalopram
Citalopram belongs to the class of antidepressants known as selective serotonin reuptake inhibitors (SSRIs) 176, 177, 178. Citalopram works by increasing the amount of serotonin, a natural substance in the brain that helps maintain mental balance. Citalopram is a medication that has been used to treat depression, including major depressive disorder. Citalopram is also sometimes used to treat obsessive-compulsive disorder [OCD] (with bothersome thoughts that won’t go away and the need to perform certain actions over and over), eating disorders, alcoholism, panic disorder (condition that causes sudden attacks of extreme fear with no apparent cause), premenstrual dysphoric disorder [PMDD] (a group of physical and emotional symptoms that occur before the menstrual period each month), social anxiety disorder (extreme fear of interacting with others or performing in front of others that interferes with normal life), post traumatic stress disorder (PTSD), tingling in the hands and feet caused by diabetes nerve damage (diabetic neuropathy), and certain male sexual problems. Citalopram may be prescribed for other uses; ask your doctor for more information.
Citalopram was approved for use in the United States in 1998 and Citalopram has become one of the most widely used antidepressant medications, with more than 16 million prescriptions being written yearly 178.
Citalopram is available as tablets of 10, 20 and 40 mg and in an oral solution (liquid) of 10 mg/5 mL in several generic forms and under the brand name of Celexa. Citalopram is usually taken once a day, in the morning or in the evening, with or without food. The recommended dosage of citalopram in adults is 20 mg once daily, increasing to 40 mg daily if necessary. Take citalopram at around the same time every day. Your doctor may start you on a low dose of citalopram and gradually increase your dose, not more often than once a week. Follow the directions on your prescription label carefully, and ask your doctor or pharmacist to explain any part you do not understand. Take citalopram exactly as directed. Do not take more or less of it or take it more often than prescribed by your doctor.
Your doctor may order certain laboratory tests and electrocardiograms (ECG; a test to monitor your heart rate and rhythm) before you start taking citalopram and during your treatment with citalopram.
It may take 1 to 4 weeks before you notice the full benefit of citalopram. Continue to take citalopram even if you feel well. Do not stop taking citalopram without talking to your doctor. Your doctor will probably decrease your dose gradually. If you suddenly stop taking citalopram, you may experience withdrawal symptoms such as mood changes, irritability, agitation, dizziness, numbness, tingling or electric shock-like sensations in the hands or feet, anxiety, confusion, headache, tiredness, nausea, sweating, shaking, frenzied or abnormally excited mood, and difficulty falling asleep or staying asleep. Tell your doctor if you experience any of these symptoms while you are decreasing your dose of citalopram or soon after you stop taking citalopram.
Common side effects of Citalopram are drowsiness, dyspepsia, nausea, headache, increased sweating and sexual dysfunction. Rare, but potentially severe adverse events include suicidal ideation and behavior, prolongation of the QTc interval, serotonin syndrome, precipitation of acute mania and acute glaucoma.
A small number of children, teenagers, and young adults (up to 24 years of age) who took antidepressants (‘mood elevators’) such as citalopram during clinical studies became suicidal (thinking about harming or killing oneself or planning or trying to do so). Children, teenagers, and young adults who take antidepressants to treat depression or other mental illnesses may be more likely to become suicidal than children, teenagers, and young adults who do not take antidepressants to treat these conditions. However, experts are not sure about how great this risk is and how much it should be considered in deciding whether a child or teenager should take an antidepressant. Children younger than 18 years of age should not normally take citalopram, but in some cases, a doctor may decide that citalopram is the best medication to treat a child’s condition.
You should know that your mental health may change in unexpected ways when you take citalopram or other antidepressants even if you are an adult over 24 years of age. You may become suicidal, especially at the beginning of your treatment and any time that your dose is increased or decreased. You, your family, or your caregiver should call your doctor right away if you experience any of the following symptoms: new or worsening depression; thinking about harming or killing yourself, or planning or trying to do so; extreme worry; agitation; panic attacks; difficulty falling asleep or staying asleep; aggressive behavior; irritability; acting without thinking; severe restlessness; and frenzied abnormal excitement. Be sure that your family or caregiver knows which symptoms may be serious so they can call the doctor if you are unable to seek treatment on your own.
Your doctor will want to see you often while you are taking citalopram, especially at the beginning of your treatment. Be sure to keep all appointments for office visits with your doctor.
The doctor or pharmacist will give you the manufacturer’s patient information sheet (Medication Guide) when you begin treatment with citalopram. Read the information carefully and ask your doctor or pharmacist if you have any questions.
No matter your age, before you take an antidepressant, you, your parent, or your caregiver should talk to your doctor about the risks and benefits of treating your condition with an antidepressant or with other treatments. You should also talk about the risks and benefits of not treating your condition. You should know that having depression or another mental illness greatly increases the risk that you will become suicidal. This risk is higher if you or anyone in your family has or has ever had bipolar disorder (mood that changes from depressed to abnormally excited) or mania (frenzied, abnormally excited mood), or has thought about or attempted suicide. Talk to your doctor about your condition, symptoms, and personal and family medical history. You and your doctor will decide what type of treatment is right for you.
Pregnancy
It is not known if citalopram can make it harder for a woman to get pregnant 179. Studies in animals found that citalopram might cause some reduced fertility (ability to get pregnant) 179.
Miscarriage is common and can occur in any pregnancy for many different reasons. There have been some studies that suggested taking antidepressant medications might slightly increase the chance of miscarriage. However, a study on citalopram and escitalopram did not find an increased chance of miscarriage in women taking these medications 179.
Every pregnancy starts out with a 3-5% chance of having a birth defect. This is called the background risk. There are reports of more than 15,000 pregnancies exposed to citalopram or escitalopram. Overall, the available data do not suggest that citalopram or escitalopram increases the chance of birth defects. While some reports have suggested an increased chance of heart defects or other birth defects, most studies have not found an increased chance of birth defects 179.
Some studies suggest that taking citalopram or escitalopram throughout the pregnancy might increase the chance of pregnancy-related problems such as preterm birth (birth before 37 weeks of pregnancy) and low birth weight (weighing less than 5 pounds, 8 ounces [2500 grams] at birth). However, research has also shown that when depression is left untreated during pregnancy, there could be an increased chance of pregnancy complications. This makes it hard to know if it is the medication, untreated depression, or other factors that are increasing the chance for these problems.
Some, but not all, studies have suggested that when pregnant women take SSRIs such as escitalopram or citalopram during the second half of the pregnancy, their babies might have an higher chance for a serious lung condition called persistent pulmonary hypertension. Persistent pulmonary hypertension happens in 1 or 2 out of 1,000 births. Among the studies looking at this, the overall chance for pulmonary hypertension when an SSRI was used in pregnancy was less than 1/100 (less than 1%).
One small study followed eleven babies exposed to citalopram during pregnancy. At one year of age there was no difference in their development compared to children who were not exposed to citalopram. Most studies find no increase in attention deficit hyperactivity disorder (ADHD) in children exposed to SSRIs during pregnancy. Most studies also find that SSRIs do not appear to increase the chance of autism spectrum disorder (ASD) after considering the effects of maternal depression or other factors.
Some medications taken during pregnancy are associated with withdrawal symptoms in a newborn after delivery. If you are taking escitalopram or citalopram at the time of delivery, your baby might have irritability, jitteriness, tremors (shivering), constant crying, different sleep patterns, problems with eating and controlling body temperature, and some problems with breathing. In most cases, these symptoms are mild and go away within a couple weeks with no treatment. Some babies may need to stay in the nursery or neonatal intensive care unit (NICU) until the symptoms go away. Most babies exposed to escitalopram or citalopram in late pregnancy do not have these symptoms. It is important that your doctor know you are taking these medications so that if symptoms occur your baby can get the care that is best for them.
Breastfeeding
Several studies have shown that small amounts of citalopram are found in breast milk 179. There have been a few case reports of sleepiness and weight loss, but in most studies no harmful effects were seen in breastfed babies 179. Studies also showed no difference in the intellectual development of babies exposed to citalopram during breastfeeding. A nursing infant should be monitored for being very sleepy (hard to wake for feeds), poor feeding and poor weight gain, especially in younger, exclusively breastfed infants and when a combination of medications for mental health are used. Be sure to talk to your doctor about all your breastfeeding questions. An alternative to Citalopram should be prescribed or you should stop breastfeeding while using citalopram.
Citalopram Dosage
The dose of Citalopram will be different for different patients. Follow your doctor’s orders or the directions on the label. The following information includes only the average doses of Citalopram. If your dose is different, do not change it unless your doctor tells you to do so.
The amount of medicine that you take depends on the strength of the medicine. Also, the number of doses you take each day, the time allowed between doses, and the length of time you take the medicine depend on the medical problem for which you are using the medicine.
For oral dosage forms (capsules)
- For depression
- Adults: 30 milligrams (mg) once a day. Your doctor may adjust your dose as needed. Do not start treatment with citalopram capsules because the only available dose strength is 30 mg. Use another dosage form of citalopram (eg, solution or tablets) for starting treatment and adjusting doses other than 30 mg once a day.
- Older adults: Use is not recommended.
- Children: Use and dose must be determined by your doctor.
For oral dosage forms (solution or tablets)
- For depression
- Adults: At first, 20 milligrams (mg) once a day, taken either in the morning or evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 40 mg per day.
- Older adults: 20 mg once a day, taken either in the morning or evening.
- Children: Use and dose must be determined by your doctor.
Missed Dose
If you miss a dose of Citalopram, take it as soon as possible. However, if it is almost time for your next dose, skip the missed dose and go back to your regular dosing schedule. Do not double doses.
Citalopram side effects
Citalopram may cause side effects. Tell your doctor if any of these symptoms are severe or do not go away:
- nausea
- diarrhea
- constipation
- vomiting
- stomach pain
- heartburn
- decreased appetite
- weight loss
- increased sweating
- increased thirst
- frequent urination
- difficulty falling asleep or staying asleep
- drowsiness
- excessive tiredness
- yawning
- weakness
- uncontrollable shaking of a part of the body
- muscle or joint pain
- dry mouth
- sexual problems in males; decreased sex drive, inability to get or keep an erection, or delayed or absent ejaculation
- sexual problems in females; decreased sex drive, or delayed orgasm or inability to have an orgasm
- heavy menstrual periods
- runny nose
Some side effects can be serious. If you experience any of the following symptoms, or those listed in the IMPORTANT WARNING or SPECIAL PRECAUTIONS sections, see your doctor immediately or get emergency medical treatment:
- chest pain
- shortness of breath
- dizziness
- fainting
- fever, sweating, confusion, fast or irregular heartbeat, severe muscle stiffness or twitching, agitation, hallucinations, loss of coordination, nausea, vomiting, or diarrhea
- coma (loss of consciousness)
- hives or blisters
- rash
- itching
- difficulty breathing or swallowing
- swelling of the face, throat, tongue, lips, eyes, hands, feet, ankles, or lower legs
- hoarseness
- unusual bleeding or bruising
- nose bleeding
- headache
- unsteadiness
- problems with thinking, concentration, or memory
- seizures
Citalopram may decrease appetite and cause weight loss in children. Your child’s doctor will watch his or her growth carefully. Talk to your child’s doctor if you have concerns about your child’s growth or weight while he or she is taking this medication. Talk to your child’s doctor about the risks of giving citalopram to your child.
Citalopram may cause other side effects. Call your doctor if you have any unusual problems while taking citalopram.
Escitalopram
Escitalopram belongs to the class of antidepressants known as selective serotonin reuptake inhibitors (SSRIs) 180. Escitalopram contains the same active medication as citalopram. These two drugs act in a very similar way by increasing the amount of serotonin, a natural substance in the brain that helps maintain mental balance. Escitalopram has been used to treat depression in adults and children and teenagers 12 years of ago or older. Escitalopram is also used to treat generalized anxiety disorder (GAD, excessive worry and tension that disrupts daily life and lasts for 6 months or longer) in adults, teenagers, and children 7 years of age and older.
Escitalopram was approved for use in the United States in 2002 and is available in tablets of 5, 10, and 20 mg and a solution (liquid) under the brand name Lexapro. It is usually taken once a day with or without food. The recommended dosage of escitalopram is 10 mg once daily, increasing to 20 mg daily if necessary. To help you remember to take escitalopram, take it at around the same time every day, in the morning or in the evening. Follow the directions on your prescription label carefully, and ask your doctor or pharmacist to explain any part you do not understand. Take escitalopram exactly as directed. Do not take more or less of it or take it more often than prescribed by your doctor.
Your doctor may start you on a low dose of escitalopram and increase your dose after 1 week in adults and after 2 or 3 weeks in teenagers and children 7 years of age and older.
It may take 1 to 4 weeks or longer before you feel the full benefit of escitalopram. Continue to take escitalopram even if you feel well. Do not stop taking escitalopram without talking to your doctor. Your doctor will probably decrease your dose gradually. If you suddenly stop taking escitalopram, you may experience withdrawal symptoms such as mood changes, irritability, agitation, nausea, dizziness, burning, numbness, or tingling in the hands or feet, anxiety, confusion, headache, sweating, shaking, frenzied or abnormally excited mood, tiredness, and difficulty falling asleep or staying asleep. Tell your doctor if you experience any of these symptoms while you are decreasing your dose of escitalopram or soon after you stop taking escitalopram.
A small number of children, teenagers, and young adults (up to 24 years of age) who took antidepressants (‘mood elevators’) such as escitalopram during clinical studies became suicidal (thinking about harming or killing oneself or planning or trying to do so). Children, teenagers, and young adults who take antidepressants to treat depression or other mental illnesses may be more likely to become suicidal than children, teenagers, and young adults who do not take antidepressants to treat these conditions. However, experts are not sure about how great this risk is and how much it should be considered in deciding whether a child or teenager should take an antidepressant. Children younger than 12 years of age should not normally take escitalopram, but in some cases, a doctor may decide that escitalopram is the best medication to treat a child’s condition.
You should know that your mental health may change in unexpected ways when you take escitalopram or other antidepressants even if you are an adult over 24 years of age. You may become suicidal, especially at the beginning of your treatment and any time that your dose is increased or decreased. You, your family, or your caregiver should call your doctor right away if you experience any of the following symptoms: new or worsening depression; thinking about harming or killing yourself, or planning or trying to do so; extreme worry; agitation; panic attacks; difficulty falling asleep or staying asleep; aggressive behavior; irritability; acting without thinking; severe restlessness; and frenzied abnormal excitement. Be sure that your family or caregiver knows which symptoms may be serious so they can call the doctor if you are unable to seek treatment on your own.
Your doctor will want to see you often while you are taking escitalopram, especially at the beginning of your treatment. Be sure to keep all appointments for office visits with your doctor.
The doctor or pharmacist will give you the manufacturer’s patient information sheet (Medication Guide) when you begin treatment with escitalopram. Read the information carefully and ask your doctor or pharmacist if you have any questions.
No matter your age, before you take an antidepressant, you, your parent, or your caregiver should talk to your doctor about the risks and benefits of treating your condition with an antidepressant or with other treatments. You should also talk about the risks and benefits of not treating your condition. You should know that having depression or another mental illness greatly increases the risk that you will become suicidal. Tell your doctor if you or anyone in your family has or has ever had bipolar disorder (mood that changes from depressed to abnormally excited) or mania (frenzied, abnormally excited mood) or has thought about or attempted suicide. Talk to your doctor about your condition, symptoms, and personal and family medical history. You and your doctor will decide what type of treatment is right for you.
Pregnancy
It is not known if escitalopram can make it harder for a woman to get pregnant 179. Studies in animals found that escitalopram might cause some reduced fertility (ability to get pregnant) 179.
Miscarriage is common and can occur in any pregnancy for many different reasons. There have been some studies that suggested taking antidepressant medications might slightly increase the chance of miscarriage. However, a study on citalopram and escitalopram did not find an increased chance of miscarriage in women taking these medications 179.
Every pregnancy starts out with a 3-5% chance of having a birth defect. This is called the background risk. There are reports of more than 15,000 pregnancies exposed to citalopram or escitalopram. Overall, the available data do not suggest that citalopram or escitalopram increases the chance of birth defects. While some reports have suggested an increased chance of heart defects or other birth defects, most studies have not found an increased chance of birth defects 179.
Some studies suggest that taking citalopram or escitalopram throughout the pregnancy might increase the chance of pregnancy-related problems such as preterm birth (birth before 37 weeks of pregnancy) and low birth weight (weighing less than 5 pounds, 8 ounces [2500 grams] at birth). However, research has also shown that when depression is left untreated during pregnancy, there could be an increased chance of pregnancy complications. This makes it hard to know if it is the medication, untreated depression, or other factors that are increasing the chance for these problems.
Some, but not all, studies have suggested that when pregnant women take SSRIs such as escitalopram or citalopram during the second half of the pregnancy, their babies might have an higher chance for a serious lung condition called persistent pulmonary hypertension. Persistent pulmonary hypertension happens in 1 or 2 out of 1,000 births. Among the studies looking at this, the overall chance for pulmonary hypertension when an SSRI was used in pregnancy was less than 1/100 (less than 1%).
Some medications taken during pregnancy are associated with withdrawal symptoms in a newborn after delivery. If you are taking escitalopram or citalopram at the time of delivery, your baby might have irritability, jitteriness, tremors (shivering), constant crying, different sleep patterns, problems with eating and controlling body temperature, and some problems with breathing. In most cases, these symptoms are mild and go away within a couple weeks with no treatment. Some babies may need to stay in the nursery or neonatal intensive care unit (NICU) until the symptoms go away. Most babies exposed to escitalopram or citalopram in late pregnancy do not have these symptoms. It is important that your doctor know you are taking these medications so that if symptoms occur your baby can get the care that is best for them.
Breastfeeding
Several studies have shown that small amounts of escitalopram are found in breast milk. There have been a few case reports of sleepiness and weight loss, but in most studies no harmful effects were seen in breastfed babies. Studies also showed no difference in the intellectual development of babies exposed to citalopram or escitalopram during breastfeeding. A nursing infant should be monitored for being very sleepy (hard to wake for feeds), poor feeding and poor weight gain, especially in younger, exclusively breastfed infants and when a combination of medications for mental health are used. Be sure to talk to your doctor about all your breastfeeding questions. An alternative to this medication should be prescribed or you should stop breastfeeding while using Escitalopram.
Escitalopram Dosage
The dose of Escitalopram will be different for different patients. Follow your doctor’s orders or the directions on the label. The following information includes only the average doses of Escitalopram. If your dose is different, do not change it unless your doctor tells you to do so.
The amount of medicine that you take depends on the strength of the medicine. Also, the number of doses you take each day, the time allowed between doses, and the length of time you take the medicine depend on the medical problem for which you are using the medicine.
For oral dosage forms (solution or tablets)
- For depression
- Adults and children 12 years of age and older: 10 milligrams (mg) once a day, taken either in the morning or evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 20 mg per day.
- Older adults: 10 mg once a day, taken either in the morning or evening.
- Children younger than 12 years of age: Use and dose must be determined by your doctor.
- For generalized anxiety disorder (GAD)
- Adults and children 7 years of age and older: At first, 10 milligrams (mg) once a day, taken either in the morning or evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 20 mg per day.
- Older adults: 10 mg once a day, taken either in the morning or evening.
- Children younger than 7 years of age: Use and dose must be determined by your doctor.
Missed Dose
If you miss a dose of Escitalopram, take it as soon as possible. However, if it is almost time for your next dose, skip the missed dose and go back to your regular dosing schedule. Do not double doses.
Escitalopram side effects
Escitalopram may cause side effects. Tell your doctor if any of these symptoms are severe or do not go away:
- nausea
- diarrhea
- constipation
- sexual problems in males; decreased sex drive, inability to get or keep an erection, or delayed or absent ejaculation
- sexual problems in females; decreased sex drive, or delayed orgasm or unable to have an orgasm
- drowsiness
- yawning
- shaking
- difficulty falling asleep or staying asleep
- increased sweating
- dizziness
- heartburn
- stomach pain
- excessive tiredness
- dry mouth
- decreased appetite
- weight loss
- flu-like symptoms
- runny nose
- sneezing
Some side effects can be serious. If you experience either of the following symptoms or those listed in the IMPORTANT WARNING or SPECIAL PRECAUTIONS sections, see your doctor immediately:
- unusual excitement
- seeing things or hearing voices that do not exist (hallucinating)
- rash
- hives or blisters
- itching
- fever
- joint pain
- difficulty breathing or swallowing
- swelling of the face, throat, tongue, lips, or eyes
- fever, sweating, confusion, fast or irregular heartbeat, severe muscle stiffness or twitching, agitation, hallucinations, loss of coordination, nausea, vomiting, or diarrhea
- abnormal bleeding or bruising
- nose bleeding
- headache
- unsteadiness
- problems with thinking, concentration, or memory
- seizures
- difficult or painful urination
Escitalopram may decrease appetite and cause weight loss and height in children. Your child’s doctor will watch his or her growth carefully. Talk to your child’s doctor if you have concerns about your child’s growth or weight while he or she is taking this medication. Talk to your child’s doctor about the risks of giving escitalopram to your child.
Escitalopram may cause other side effects. See your doctor if you have any unusual problems while taking Escitalopram.
Fluoxetine
Fluoxetine belongs to the class of antidepressants known as selective serotonin reuptake inhibitors (SSRIs) 181, 182. Fluoxetine works by increasing the amount of serotonin, a natural substance in the brain that helps maintain mental balance. Fluoxetine is a medication that has been used to treat depression, obsessive-compulsive disorder (OCD, bothersome thoughts that won’t go away and the need to perform certain actions over and over), Tourette’s syndrome, bulimia nervosa, panic disorder (a type of anxiety disorder characterized by recurring, unexpected panic attacks, which are sudden episodes of intense fear or discomfort, often accompanied by physical symptoms, and persistent worry about future attacks), and premenstrual dysphoric disorder [PMDD] symptoms (a more severe form of premenstrual syndrome [PMS] characterized by extreme mood swings, bloating, irritability, breast tenderness and depression, occurring in the week or two before menstruation). Fluoxetine has also been used to treat body dysmorphic disorder, borderline personality disorder, hot flushes of menopause, post traumatic stress disorder (PTSD), alcoholism, attention-deficit/hyperactivity disorder (ADHD), sleep disorders, headaches, mental illness, sexual problems, phobias and Raynaud’s phenomenon. Fluoxetine is also used along with olanzapine (Zyprexa) to treat depression that did not respond to other medications and episodes of depression in people with bipolar disorder type 1 (manic-depressive disorder; a disease that causes episodes of depression, episodes of mania, and other abnormal moods). Fluoxetine may be prescribed for other uses; ask your doctor for more information.
Fluoxetine was approved for use in the United States in 1987 and it became one of the most widely used antidepressant medications, more than 20 million prescriptions being written yearly 181.
Fluoxetine is available as tablets, delayed-release (releases the medication in the intestine) capsules of 10, 20 and 40 mg and in an oral solution (liquid) of 20 mg/5 mL, in multiple generic forms and under the brand names of Prozac and Sarafem. Fixed combinations of fluoxetine with olanzapine (Symbyax and generic forms) are also available. A long acting formulation of 90 mg of fluoxetine has been developed for once weekly dosing (Prozac weekly).
Fluoxetine may be taken with or without food. Fluoxetine capsules, tablets, and liquid are usually taken once a day in the morning or twice a day in the morning and at noon. The recommended dosage of standard formulations of fluoxetine in adults is 20 mg once daily, increasing to 40 mg daily if necessary and not exceeding 80 mg daily.
Fluoxetine delayed-released capsules are usually taken once a week. Take fluoxetine at around the same time(s) every day. Swallow the delayed-release capsules whole; do not cut, crush, or chew them. Follow the directions on your prescription label carefully, and ask your doctor or pharmacist to explain any part you do not understand. Take fluoxetine exactly as directed. Your doctor may start you on a low dose of fluoxetine and gradually increase your dose. Do not take more or less of it or take it more often than prescribed by your doctor.
It may take 4 to 5 weeks or longer before you feel the full benefit of fluoxetine. Continue to take fluoxetine even if you feel well. Do not stop taking fluoxetine without talking to your doctor. If you suddenly stop taking fluoxetine, you may experience withdrawal symptoms such as mood changes, irritability, agitation, dizziness, numbness or tingling in the hands or feet, anxiety, sweating, confusion, headache, tiredness, and difficulty falling asleep or staying asleep. Your doctor will probably decrease your dose gradually.
Common side effects of fluoxetine are drowsiness, dyspepsia, nausea, headache, increased sweating, increased appetite, weight gain and sexual dysfunction.
A small number of children, teenagers, and young adults (up to 24 years of age) who took antidepressants (‘mood elevators’) such as fluoxetine during clinical studies became suicidal (thinking about harming or killing oneself or planning or trying to do so). Children, teenagers, and young adults who take antidepressants to treat depression or other mental illnesses may be more likely to become suicidal than children, teenagers, and young adults who do not take antidepressants to treat these conditions. However, experts are not sure about how great this risk is and how much it should be considered in deciding whether a child or teenager should take an antidepressant.
You should know that your mental health may change in unexpected ways when you take fluoxetine or other antidepressants even if you are an adult over 24 years of age. You may become suicidal, especially at the beginning of your treatment and any time that your dose is increased or decreased. You, your family, or your caregiver should call your doctor right away if you experience any of the following symptoms: new or worsening depression; thinking about harming or killing yourself, or planning or trying to do so; extreme worry; agitation; panic attacks; difficulty falling asleep or staying asleep; aggressive behavior; irritability; acting without thinking; severe restlessness; and frenzied abnormal excitement. Be sure that your family or caregiver knows which symptoms may be serious so they can call the doctor if you are unable to seek treatment on your own.
Your doctor will want to see you often while you are taking fluoxetine, especially at the beginning of your treatment. Be sure to keep all appointments for office visits with your doctor.
The doctor or pharmacist will give you the manufacturer’s patient information sheet (Medication Guide) when you begin treatment with fluoxetine. Read the information carefully and ask your doctor or pharmacist if you have any questions.
No matter your age, before you take an antidepressant, you, your parent, or your caregiver should talk to your doctor about the risks and benefits of treating your condition with an antidepressant or with other treatments. You should also talk about the risks and benefits of not treating your condition. You should know that having depression or another mental illness greatly increases the risk that you will become suicidal. This risk is higher if you or anyone in your family has or has ever had bipolar disorder (mood that changes from depressed to abnormally excited) or mania (frenzied, abnormally excited mood) or has thought about or attempted suicide. Talk to your doctor about your condition, symptoms, and personal and family medical history. You and your doctor will decide what type of treatment is right for you.
Pregnancy
Fluoxetine has been studied in women having medical treatments because they were already having a hard time getting pregnant. In these studies, those who took fluoxetine got pregnant at the same rate as those who did not take fluoxetine 183.
Out of all babies born each year, about 3 out of 100 (3%) will have a birth defect. Doctors look at research studies to try to understand if an exposure, like fluoxetine, might increase the chance of birth defects in a pregnancy. Fluoxetine use is not expected to increase the chance of birth defects. There are reports of over 10,000 pregnancies exposed to fluoxetine in the first trimester (when many major birth defects can happen). No pattern of birth defects has been found and most studies have not found an increased chance of birth defects related to fluoxetine use.
Some studies have suggested an increased chance of heart defects or other birth defects. However, there is no proven increased chance of birth defects directly related to fluoxetine.
Some studies suggest a higher chance of preterm delivery (birth before week 37) or low birth weight (weighing less than 5 pounds, 8 ounces [2500 grams] at birth) with the use of fluoxetine in pregnancy. However, research has also shown that when conditions such as depression or anxiety are untreated or undertreated during pregnancy, there could be an increased chance of pregnancy complications. This makes it hard to know if it is the medication, the underlying condition, or other factors that might increase the chance for these problems.
Some, but not all, studies have suggested that when women who are pregnant take SSRIs during the second half of pregnancy, their babies might have an increased chance for a serious lung condition called persistent pulmonary hypertension (PPH). Persistent pulmonary hypertension happens in 1 or 2 out of 1,000 births. A recent report that combined results from several studies suggested the chance for persistent pulmonary hypertension might be increased if an SSRI was used during pregnancy. However, it was not clear if this was due to medication exposure or to other exposures that people who take SSRIs have in common, such as higher rates of smoking. Data from studies suggest the overall chance for persistent pulmonary hypertension when an SSRI is used in pregnancy is less than 1/100 (less than 1%).
The use of fluoxetine during pregnancy and/or in the third trimester can cause temporary symptoms in newborns soon after birth. These symptoms are sometimes referred to as withdrawal. Symptoms include being irritable and/or jittery, crying, tight muscles, trouble breathing, unusual sleep patterns, tremors (shivers), and/or trouble eating. In most cases symptoms are mild and go away in a few weeks with no treatment, or with only supportive care. Not all babies exposed to fluoxetine will have these symptoms. There might be a higher chance for withdrawal symptoms if other psychiatric medications are also taken with fluoxetine during pregnancy. It is important that your doctor know you are taking fluoxetine so that if symptoms occur your baby can get the care that is best for them.
Does taking fluoxetine in pregnancy affect future behavior or learning for the child?
A few studies have looked at the development of children from age 16 months to 7 years and did not find differences between children who were exposed to fluoxetine during pregnancy and those who were not. Most studies found no increase in attention deficit hyperactivity disorder (ADHD) in children exposed to SSRIs like fluoxetine during pregnancy. Most studies also find that SSRIs like fluoxetine do not appear to increase the chance of autism spectrum disorders (ASD) after adjusting for factors such as maternal illness.
Breastfeeding
Fluoxetine gets into breast milk and most reports find no side effects in breastfed babies 183. In a small number of cases, irritability, vomiting, diarrhea, and less sleep have been reported. One study noted slightly less weight gain in infants exposed to fluoxetine via breast milk; however, this would likely only be an issue if the infant’s weight gain was already a concern 183. One study showed that mental and physical development was normal for infants exposed to fluoxetine in breast milk in their first year of life 183. Studies in women breastfeeding have demonstrated harmful infant effects 184. If you suspect the baby has any symptoms (such as irritability, vomiting, diarrhea, trouble sleeping, or trouble gaining weight) contact the child’s doctor.
The product label for fluoxetine recommends women who are breastfeeding not use this medication. But the benefit of treating your condition might outweigh possible risks. Your doctor can talk with you about using fluoxetine and what treatment is best for you. Be sure to talk to your doctor about all your breastfeeding questions.
An alternative to Fluoxetine should be prescribed or you should stop breastfeeding while using Fluoxetine.
Fluoxetine Dosage
The dose of Fluoxetine will be different for different patients. Follow your doctor’s orders or the directions on the label. The following information includes only the average doses of Fluoxetine. If your dose is different, do not change it unless your doctor tells you to do so.
The amount of medicine that you take depends on the strength of the medicine. Also, the number of doses you take each day, the time allowed between doses, and the length of time you take the medicine depend on the medical problem for which you are using the medicine.
For oral dosage forms (capsules, delayed-release capsules, pulvules, or solution)
- For bulimia nervosa
- Adults: 60 milligrams (mg) once a day in the morning.
- Children: Use and dose must be determined by your doctor.
- For depression
- Adults: At first, 20 milligrams (mg) once a day in the morning. Your doctor may adjust your dose as needed. If you are taking more than 20 mg per day, you may take the capsule once a day in the morning or 2 times a day (eg, morning and noon). However, the dose is usually not more than 80 mg per day.
- Children 8 years of age and older: At first, 10 or 20 mg once a day in the morning. Your doctor may adjust your dose as needed.
- Children younger than 8 years of age: Use and dose must be determined by your doctor.
For depression associated with bipolar disorder (combination with olanzapine)
- Adults: At first, 20 milligrams (mg) of fluoxetine and 5 mg of olanzapine once a day, taken in the evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg of fluoxetine and 12 mg of olanzapine per day.
- Children 10 years of age and older: At first, 20 milligrams (mg) of fluoxetine and 2.5 mg of olanzapine once a day, taken in the evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg of fluoxetine and 12 mg of olanzapine per day.
- Children younger than 10 years of age: Use and dose must be determined by your doctor.
For treatment resistant depression (combination with olanzapine)
- Adults: At first, 20 milligrams (mg) of fluoxetine and 5 mg of olanzapine once a day, taken in the evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg of fluoxetine and 20 mg of olanzapine per day.
- Children: Use and dose must be determined by your doctor.
For obsessive-compulsive disorder (OCD)
- Adults: At first, 20 milligrams (mg) once a day in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 80 mg per day.
- Children 7 years of age and older: At first, 10 mg once a day in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 60 mg per day.
- Children younger than 7 years of age: Use and dose must be determined by your doctor.
For panic disorder
- Adults: At first, 10 milligrams (mg) once a day in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 60 mg per day.
- Children: Use and dose must be determined by your doctor.
For premenstrual dysphoric disorder (PMDD)
- Adults: At first, 20 milligrams (mg) once a day in the morning. Your doctor may have you take 20 mg every day of your menstrual cycle or for only 15 days of your cycle. Your doctor may adjust your dose as needed. However, the dose is usually not more than 80 mg per day.
- Children: Use and dose must be determined by your doctor.
Missed Dose
If you miss a dose of Fluoxetine, take it as soon as possible. However, if it is almost time for your next dose, skip the missed dose and go back to your regular dosing schedule. Do not double doses.
Fluoxetine side effects
Fluoxetine may cause side effects. Tell your doctor if any of these symptoms are severe or do not go away:
- nervousness
- anxiety
- difficulty falling asleep or staying asleep
- nausea
- diarrhea
- dry mouth
- heartburn
- yawning
- weakness
- uncontrollable shaking of a part of the body
- loss of appetite
- weight loss
- unusual dreams
- stuffy nose
- sexual problems in males; decreased sex drive, inability to get or keep an erection, or delayed or absent ejaculation
- sexual problems in females; decreased sex drive, or delayed orgasm or unable to have an orgasm
- excessive sweating
- headache, confusion, weakness, difficulty concentrating, or memory problems
Some side effects can be serious. If you experience any of the following symptoms or those listed in the IMPORTANT WARNING or SPECIAL PRECAUTIONS section, see your doctor immediately:
- rash
- hives or blisters
- itching
- fever
- joint pain
- swelling of the face, throat, tongue, lips, eyes, hands, feet, ankles, or lower legs
- difficulty breathing or swallowing
- agitation, fever, sweating, confusion, fast or irregular heartbeat, shivering, severe muscle stiffness or twitching, hallucinations, loss of coordination, nausea, vomiting, or diarrhea
- fast, slow, or irregular heartbeat
- shortness of breath
- dizziness or fainting
- seizures
- abnormal bleeding or bruising
Fluoxetine may decrease appetite and cause weight loss in children. Your child’s doctor will watch his or her growth carefully. Talk to your child’s doctor if you have concerns about your child’s growth or weight while he or she is taking this medication. Talk to your child’s doctor about the risks of giving fluoxetine to your child.
Fluoxetine may cause other side effects. See your doctor if you have any unusual problems while taking fluoxetine.
Paroxetine
Paroxetine is a selective serotonin reuptake inhibitor (SSRI) that has been used to treat depression, generalized anxiety disorder (GAD, excessive worrying that is difficult to control), social anxiety disorder (extreme fear of interacting with others or performing in front of others that interferes with normal life), obsessive compulsive disorder (OCD, bothersome thoughts that won’t go away and the need to perform certain actions over and over), premenstrual dysphoric disorder (PMDD, physical and psychological symptoms that occur before the onset of the menstrual period each month), post-traumatic stress disorder (PTSD, disturbing psychological symptoms that develop after a frightening experience), and panic disorder (sudden, unexpected attacks of extreme fear and worry about these attacks). Paroxetine is also used to treat chronic headaches, diabetic neuropathy (numbness in the hands and feet caused by diabetes), premature ejaculation and hot flashes (sudden feelings of warmth, especially in the face, neck, and chest) in women who are experiencing menopause (stage of life when menstrual periods become less frequent and stop and women may experience other symptoms and body changes). There is not enough information available at this time to know how paroxetine works to treat hot flashes in menopause. Paroxetine is also sometimes used with other medications to treat bipolar disorder (mood that changes from depressed to abnormally excited). Paroxetine may be prescribed for other uses; ask your doctor or pharmacist for more information.
Paroxetine was approved for use in the United States in 1992 and it remains in wide use, with more than 15 million prescriptions being filled yearly 185.
Paroxetine is available as tablets of 10, 20, 30 and 40 mg and an oral suspension (liquid) in generic forms and under the brand names of Paxil and Pexeva. The recommended dosage for depression in adults is 20 mg once daily, increasing the dosage by 10 mg increments weekly to a maximum of 50 mg. Controlled release or extended-release (long-acting) tablets are also available that have slightly different dosing recommendations.
The tablets, suspension, and controlled-release tablets are usually taken once daily in the morning or evening, with or without food. The capsules are usually taken once a day at bedtime with or without food. You may want to take paroxetine with food to prevent stomach upset. Take paroxetine at around the same time every day. Follow the directions on your prescription label carefully, and ask your doctor or pharmacist to explain any part you do not understand. Take paroxetine exactly as directed. Do not take more or less of it or take it more often than prescribed by your doctor.
Shake the liquid well before each use to mix the medication evenly.
Swallow the extended-release and regular tablets whole; do not chew or crush them.
If you are taking paroxetine tablets, suspension, or controlled-release tablets, your doctor may start you on a low dose of paroxetine and gradually increase your dose, not more than once a week.
Paroxetine capsules contain a lower dose of paroxetine than is needed to treat depression and other forms of mental illness. Do not take paroxetine capsules to treat a mental illness. If you think you have depression or another mental illness, talk to your doctor about treatment.
Paroxetine may help control your symptoms but will not cure your condition. It may take several weeks or longer before you feel the full benefit of paroxetine. Continue to take paroxetine even if you feel well. Do not stop taking paroxetine without talking to your doctor. Your doctor may decrease your dose gradually. If you suddenly stop taking paroxetine tablets, suspension, or controlled-release tablets, you may experience withdrawal symptoms such as depression; mood changes; frenzied or abnormally excited mood; irritability; anxiety; confusion; dizziness; headache; tiredness; numbness or tingling in the arms, legs, hands, or feet; unusual dreams; difficulty falling asleep or staying asleep; nausea; or sweating. Tell your doctor if you experience any of these symptoms when your dose of paroxetine is decreased.
Common side effects of Paroxetine are drowsiness, dyspepsia, nausea, headache, increased sweating, increased appetite, weight gain and sexual dysfunction. Rare but potentially severe adverse events include suicidal ideation and behavior, activation of mania, serotonin syndrome, discontinuation syndrome, increased risk of bleeding, seizures, acute glaucoma, hypersensitivity reactions and embryo-fetal toxicity.
A small number of children, teenagers, and young adults (up to 24 years of age) who took antidepressants (‘mood elevators’) such as paroxetine during clinical studies became suicidal (thinking about harming or killing oneself or planning or trying to do so). Children, teenagers, and young adults who take antidepressants to treat depression or other mental illnesses may be more likely to become suicidal than children, teenagers, and young adults who do not take antidepressants to treat these conditions. However, there are also risks when depression is not treated in children and teenagers. Talk to your child’s doctor about these risks and whether your child should take an antidepressant, Children younger than 18 years of age should not normally take paroxetine, but in some cases, a doctor may decide that paroxetine is the best medication to treat a child’s condition.
You should know that your mental health may change in unexpected ways when you take paroxetine or other antidepressants to treat depression or other mental illness even if you are an adult over 24 years of age. You may also experience changes in your mental health if you are a woman taking a low dose of paroxetine to treat hot flashes and you have never had depression or another mental illness. You may become suicidal, especially at the beginning of your treatment and any time that your dose is increased or decreased. You, your family, or your caregiver should call your doctor right away if you experience any of the following symptoms: new or worsening depression; thinking about harming or killing yourself, or planning or trying to do so; extreme worry; agitation; panic attacks; difficulty falling asleep or staying asleep; aggressive behavior; irritability; acting without thinking; severe restlessness; and frenzied abnormal excitement. Be sure that your family or caregiver knows which symptoms may be serious so they can call the doctor if you are unable to seek treatment on your own.
Your doctor will want to see you often while you are taking paroxetine, especially at the beginning of your treatment. Be sure to keep all appointments for office visits with your doctor.
The doctor or pharmacist will give you the manufacturer’s patient information sheet (Medication Guide) when you begin treatment with paroxetine. Read the information carefully and ask your doctor or pharmacist if you have any questions.
No matter what your age, before you take an antidepressant, you, your parent, or your caregiver should talk to your doctor about the risks and benefits of treating your condition with an antidepressant or with other treatments. You should also talk about the risks and benefits of not treating your condition. You should know that having depression or another mental illness greatly increases the risk that you will become suicidal. This risk is higher if you or anyone in your family has or has ever had bipolar disorder (mood that changes from depressed to abnormally excited) or mania (frenzied, abnormally excited mood) or has thought about or attempted suicide. Talk to your doctor about your condition, symptoms, and personal and family medical history. You and your doctor will decide what type of treatment is right for you.
Pregnancy
Paroxetine is contraindicated in pregnancy and is classified as category D/X due to its teratogenic effects in causing cardiovascular defects, specifically heart malformations if prescribed in the first trimester 164.
In some women, paroxetine may raise the levels of a hormone called prolactin 186. High levels of prolactin can stop ovulation (part of the menstrual cycle when an ovary releases an egg). This can make it harder to get pregnant. Paroxetine has also been associated with changes in sexual function such as less desire to have sex 186.
Miscarriage is common and can occur in any pregnancy for many different reasons. Some studies suggest that taking paroxetine may slightly increase the chance of miscarriage. However, research also shows that depression itself may increase the chance of miscarriage. This makes it hard to know if an increased chance of miscarriage is due to medication, the underlying condition, or other factors.
Every pregnancy starts out with a 3-5% chance of having a birth defect. This is called the background risk. Some studies suggest that exposure to paroxetine in the first trimester might be associated with a small increased chance for heart defects. Other studies did not find a possible increased risk. The background rate of heart defects for any pregnancy is about 1 in 100 (1%). If there is an increased chance of birth defects with paroxetine use in the first trimester, it is expected to be small. Paroxetine exposure in pregnancy has not been shown to increase the chance of other birth defects.
Some studies suggest a higher chance for preterm delivery (birth before week 37) with the use of an SSRI such as paroxetine, while others do not. Conditions such as depression may increase the chance of pregnancy-related problems, such as preterm delivery. This makes it hard to know if it is the medication, the condition being treating, or other factors that are increasing the chance of pregnancy-related problems.
Some, but not all, studies have suggested that when women who are pregnant take SSRIs during the second half of the pregnancy, their babies might have a higher chance for a serious lung condition called persistent pulmonary hypertension 186. Persistent pulmonary hypertension happens in 1 or 2 out of 1,000 births. Among the studies looking at this, the overall chance for pulmonary hypertension when an SSRI was used in pregnancy was less than 1/100 (less than 1%).
The use of paroxetine during pregnancy can cause temporary symptoms in newborns soon after birth. These symptoms are sometimes referred to as withdrawal. Symptoms may include jitteriness, increased muscle tone, irritability, changes in sleep patterns, tremors, trouble eating, and trouble breathing. These symptoms are usually mild and go away on their own. Some babies may need to stay in a special care nursery for several days. Not all babies exposed to paroxetine will have these symptoms. It is important that your doctor know you are taking paroxetine so that if symptoms occur your baby can get the care that’s best for them.
It is not known if paroxetine can increase the chance for behavior or learning issues. One study looking at prescriptions for SSRIs including paroxetine suggested an increased chance of autism spectrum disorder. Studies based on filled prescriptions/prescription records cannot tell if a person took the medication, so it is hard to know if the outcomes are related to the medication or other factors. Also, this study did not look at factors such as other exposures, paternal mental illness, or other family history of autism. Another prescription study did not report an increased chance for autism spectrum disorder.
One study looking at pregnancies exposed at least during the third trimester to SSRIs including paroxetine reported no differences in developmental outcomes between the exposed 46 infants or the unexposed 23 infants at 2 and 8 months of age. A follow-up study on some of these children found no difference in behaviors such as emotional reactivity, withdrawal, irritability, depression, or anxiety in the exposed group (22 children) when compared to the group that was not exposed to paroxetine (14 children).
Breastfeeding
Paroxetine passes into breastmilk in small amounts. In some cases, mild side effects have been reported. If you suspect the baby has any symptoms (trouble sleeping, restlessness, or increased crying), contact the child’s doctor. Be sure to talk to your doctor about all your breastfeeding questions. An alternative to Paroxetine should be prescribed or you should stop breastfeeding while using Paroxetine.
Paroxetine dosage
The dose of Paroxetine will be different for different patients. Follow your doctor’s orders or the directions on the label. The following information includes only the average doses of Paroxetine. If your dose is different, do not change it unless your doctor tells you to do so.
The amount of medicine that you take depends on the strength of the medicine. Also, the number of doses you take each day, the time allowed between doses, and the length of time you take the medicine depend on the medical problem for which you are using the medicine.
Oral dosage form (capsules)
- For moderate to severe hot flashes caused by menopause
- Adults: 7.5 milligrams (mg) once a day, at bedtime.
- Children: Use is not recommended.
Oral dosage form (suspension)
- For depression
- Adults: At first, 20 milligrams (mg) (10 milliliters [mL]) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg (25 mL) per day.
- Older adults: At first, 10 mg (5 mL) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 40 mg (20 mL) per day.
- Children: Use and dose must be determined by your doctor.
- For generalized anxiety disorder (GAD)
- Adults: At first, 20 milligrams (mg) (10 milliliters [mL]) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg (25 mL) per day.
- Older adults: At first, 10 mg (5 mL) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 40 mg (20 mL) per day.
- Children: Use and dose must be determined by your doctor.
- For obsessive-compulsive disorder (OCD)
- Adults: At first, 20 milligrams (mg) (10 milliliters [mL]) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 60 mg (30 mL) per day.
- Older adults: At first, 10 mg (5 mL) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 40 mg (20 mL) per day.
- Children: Use and dose must be determined by your doctor.
- For panic disorder
- Adults: At first, 10 milligrams (mg) (5 milliliters [mL]) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 60 mg (30 mL) per day.
- Older adults: At first, 10 mg (5 mL) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 40 mg (20 mL) per day.
- Children: Use and dose must be determined by your doctor.
- For post traumatic stress disorder (PTSD)
- Adults: At first, 20 milligrams (mg) (10 milliliters [mL]) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg (25 mL) per day.
- Older adults: At first, 10 mg (5 mL) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose usually is not more than 40 mg (20 mL) per day.
- Children: Use and dose must be determined by your doctor.
- For social anxiety disorder
- Adults: At first, 20 milligrams (mg) (10 milliliters [mL]) once a day, usually taken in the morning.
- Older adults: At first, 10 mg (5 mL) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose usually is not more than 20 mg (10 mL) per day.
- Children: Use and dose must be determined by your doctor.
Oral dosage form (tablets)
- For depression
- Adults: At first, 20 milligrams (mg) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg per day.
- Older adults: At first, 10 mg once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 40 mg per day.
- Children: Use and dose must be determined by your doctor.
- For generalized anxiety disorder (GAD)
- Adults: At first, 20 milligrams (mg) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg per day.
- Older adults: At first, 10 mg once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose usually is not more than 40 mg per day.
- Children: Use and dose must be determined by your doctor.
- For obsessive-compulsive disorder (OCD)
- Adults: At first, 20 milligrams (mg) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose usually is not more than 60 mg per day.
- Older adults: At first, 10 mg once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 40 mg per day.
- Children: Use and dose must be determined by your doctor.
- For panic disorder
- Adults: At first, 10 milligrams (mg) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 60 mg per day.
- Older adults: At first, 10 mg once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 40 mg per day.
- Children: Use and dose must be determined by your doctor.
- For post traumatic stress disorder (PTSD)
- Adults: At first, 20 milligrams (mg) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose usually is not more than 50 mg per day.
- Older adults: At first, 10 mg once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 40 mg per day.
- Children: Use and dose must be determined by your doctor.
- For social anxiety disorder
- Adults: At first, 20 milligrams (mg) once a day, usually taken in the morning.
- Older adults: At first, 10 mg once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 20 mg per day.
- Children: Use and dose must be determined by your doctor.
Oral dosage form (extended-release tablets)
- For depression
- Adults: At first, 25 milligrams (mg) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose usually is not more than 62.5 mg per day.
- Older adults: At first, 12.5 mg once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg per day.
- Children: Use and dose must be determined by your doctor.
- For panic disorder
- Adults: At first, 12.5 milligrams (mg) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose usually is not more than 75 mg per day.
- Older adults: At first, 12.5 mg once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 50 mg per day.
- Children: Use and dose must be determined by your doctor.
- For premenstrual dysphoric disorder (PMDD)
- Adults: At first, 12.5 milligrams (mg) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 25 mg per day.
- Older adults and children: Use and dose must be determined by your doctor.
- For social anxiety disorder
- Adults: At first, 12.5 milligrams (mg) once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 37.5 mg per day.
- Older adults: At first, 12.5 mg once a day, usually taken in the morning. Your doctor may adjust your dose as needed. However, the dose is usually not more than 37.5 mg per day.
- Children: Use and dose must be determined by your doctor.
Missed Dose
If you miss a dose of Paroxetine, take it as soon as possible. However, if it is almost time for your next dose, skip the missed dose and go back to your regular dosing schedule. Do not double doses.
Paroxetine side effects
Paroxetine may cause side effects. Tell your doctor if any of these symptoms are severe or do not go away:
- headache
- dizziness
- weakness
- difficulty concentrating
- nervousness
- forgetfulness
- confusion
- sleepiness
- nausea
- vomiting
- diarrhea
- constipation
- gas
- stomach pain
- heartburn
- changes in ability to taste food
- decreased appetite
- weight loss or gain
- sexual problems in males; decreased sex drive, inability to get or keep an erection, or delayed or absent ejaculation
- sexual problems in females; decreased sex drive, or delayed orgasm or unable to have an orgasm
- dry mouth
- sweating
- yawning
- pain in the back, muscles, bones, or anywhere in the body
- tenderness or swelling of joints
Some side effects can be serious. If you experience any of the following symptoms or those listed in the IMPORTANT WARNING or SPECIAL PRECAUTIONS sections, call your doctor immediately or get emergency medical treatment:
- seeing things or hearing voices that do not exist (hallucinating)
- fainting
- rapid, pounding, or irregular heartbeat
- chest pain
- difficulty breathing
- seizures
- agitation, fever, sweating, confusion, fast or irregular heartbeat, and severe muscle stiffness or twitching, hallucinations, loss of coordination, nausea, vomiting, or diarrhea
- abnormal bleeding or bruising
- tiny red spots directly under the skin
- peeling or blistering of skin
- sore throat, fever, chills, cough, and other signs of infection
- uncontrollable shaking of a part of the body
- unsteady walking that may cause falling
- numbness or tingling in your hands, feet, arms, or legs
- painful erection that lasts for hours
- sudden nausea, vomiting, weakness, cramping, bloating, swelling, tightness in hands and feet, dizziness, headache and/or confusion
- hives
- rash
- itching
- swelling of the face, throat, tongue, lips, eyes, hands, feet, ankles, or lower legs
- hoarseness
- black and tarry stools
- red blood in stools
- bloody vomit
- vomit that looks like coffee grounds
- bone pain or fracture
- tenderness, swelling, or bruising of one part of your body
Paroxetine may decrease appetite and cause weight loss in children. Your child’s doctor will watch his or her growth carefully. Talk to your child’s doctor if you have concerns about your child’s growth or weight while he or she is taking this medication. Talk to your child’s doctor about the risks of giving paroxetine to your child.
Paroxetine may cause other side effects. See your doctor if you have any unusual problems while taking Paroxetine.
Sertraline
Sertraline is a selective serotonin reuptake inhibitor (SSRI) medication that has been used to treat depression, obsessive-compulsive disorder (OCD, bothersome thoughts that won’t go away and the need to perform certain actions over and over), panic attacks (sudden, unexpected attacks of extreme fear and worry about these attacks), and major anxiety disorders including social anxiety disorder, post-trauma stress disorder (PTSD, disturbing psychological symptoms that develop after a frightening experience) and generalized anxiety disorder (GAD), panic disorder, premenstrual dysphoric disorder [PMDD] (a severe form of premenstrual syndrome [PMS] with mood swings, irritability, bloating, and breast tenderness), and social anxiety disorder (extreme fear of interacting with others or performing in front of others that interferes with normal life). Sertraline is also used for headache, diabetic neuropathy and premature ejaculation. Sertraline works by blocking the reuptake of serotonin in your brain neurons, thus increasing serotonin levels in your brain which is associated with its psychiatric effects.
Sertraline was approved for use in the United States in 1991, and it remains in wide use, with almost 40 million prescriptions being filled yearly 187.
Sertraline is available as tablets of 25, 50 and 100 mg and as an oral suspension (liquid) in multiple generic forms and under the brand name of Zoloft. Sertraline is usually taken once daily in the morning or evening. The recommended dosage for depression in adults is 50 or 100 mg once daily, increasing the dosage by 25 or 50 mg increments to a maximum of 200 mg. Take sertraline at around the same time every day. Follow the directions on your prescription label carefully, and ask your doctor or pharmacist to explain any part you do not understand. Take sertraline exactly as directed. Do not take more or less of it or take it more often than prescribed by your doctor.
To treat premenstrual dysphoric disorder (PMDD), sertraline is taken once a day, either every day of the month or on certain days of the month.
Sertraline concentrate must be diluted before use. Immediately before taking it, use the provided dropper to remove the amount of concentrate your doctor has told you to take. Mix the concentrate with 4 ounces (1/2 cup [120 milliliters]) of water, ginger ale, lemon or lime soda, lemonade, or orange juice. After mixing, the diluted solution may be hazy; this is normal. Do not mix the concentrate with any liquids other than the ones listed. Drink the diluted solution immediately.
Your doctor may start you on a low dose of sertraline and gradually increase your dose, not more than once a week.
It may take a few weeks or longer before you feel the full benefit of sertraline. Continue to take sertraline even if you feel well. Do not stop taking sertraline without talking to your doctor. If you suddenly stop taking sertraline, you may experience withdrawal symptoms such as nausea, sweating, depression, mood changes, frenzied or abnormally excited mood, irritability, anxiety, confusion, dizziness, headache, tiredness, seizures, ringing in the ears, numbness or tingling in the arms, legs, hands, or feet, difficulty falling asleep or staying asleep.
Common side effects of Sertraline are drowsiness, dyspepsia, nausea, headache, increased sweating, increased appetite, weight gain and sexual dysfunction. Rare but potentially severe adverse events include suicidal ideation and behavior, activation of mania, serotonin syndrome, discontinuation syndrome, increased risk of bleeding, seizures, acute glaucoma, hypersensitivity reactions and embryo-fetal toxicity.
A small number of children, teenagers, and young adults (up to 24 years of age) who took antidepressants (‘mood elevators’) such as sertraline during clinical studies became suicidal (thinking about harming or killing oneself or planning or trying to do so). Children, teenagers, and young adults who take antidepressants to treat depression or other mental illnesses may be more likely to become suicidal than children, teenagers, and young adults who do not take antidepressants to treat these conditions. However, experts are not sure about how great this risk is and how much it should be considered in deciding whether a child or teenager should take an antidepressant.
You should know that your mental health may change in unexpected ways when you take sertraline or other antidepressants even if you are an adult over 24 years of age. You may become suicidal, especially at the beginning of your treatment and any time that your dose is increased or decreased. You, your family, or your caregiver should call your doctor right away if you experience any of the following symptoms: new or worsening depression; thinking about harming or killing yourself, or planning or trying to do so; extreme worry; agitation; panic attacks; new or worsening anxiety; difficulty falling asleep or staying asleep; aggressive behavior; irritability; acting without thinking; severe restlessness; and frenzied abnormal excitement. Be sure that your family or caregiver knows which symptoms may be serious so they can call the doctor if you are unable to seek treatment on your own.
Your doctor will want to see you often while you are taking sertraline, especially at the beginning of your treatment. Be sure to keep all appointments for office visits with your doctor.
The doctor or pharmacist will give you the manufacturer’s patient information sheet (Medication Guide) when you begin treatment with sertraline. Read the information carefully and ask your doctor or pharmacist if you have any questions.
No matter what your age, before you take an antidepressant, you, your parent, or your caregiver should talk to your doctor about the risks and benefits of treating your condition with an antidepressant or with other treatments. You should also talk about the risks and benefits of not treating your condition. You should know that having depression or another mental illness greatly increases the risk that you will become suicidal. This risk is higher if you or anyone in your family has or has ever had bipolar disorder (mood that changes from depressed to abnormally excited) or mania (frenzied, abnormally excited mood) or has thought about or attempted suicide. Talk to your doctor about your condition, symptoms, and personal and family medical history. You and your doctor will decide what type of treatment is right for you.
Pregnancy
It is not known if sertraline can make it harder to get pregnant. One study found that women who take SSRIs have a slightly lower chance of getting pregnant. However, some conditions, including depression, can make it harder to get pregnant. This makes it hard to know if the medication, the condition being treated, or other factors might affect fertility.
Miscarriage is common and can occur in any pregnancy for many different reasons. Use of sertraline and the chance of miscarriage has not been well studied. One study found no differences in the chance of miscarriage in women who filled prescriptions for sertraline during the first 35 days of pregnancy and those who stopped filling prescriptions before pregnancy. Also, some conditions, including depression, may increase the chance of miscarriage. This makes it hard to know if the medication, the condition being treated, or other factors might affect the chance of miscarriage.
Every pregnancy starts out with a 3-5% chance of having a birth defect. This is called the background risk. There are reports of more than 25,000 pregnancies exposed to sertraline. Some studies have suggested an increased chance for heart defects or other birth defects. However, most studies have not found an increased chance of birth defects when sertraline is used in pregnancy. Overall, the available data does not suggest that sertraline increases the chance of birth defects above the background risk.
Some studies suggest a higher chance for pregnancy-related problems, such as preterm delivery (birth before week 37) or low birth weight (weighing less than 5 pounds, 8 ounces [2500 grams] at birth) with the use of sertraline in pregnancy. However, research has also shown that when conditions such as depression or anxiety are untreated or undertreated during pregnancy, there could be an increased chance for pregnancy complications. This makes it hard to know if it is the medication, the underlying condition, or other factors that might increase the chance for these problems.
Some, but not all, studies have suggested that when women who are pregnant take SSRIs during the second half of pregnancy, their babies might have an increased chance for a serious lung condition called persistent pulmonary hypertension (PPH). Persistent pulmonary hypertension happens in 1 or 2 out of 1,000 births. A recent report that combined results from several studies suggested the chance for persistent pulmonary hypertension might be increased if an SSRI was used during pregnancy. However, it was not clear if this was due to medication exposure or to other exposures that people who take SSRIs have in common, such as higher rates of smoking. Data from studies suggest the overall chance for persistent pulmonary hypertension when an SSRI is used in pregnancy is less than 1/100 (less than 1%).
The use of sertraline during pregnancy can cause temporary symptoms in newborns soon after birth. These symptoms are sometimes referred to as withdrawal. Symptoms can include irritability, jitteriness, tremors (shivering), constant crying, changes in sleep patterns, lower muscle tone (hypotonia), skin discoloration (cyanosis), problems with eating, trouble controlling body temperature, and problems with breathing (apnea). In most cases, these symptoms are mild and go away within a couple weeks with no treatment required. Some babies may need to stay in the nursery or neonatal intensive care unit (NICU) until the symptoms go away. Not all babies exposed to sertraline will have these symptoms. It is important that your doctor know you are taking sertraline so that if symptoms do occur, your baby can get the care that is best for them.
One study on a small number of children who were exposed to SSRIs during pregnancy reported a lower score on motor skill tests than other children. Another small study looked at behaviors in children ages 4-5 years old. This study found no difference in behavior between children who were exposed to sertraline or other SSRIs during pregnancy and those children who were not.
Breastfeeding
Sertraline gets into breastmilk in small amounts. Most reports show no problems for babies who are exposed to sertraline through breast milk. Babies who were also exposed to sertraline in the third trimester of pregnancy may have a lower chance of withdrawal after birth if they are breastfed. Be sure to talk to your doctor about all your breastfeeding questions.
Sertraline dosage
The dose of Sertraline will be different for different patients. Follow your doctor’s orders or the directions on the label. The following information includes only the average doses of Sertraline. If your dose is different, do not change it unless your doctor tells you to do so.
The amount of medicine that you take depends on the strength of the medicine. Also, the number of doses you take each day, the time allowed between doses, and the length of time you take the medicine depend on the medical problem for which you are using the medicine.
For oral dosage forms (solution or tablets)
- For depression
- Adults: At first, 50 milligrams (mg) once a day, taken either in the morning or evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 200 mg per day.
- Children: Use and dose must be determined by your doctor.
- For obsessive-compulsive disorder (OCD)
- Adults and teenagers: At first, 50 milligrams (mg) once a day, taken either in the morning or evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 200 mg per day.
- Children 6 to 12 years of age: At first, 25 mg once a day, taken either in the morning or evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 200 mg per day.
- Children younger than 6 years of age: Use and dose must be determined by your doctor.
- For panic disorder, posttraumatic stress disorder (PTSD) or social anxiety disorder
- Adults: At first, 25 milligrams (mg) once a day, taken either in the morning or evening. Your doctor may adjust your dose as needed. However, the dose is usually not more than 200 mg per day.
- Children: Use and dose must be determined by your doctor.
- For premenstrual dysphoric disorder (PMDD)
- Adults: At first, 50 milligrams (mg) once a day throughout your menstrual cycle or just during the premenstrual time. Your doctor may adjust your dose as needed. However, the dose is usually not more than 150 mg per day throughout your menstrual cycle or 100 mg per day if you are only taking it during your premenstrual time.
- Children: Use and dose must be determined by your doctor.
Missed Dose
If you miss a dose of Sertraline, take it as soon as possible. However, if it is almost time for your next dose, skip the missed dose and go back to your regular dosing schedule. Do not double doses.
Sertraline side effects
Sertraline may cause side effects. See your doctor if any of these symptoms are severe or do not go away:
- nausea
- diarrhea
- constipation
- vomiting
- difficulty falling asleep or staying asleep
- dry mouth
- heartburn
- loss of appetite
- weight changes
- dizziness
- excessive tiredness
- headache
- nervousness
- uncontrollable shaking of a part of the body
- sexual problems in males; decreased sex drive, inability to get or keep an erection, or delayed or absent ejaculation
- sexual problems in females; decreased sex drive, or delayed orgasm or unable to have an orgasm
- excessive sweating
Some side effects can be serious. If you experience any of the following symptoms or those listed in the IMPORTANT WARNING or SPECIAL PRECAUTIONS section, see your doctor immediately:
- seizures
- abnormal bleeding or bruising
- agitation, hallucinations, fever, sweating, confusion, fast heartbeat, shivering, severe muscle stiffness or twitching, loss of coordination, nausea, vomiting, or diarrhea
- headache, weakness, unsteadiness, confusion, or memory problems
- rash
- hives
- swelling
- difficulty breathing
Sertraline may decrease appetite and cause weight loss in children. Your child’s doctor will watch his or her growth carefully. Talk to your child’s doctor if you have concerns about your child’s growth or weight while he or she is taking this medication. Talk to your child’s doctor about the risks of giving sertraline to your child.
Sertraline may cause other side effects. Call your doctor if you have any unusual problems while taking Sertraline.
Vilazodone
Vilazodone is a serotonin reuptake inhibitor (SSRI) and partial serotonin receptor agonist also called serotonin modulators 188, 189. Vilazodone works by blocking the reuptake of serotonin in your brain neurons, thus increasing serotonin levels in your brain which is associated with its psychiatric effects. Vilazodone is also a partial serotonin (5-HT1A) receptor agonist, which may add to its antidepressant effects. Vilazodone is used to treat depression.
Vilazodone was approved for use in the United States in 2011 for use in treatment of major depressive disorder 188. However, there is limited clinical experience with its use.
Vilazodone is available as tablets of 10, 20 and 40 mg under the brand name Viibryd. Take vilazodone at around the same time every day. The recommended initial dose of vilazodone in adults is 10 mg daily, which can then be increased to the typical maintenance dose of 40 mg once daily. Vilazodone is usually taken with food once a day. Follow the directions on your prescription label carefully, and ask your doctor or pharmacist to explain any part you do not understand. Take vilazodone exactly as directed. Do not take more or less of it or take it more often than prescribed by your doctor.
Your doctor will probably start you on a low dose of vilazodone and gradually increase your dose, not more than once every 7 days.
Vilazodone controls depression but does not cure it. It may take several weeks before you feel the full benefit of vilazodone. Continue to take vilazodone even if you feel well. Do not stop taking vilazodone without talking to your doctor. Your doctor will probably decrease your dose gradually. If you suddenly stop taking vilazodone, you may experience withdrawal symptoms such as dizziness; nausea; headache; confusion; irritability; agitation; difficulty falling asleep or staying asleep; anxiety; extreme tiredness; seizures; pain, burning, or tingling in the hands or feet; frenzied or abnormally excited mood; or sweating. Tell your doctor if you experience any of these symptoms while you are decreasing your dose of vilazodone or soon after you stop taking vilazodone.
Common, non-serious side effects of Vilazodone include diarrhea, nausea, fatigue, drowsiness, headache, insomnia, weight gain and sexual dysfunction. Vilazodone overdose is associated with acute serotonin syndrome. Rare, but potentially severe adverse effects include suicidal thinking and behavior, activation of symptoms of mania, serotonin syndrome, sexual dysfunction, hyponatremia and hypersensitivity reactions.
A small number of children, teenagers, and young adults (up to 24 years of age) who took antidepressants (‘mood elevators’) such as vilazodone during clinical studies became suicidal (thinking about harming or killing oneself or planning or trying to do so). Children, teenagers, and young adults who take antidepressants to treat depression or other mental illnesses may be more likely to become suicidal than children, teenagers, and young adults who do not take antidepressants to treat these conditions. However, experts are not sure about how great this risk is and how much it should be considered in deciding whether a child or teenager should take an antidepressant. Children younger than 18 years of age should not normally take vilazodone, but in some cases, a doctor may decide that vilazodone is the best medication to treat a child’s condition.
You should know that your mental health may change in unexpected ways when you take vilazodone or other antidepressants, even if you are an adult over 24 years of age. You may become suicidal, especially at the beginning of your treatment and any time that your dose is increased or decreased. You, your family, or your caregiver should call your doctor right away if you experience any of the following symptoms: new or worsening depression; thinking about harming or killing yourself, or planning or trying to do so; extreme worry; agitation; panic attacks; difficulty falling asleep or staying asleep; aggressive behavior; irritability; acting without thinking; severe restlessness; frenzied abnormal excitement; or any other changes in your usual thoughts, mood, or behavior. Be sure that your family or caregiver knows which symptoms may be serious so they can call the doctor if you are unable to seek treatment on your own.
Your doctor will want to see you often while you are taking vilazodone, especially at the beginning of your treatment. Be sure to keep all appointments for office visits with your doctor.
Your doctor or pharmacist will give you the manufacturer’s patient information sheet (Medication Guide) when you begin treatment with vilazodone and each time you refill your prescription. Read the information carefully and ask your doctor or pharmacist if you have any questions.
No matter your age, before you take an antidepressant, you, your parent, or your caregiver should talk to your doctor about the risks and benefits of treating your condition with an antidepressant or with other treatments. You should also talk about the risks and benefits of not treating your condition. You should know that having depression or another mental illness greatly increases the risk that you will become suicidal. This risk is higher if you or anyone in your family has or has ever had bipolar disorder (mood that changes from depressed to abnormally excited) or mania (frenzied, abnormally excited mood) or has thought about or attempted suicide. Talk to your doctor about your condition, symptoms, and personal and family medical history. You and your doctor will decide what type of treatment is right for you.
Pregnancy
It is not known if vilazodone can make it harder to get pregnant 190.
Miscarriage is common and can occur in any pregnancy for many different reasons. Studies have not been done to see if vilazodone increases the chance for miscarriage. However, depression itself might increase the chance for miscarriage.
Every pregnancy starts out with a 3-5% chance of having a birth defect. This is called the background risk. Animal studies did not find an increased chance of birth defects. We have been unable to locate studies on the use of vilazodone in human pregnancies.
Studies have not been done to see if vilazodone can cause other pregnancy-related problems, such as preterm delivery (birth before week 37) or low birth weight (weighing less than 5 pounds, 8 ounces [2500 grams] at birth).
It is not known if the use of vilazodone during pregnancy can cause withdrawal symptoms in a baby after birth. The use of other antidepressants during pregnancy has been associated with temporary symptoms in some newborns after birth. These symptoms are sometimes referred to as withdrawal. Symptoms may include jitteriness, increased muscle tone, irritability, changes in sleep patterns, tremors, trouble eating, and trouble breathing. These symptoms are usually mild and go away on their own. Some babies may need to stay in a special care nursery for several days. Not all babies exposed to an antidepressant will have these symptoms. It is important that your doctor know you are taking vilazodone so that if symptoms occur your baby can get the best care.
Studies have not been done to see if vilazodone use in pregnancy can cause behavior or learning issues for the child.
Breastfeeding
It is not known if vilazodone gets into breast milk or causes side effects for a baby who receives the milk. The benefit of continuing vilazodone while breastfeeding may outweigh the risks of an untreated mental health condition or the risks of not breastfeeding. Your doctor can talk with you about vilazodone and what treatment is best for you. Be sure to talk to your doctor about all of your breastfeeding questions.
Vilazodone dosage
Adult Dose for Depression
- Initial dose: 10 mg orally once a day for 7 days, followed by 20 mg orally once a day for an additional 7 days
- Maintenance dose: 40 mg orally once a day
Take vilazodone with food 147.
Missed Dose
If you miss a dose of vilazodone, take it as soon as possible. However, if it is almost time for your next dose, skip the missed dose and go back to your regular dosing schedule. Do not double doses.
Vilazodone side effects
Vilazodone may cause side effects. Tell your doctor if any of these symptoms are severe or do not go away:
- dry mouth
- increased appetite
- heartburn
- gas
- dizziness
- pain, burning, or tingling in the hands or feet
- uncontrollable shaking of a part of the body
- unusual dreams
- tiredness
- joint pain
- sexual problems in males; decreased sex drive, inability to get or keep an erection, or delayed or absent ejaculation
- sexual problems in females; decreased sex drive, or delayed orgasm or unable to have an orgasm
Some side effects can be serious. If you experience any of these symptoms or those listed in the IMPORTANT WARNING or SPECIAL PRECAUTIONS sections, see your doctor immediately:
- rash
- hives
- swelling
- difficulty breathing
- loss of consciousness
- seizures
- fever, sweating, confusion, fast or irregular heartbeat, and severe muscle stiffness or twitching, agitation, hallucinations, loss of coordination, nausea, vomiting, or diarrhea
- unusual bleeding or bruising
- nosebleeds
- small red or purple dots on the skin
- hallucinations (seeing things or hearing voices that do not exist)
- headache
- difficulty concentrating
- memory problems
- weakness
- problems with coordination
- increased falls
- fainting
Vilazodone may cause other side effects. See your doctor if you have any unusual problems while taking Vilazodone.
Serotonin syndrome
Serotonin syndrome also called serotonin toxicity is a potentially life threatening condition due to excessive serotonin in your body 174, 175. Serotonin syndrome most often occurs when two medicines that affect the body’s level of serotonin are taken together at the same time. The medicines cause too much serotonin to be released or to remain in the brain area. Serotonin syndrome (serotonin toxicity) can occur when you increase the dose of such a drug or add a new drug to your regimen. Certain illegal drugs and dietary supplements also are associated with serotonin syndrome. For example, you can develop serotonin syndrome if you take migraine medicines called triptans together with antidepressants called selective serotonin reuptake inhibitors (SSRIs), and selective serotonin/norepinephrine reuptake inhibitors (SSNRIs).
Common selective serotonin reuptake inhibitors (SSRIs) include:
- Citalopram (Celexa)
- Escitalopram (Lexapro)
- Fluoxetine (Prozac)
- Paroxetine (Paxil)
- Sertraline (Zoloft)
Selective serotonin/norepinephrine reuptake inhibitors (SSNRIs) include:
- Desvenlafaxine (Pristiq)
- Duloxetine (Cymbalta)
- Levomilnacipran (Fetzima)
- Milnacipran (Savella)
- Venlafaxine (Effexor)
Common triptans include:
- Almotriptan (Axert)
- Eletriptan (Relpax)
- Frovatriptan (Frova)
- Naratriptan (Amerge)
- Rizatriptan (Maxalt)
- Sumatriptan (Imitrex)
- Zolmitriptan (Zomig)
If you take these medicines, be sure to read the warning on the packaging. It tells you about the potential risk of serotonin syndrome. However, do not stop taking your medicine. Talk to your doctor about your concerns first.
Serotonin syndrome is more likely to occur when you first start taking or increase the dose of the medicine..
Older antidepressants called monoamine oxidase inhibitors (MAOIs) can also cause serotonin syndrome when combined with the medicines described above, as well as meperidine (Demerol, a painkiller), fentanyl, dextromethorphan (cough medicine), and others.
Drugs of abuse, such as ecstasy, LSD, opiates, cocaine, methamphetamine and amphetamines have also been associated with serotonin syndrome.
Serotonin is a chemical your body produces that’s needed for your nerve cells and brain to function. But too much serotonin causes symptoms that can range from mild (shivering and diarrhea) to severe (muscle rigidity, fever and seizures). Severe serotonin syndrome can be fatal if not treated.
Many medications can cause serotonin toxicity, and drug interactions are an important factor. Serotonin syndrome should be suspected in patients taking serotonergic drugs who present with autonomic or mental status changes and neurological findings. The findings of clonus, ocular clonus, hyperreflexia, and hypertonicity should prompt a doctor evaluation and medication review.
People with serotonin syndrome will usually stay in the hospital for at least 24 hours for close observation. Treatment is based on severity and focuses on prompt cessation of offending agents, treatment of hyperthermia, and use of benzodiazepines to decrease hypertonicity and neurological excitability.
Serotonin syndrome treatment may include:
- Benzodiazepine medicines to decrease agitation, seizure-like movements, and muscle stiffness
- Cyproheptadine (Periactin), a drug that blocks serotonin production
- Intravenous (through the vein) fluids
- Stopping medicines that caused serotonin syndrome
- In life-threatening cases, medicines that keep the muscles still (paralyze them), and a temporary breathing tube and breathing machine will be needed to prevent further muscle damage.
Milder forms of serotonin syndrome may go away within a day of stopping the medications that cause symptoms and, sometimes, taking drugs that block serotonin. The use of serotonin (5-HT) antagonists should be considered in moderate and severe cases. Increased awareness and monitoring of patients beginning treatment with antidepressants can decrease the risk of worsening anxiety, agitation, and possibly suicide.
Serotonin syndrome key points
- Serotonin syndrome is not an idiosyncratic drug reaction, but a predictable response to elevated serotonin levels. Medications that affect any of the steps in serotonin metabolism or regulation can provoke toxicity.
- Antidepressants are frequently implicated in serotonin syndrome. Interactions with other medications, such as common over-the-counter products (eg, dextromethorphan), can cause serious toxicity.
- Symptoms can occur within 6 to 8 hours of initiating or increasing the dosage of serotonergic medications. Drugs with long half-lives can interact several weeks after discontinuation.
- Treatment is based on the severity of the presentation. Many cases will be self-limited if the medications are stopped.
What does serotonin syndrome feel like?
Signs and symptoms of serotonin syndrome include:
- Agitation or restlessness
- Confusion
- Sweating
- Tremors
- Nausea
- Vomiting
- Raised body temperature
- Restlessness
- Overactive reflexes
- Fast heart rate and high blood pressure
- Dilated pupils
- Loss of muscle coordination or twitching muscles
- Muscle rigidity
- Heavy sweating
- Diarrhea
- Headache
- Shivering
- Goose bumps
Severe serotonin syndrome can be life-threatening. Signs and symptoms include:
- High fever
- Seizures
- Irregular heartbeat
- Unconsciousness
Symptoms of serotonin syndrome typically occur within hours of taking a drug. Individuals often experience an increased heart rate, restlessness, and confusion. The individual may experience heavy sweating, muscle twitching or rigidity, dilation of the pupils, shivering, headaches, and diarrhea. Symptoms often last for 1 to 3 days before they resolve. More severe cases of serotonin syndrome may involve high fever, irregular heartbeats, seizures, and loss of consciousness.
- If you believe that you or someone around you may be experiencing serotonin syndrome, after starting a new drug or increasing the dose of a drug you’re already taking, call your doctor right away or go to the emergency room. If you have severe or rapidly worsening symptoms, seek emergency treatment immediately. Doctors will check the drug levels, look for signs of infection, and monitor your vital signs. Treatment depends on the severity and type of symptoms. While some individuals will not require treatment, others may require hospitalization. If you or your child is experiencing serotonin syndrome, talk to your doctor about the most current treatment options.
Figure 6. Serotonin syndrome
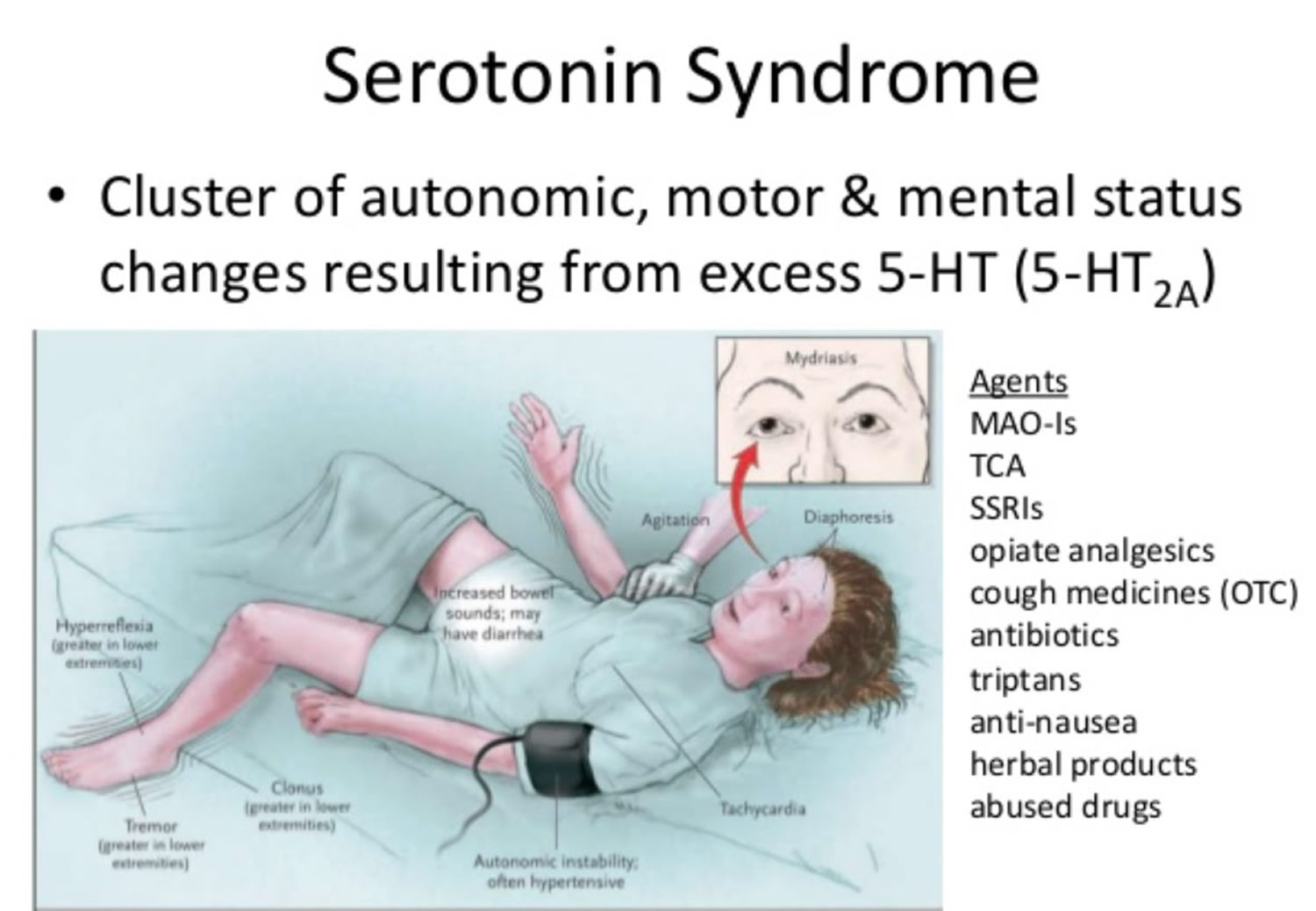
How long does serotonin syndrome last?
People may get slowly worse and can become severely ill if not quickly treated. Untreated, serotonin syndrome can be deadly. With treatment, symptoms usually go away in less than 24 hours.
Milder forms of serotonin syndrome usually go away within 24 to 72 hours of stopping medications that increase serotonin, and by taking medications to block the effects of serotonin already in your system if they’re needed.
However, symptoms of serotonin syndrome caused by some antidepressants could take several weeks to go away completely. These medications remain in your system longer than do other medications that can cause serotonin syndrome.
Serotonin syndrome long term effects
Serotonin syndrome generally doesn’t cause any problems once serotonin levels are back to normal.
Uncontrolled muscle spasms can cause severe muscle breakdown. The products produced when the muscles break down are released into the blood and eventually go through the kidneys. This can cause severe kidney damage if serotonin syndrome isn’t recognized and treated properly.
If left untreated, severe serotonin syndrome can lead to unconsciousness and death.
Serotonin syndrome causes
Excessive accumulation of serotonin in your body creates the symptoms of serotonin syndrome.
Under normal circumstances, nerve cells in your brain and spinal cord (central nervous system) produce serotonin that helps regulate your attention, behavior and body temperature.
Other nerve cells in your body, primarily in your intestines, also produce serotonin. In these other areas, serotonin plays a role in regulating your digestive process, blood flow and breathing.
Although it’s possible that taking just one drug that increases serotonin levels can cause serotonin syndrome in susceptible individuals, this condition occurs most often when you combine certain medications.
For example, serotonin syndrome may occur if you take an antidepressant with a migraine medication. It may also occur if you take an antidepressant with an opioid pain medication.
Another cause of serotonin syndrome is intentional overdose of antidepressant medications.
A number of over-the-counter and prescription drugs may be associated with serotonin syndrome, especially antidepressants. Illicit drugs and dietary supplements also may be associated with the condition. These drugs and supplements include:
- Selective serotonin reuptake inhibitors (SSRIs), antidepressants such as citalopram (Celexa), fluoxetine (Prozac, Sarafem), fluvoxamine, paroxetine (Paxil) and sertraline (Zoloft)
- Serotonin and norepinephrine reuptake inhibitors (SNRIs), antidepressants such as trazodone, duloxetine (Cymbalta) and venlafaxine (Effexor)
- Bupropion (Wellbutrin, Zyban), an antidepressant and tobacco-addiction medication
- Tricyclic antidepressants, such as amitriptyline and nortriptyline (Pamelor)
- Monoamine oxidase inhibitors (MAOIs), antidepressants such as isocarboxazid (Marplan) and phenelzine (Nardil)
- Anti-migraine medications such as triptans (Axert, Amerge, Imitrex), carbamazepine (Tegretol) and valproic acid (Depakene)
- Pain medications such as opioid pain medications including codeine (Tylenol with codeine), fentanyl (Duragesic), hydrocodone meperidine (Demerol), oxycodone (Oxycontin, Percocet, Percodan) and tramadol (Ultram).
- Lithium (Lithobid), a mood stabilizer
- Illicit drugs, including LSD, Ecstasy, cocaine and amphetamines
- Herbal supplements, including St. John’s wort, ginseng and nutmeg
- Over-the-counter cough and cold medications containing dextromethorphan (Delsym, Mucinex DM, others)
- Anti-nausea medications such as granisetron, metoclopramide (Reglan), droperidol (Inapsine) and ondansetron (Zofran)
- Linezolid (Zyvox), an antibiotic
- Ritonavir (Norvir), an anti-retroviral medication used to treat HIV/AIDS
Risk factors for serotonin syndrome
Some people are more susceptible to the drugs and supplements that cause serotonin syndrome than are others, but the condition can occur in anyone.
You’re at increased risk of serotonin syndrome if:
- You recently started taking or increased the dose of a medication known to increase serotonin levels.
- You take more than one drug known to increase serotonin levels.
- You take herbal supplements known to increase serotonin levels.
- You use an illicit drug known to increase serotonin levels.
Serotonin syndrome prevention
Taking more than one serotonin-related medication or increasing your dose of a serotonin-related medication increases your risk of serotonin syndrome.
Talk to your doctor about possible risks. Don’t stop taking any such medications on your own. If your doctor prescribes a new medication, make sure he or she knows about all the other medications you’re taking, especially if you receive prescriptions from more than one doctor.
If you and your doctor decide the benefits of combining certain serotonin-level-affecting drugs outweigh the risks, be alert to the possibility of serotonin syndrome.
Serotonin syndrome signs and symptoms
Serotonin syndrome symptoms usually occur within several hours of taking a new drug or increasing the dose of a drug you’re already taking.
Signs and symptoms of serotonin syndrome include:
- Agitation or restlessness
- Confusion
- Rapid heart rate and high blood pressure
- Dilated pupils
- Loss of muscle coordination or twitching muscles
- Muscle rigidity
- Heavy sweating
- Diarrhea
- Headache
- Shivering
- Goose bumps
Severe serotonin syndrome can be life-threatening. Signs and symptoms include:
- High fever
- Seizures
- Irregular heartbeat
- Unconsciousness
Table 4. Serotonin syndrome signs and symptoms
| SERIOUSNESS | AUTONOMIC SIGNS | NEUROLOGICAL SIGNS | MENTAL STATUS | OTHER |
|---|---|---|---|---|
| Mild | Afebrile or low-grade fever Tachycardia Mydriasis Diaphoresis or shivering | Intermittent tremor Akathisia Myoclonus Mild hyperreflexia | Restlessness Anxiety | |
| Moderate | Increased tachycardia Fever (up to 41°C) Diarrhea with hyperactive bowel sounds Diaphoresis with normal skin colour | Hyperreflexia Inducible clonus Ocular clonus (slow continuous lateral eye movements) Myoclonus | Easily startled Increased confusion Agitation and hypervigilance | Rhabdomyolysis Metabolic acidosis Renal failure Disseminated intravascular coagulopathy (secondary to hyperthermia) |
| Severe | Temperature often more than 41°C (Secondary to increased tone) | Increased muscle tone(lower limb > upper) Spontaneous clonus Substantial myoclonus or hyperreflexia | Delirium Coma | As above |
Serotonin syndrome complications
Uncontrolled muscle spasms can cause severe muscle breakdown. The products produced when the muscles break down are released into the blood and eventually go through the kidneys. This can cause severe kidney damage if not recognized and treated properly.
Untreated, serotonin syndrome can be deadly. With treatment, symptoms usually go away in less than 24 hours. Permanent organ damage may result, even with treatment.
Serotonin syndrome diagnosis
No single test can confirm a serotonin syndrome diagnosis. Your doctor will diagnose the condition by ruling out other possibilities.
Your doctor will likely begin by asking about your symptoms, medical history and any medications you’re taking. Your doctor will also conduct a physical examination.
To be diagnosed with serotonin syndrome, the person must have been taking a drug that changes the body’s serotonin level (serotonergic drug) and have at least 3 of the following signs or symptoms:
- Agitation
- Diarrhea
- Heavy sweating not due to activity
- Fever
- Mental status changes, such as confusion or hypomania
- Muscle spasms (myoclonus)
- Overactive reflexes (hyperreflexia)
- Shivering
- Tremor
- Uncoordinated movements (ataxia)
Serotonin syndrome is not diagnosed until all other possible causes have been ruled out. This may include infections, intoxication, metabolic and hormone problems, and drug or alcohol withdrawal. Some symptoms of SS can mimic those due to an overdose of cocaine, lithium, or an monoamine oxidase inhibitor (MAOI).
If a person has just started taking or increased the dosage of a tranquilizer (neuroleptic drug), other conditions such as neuroleptic malignant syndrome (NMS) will be considered.
To make sure your symptoms are caused by serotonin syndrome and not due to another cause, your doctor may use tests to:
- Measure levels of any drugs you’re using
- Check for signs of infection
- Check body functions that may be affected by serotonin syndrome
A number of conditions can cause symptoms similar to those of serotonin syndrome. Minor symptoms can be caused by numerous conditions. Causes of moderate and severe symptoms include:
- Anticholinergic syndrome, malignant hyperthermia and neuroleptic malignant syndrome (NMS), serious conditions caused by certain medications
- An overdose of illegal drugs, antidepressant medications or other medications that increase serotonin levels
- Damage (toxicity) associated with illegal drug use
- Severe alcohol withdrawal
Your doctor may order tests to rule out other causes of your symptoms. Tests may include:
- Blood and urine tests
- Chest X-ray
- Computerized tomography
- Spinal tap (lumbar puncture).
Serotonin syndrome treatment
Treatment of serotonin syndrome depends on the severity of your symptoms.
- If your symptoms are minor, a visit to the doctor and stopping the medication causing the problem may be enough.
- If you have symptoms that concern your doctor, you may need to go to the hospital. Your doctor may have you stay in the hospital for several hours to make sure your symptoms are improving.
- If you have severe serotonin syndrome, you’ll need intensive treatment in a hospital.
Depending on your symptoms, you may receive the following treatments:
- Muscle relaxants. Benzodiazepines, such as diazepam (Valium) or lorazepam (Ativan), can help control agitation, seizures and muscle stiffness.
- Serotonin-production blocking agents. If other treatments aren’t working, medications such as cyproheptadine can help by blocking serotonin production.
- Oxygen and intravenous (IV) fluids. Breathing oxygen through a mask helps maintain normal oxygen levels in your blood, and IV fluids are used to treat dehydration and fever.
- Drugs that control heart rate and blood pressure. These may include esmolol (Brevibloc) or nitroprusside (Nitropress), to reduce a high heart rate or high blood pressure. If your blood pressure is too low, your doctor may give you phenylephrine (Neo-Synephrine) or epinephrine (Adrenalin, Epipen).
- A breathing tube and machine and medication to paralyze your muscles. You may need a breathing tube and medication to paralyze your muscles if you have a high fever.
Prompt recognition of toxicity and discontinuation of offending medications are most important. Many cases of serotonin syndrome are self-limiting if medications are stopped early. The mortality of severe serotonin syndrome is estimated to range from 2% to 12%. The Toxic Exposure Surveillance System in the United States reported 93 deaths due to serotonin syndrome in 2002 192.
Treatment is guided by the severity of presentation. There are currently no randomized placebo-controlled trials to guide pharmacologic treatment of serotonin syndrome, although case reports and series suggest several medications that are beneficial in managing the condition.
Supportive care, including intravenous fluids, is indicated in patients with vital sign abnormalities. Neurological symptoms, including serious myoclonus and hyperreflexia, are sometimes treated with benzodiazepines, but there is little evidence for this approach. Hyperthermia should be aggressively managed with external cooling, hydration, and benzodiazepines (eg, diazepam, lorazepam). Patients with a temperature higher than 105.8 °F (41°C) should be intubated with induced neuromuscular paralysis. There is a limited role for traditional antipyretics, as the mechanism of serotonin syndrome is due to muscle tone rather than central thermoregulation. Patient agitation secondary to delirium should be managed using nonpharmacologic strategies and benzodiazepines, when appropriate. Physical restraints should be avoided, as they can increase hyperthermia, lactic acidosis, and rhabdomyolysis 193.
The antihistamine cyproheptadine, which is also a serotonin inhibitor, should be considered in moderate cases and is recommended in severe cases, despite a lack of randomized controlled trial evidence 193. It is available only as an oral preparation; the initial dose is 12 mg; the dosage is then adjusted to 2 mg every 2 hours until symptoms improve 194.
Some antipsychotics have serotonin antagonist effects and are sometimes used 193. Sublingual olanzapine and intramuscular chlorpromazine (50 to 100 mg) are options 195. Chlorpromazine can cause serious hypotension and should be avoided in severe cases with shock.
Caution is required in using antipsychotics for treating serotonin syndrome, as neuroleptic malignant syndrome (NMS) can be misdiagnosed as serotonin syndrome. Also, case reports have implicated antipsychotics as precipitants of serotonin syndrome in individuals receiving concomitant serotonergic medications 196.
Dantrolene, a skeletal muscle relaxant used for treatment of neuroleptic malignant syndrome (NMS), has been reported to improve symptoms of serotonin syndrome in a case series 197; however, it has also been implicated in the development of serotonin toxicity and is not generally recommended 191. Propranolol, which has serotonin antagonist activity and a long half-life, can potentiate hypotension and make improvement in tachycardia a less effective strategy for monitoring response to treatment 191.
Serotonin syndrome prognosis
Serotonin syndrome prognosis (outlook) varies depending on the degree of serotonin toxicity, which is primarily attributable to the dose and type of serotonergic agent 175. Serotonin syndrome typically resolves rapidly and safely when quickly treated 175. Generally, full recovery from serotonin syndrome is expected after stabilization. However, monoamine oxidase inhibitors (MAOIs) are associated with the most severe cases of serotonin syndrome, but intentional overdoses of any serotonergic drug can also result in severe presentations. Mild serotonin syndrome cases may only require observation, while severe serotonin syndrome cases require intensive care. The duration of serotonin syndrome depends on a few factors but is most impacted by the half-life of the culprit medications. When the altered mental status resolves, vital signs are normal, and a normal neurological examination is elicited (no clonus or hyperreflexia), the patient is safe for discharge. After serotonin syndrome resolves, a comprehensive assessment is needed to assess the risks and benefits of resuming the previous serotonergic regimen to prevent another episode of serotonin syndrome 198, 199.
- Bakshi A, Tadi P. Biochemistry, Serotonin. [Updated 2022 Oct 5]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK560856[↩][↩][↩][↩][↩][↩][↩][↩][↩]
- Smith C, Smith M, Cunningham R, Davis S. Recent Advances in Antiemetics: New Formulations of 5-HT3 Receptor Antagonists in Adults. Cancer Nurs. 2020 Jul/Aug;43(4):E217-E228. doi: 10.1097/NCC.0000000000000694[↩][↩][↩][↩]
- David DJ, Gardier AM. Les bases de pharmacologie fondamentale du système sérotoninergique : application à la réponse antidépressive [The pharmacological basis of the serotonin system: Application to antidepressant response]. Encephale. 2016 Jun;42(3):255-63. French. doi: 10.1016/j.encep.2016.03.012[↩][↩]
- Hannon J., Hoyer D. (2008). Molecular biology of 5-HT receptors. Behav. Brain Res. 195, 198–213. 10.1016/j.bbr.2008.03.020[↩]
- Sharp T, Barnes NM. Central 5-HT receptors and their function; present and future. Neuropharmacology (2020) 177:108155. doi: 10.1016/j.neuropharm.2020.108155[↩]
- Celli J, Rappold G, Niesler B. The human serotonin type 3 receptor gene (HTR3A-e) allelic variant database. Hum Mutat (2017) 38:137–47. doi: 10.1002/humu.23136[↩]
- Lv J, Liu F. The Role of Serotonin beyond the Central Nervous System during Embryogenesis. Front Cell Neurosci. 2017 Mar 13;11:74. doi: 10.3389/fncel.2017.00074[↩][↩][↩]
- Bonnin A, Levitt P. Fetal, maternal and placental sources of serotonin and new implications for developmental programming of the brain. Neuroscience (2011) 197:1–7. doi: 10.1016/j.neuroscience.2011.10.005[↩]
- Perić M, Bečeheli I, Čičin-Šain L, Desoye G, Štefulj J. Serotonin system in the human placenta – the knowns and unknowns. Front Endocrinol (Lausanne). 2022 Dec 1;13:1061317. doi: 10.3389/fendo.2022.1061317[↩]
- Dale GL. Coated-platelets: An emerging component of the procoagulant response. J Thromb Haemost (2005) 3:2185–92. doi: 10.1111/j.1538-7836.2005.01274.x[↩]
- Walther DJ, Peter J-U, Winter S, Höltje M, Paulmann N, Grohmann M, et al. Serotonylation of small GTPases is a signal transduction pathway that triggers platelet alpha-granule release. Cell (2003) 115:851–62. doi: 10.1016/s0092-8674(03)01014-6[↩]
- Paulmann N, Grohmann M, Voigt JP, Bert B, Vowinckel J, Bader M, et al. Intracellular serotonin modulates insulin secretion from pancreatic β-cells by protein serotonylation. PloS Biol (2009) 7:1–10. doi: 10.1371/journal.pbio.1000229[↩]
- Farrelly LA, Thompson RE, Zhao S, Lepack AE, Lyu Y, Bhanu NV, et al. Histone serotonylation is a permissive modification that enhances TFIID binding to H3K4me3. Nature (2019) 567:535–9. doi: 10.1038/s41586-019-1024-7[↩]
- Negri L. Vittorio Erspamer (1909-1999) [Vittorio Erspamer (1909-1999)]. Med Secoli. 2006;18(1):97-113. Italian.[↩]
- Vialli M, Erspamer V. Ricerche sul secreto delle cellule enterocromaffini. Z fur Zellforschung und Mikrosk Anat (1937) 27:81–99. doi: 10.1007/bf00391792[↩]
- Rapport MM, ArdaAlden G, Page IH. Serum vasoconstrictor (Serotonin). J Biol Chem (1948) 176:1243–51. doi: 10.1016/s0021-9258(18)57137-4[↩]
- Rapport M. M., Green A. A., Page I. H. (1948). Serum vasoconstrictor (Serotonin). IV. isolation and characterization. J. Biol. Chem. 176, 1243–1251. https://www.jbc.org/article/S0021-9258(18)57137-4/pdf[↩]
- Gershon MD, Tack J. The serotonin signaling system: from basic understanding to drug development for functional GI disorders. Gastroenterology. 2007 Jan;132(1):397-414. doi: 10.1053/j.gastro.2006.11.002[↩]
- Lechin F, van der Dijs B, Orozco B, Lechin M, Lechin AE. Increased levels of free serotonin in plasma of symptomatic asthmatic patients. Ann Allergy Asthma Immunol (1996) 77:245–53. doi: 10.1016/S1081-1206(10)63263-2[↩]
- Slominski AT, Zmijewski MA, Cezary S, Zbytek B, Slominski RM, Steketee JD. Sensing the environment: Regulation of local and globalhomeostasis by the skin neuroendocrine system. Adv Anat Embryol Cell Biol (2012) 212:1–98. doi: 10.1007/978-3-642-19683-6_1[↩]
- Spohn SN, Mawe GM. Non-conventional features of peripheral serotonin signalling-the gut and beyond. Nat Rev Gastroenterol Hepatol (2017) 14:412–20. doi: 10.1038/nrgastro.2017.51[↩]
- Yabut JM, Desjardins EM, Chan EJ, Day EA, Leroux JM, Wang B, et al. Genetic deletion of mast cell serotonin synthesis prevents the development of obesity and insulin resistance. Nat Commun (2020) 11:463. doi: 10.1038/s41467-019-14080-7[↩]
- Berger M, Gray JA, Roth BL. The expanded biology of serotonin. Annu Rev Med (2009) 60:355–66. doi: 10.1146/annurev.med.60.042307.110802[↩][↩]
- Azmitia EC. Evolution of serotonin: Sunlight to suicide. Amsterdam, Netherlands: Elsevier B.V; (2010). doi: 10.1016/s1569-7339(10)70069-2[↩]
- Jacobs BL, Azmitia EC. Structure and function of the brain serotonin system. Physiol Rev (1992) 72:165–229. doi: 10.1152/physrev.1992.72.1.165[↩]
- Yabut JM, Crane JD, Green AE, Keating DJ, Khan WI, Steinberg GR. Emerging roles for serotonin in regulating metabolism: New implications for an ancient molecule. Endocr Rev (2019) 40:1092–107. doi: 10.1210/er.2018-00283[↩]
- Whitaker-Azmitia PM, Druse M, Walker P, Lauder JM. Serotonin as a developmental signal. Behav Brain Res (1996) 73:19–29. doi: 10.1016/0166-4328(96)00071-x[↩]
- Papadimas GK, Tzirogiannis KN, Mykoniatis MG, Grypioti AD, Manta GA, Panoutsopoulos GI. The emerging role of serotonin in liver regeneration. Swiss Med Weekly (2012) 142:1–6. doi: 10.4414/smw.2012.13548[↩]
- Gaspar P, Cases O, Maroteaux L. The developmental role of serotonin: News from mouse molecular genetics. Nat Rev Neurosci (2003) 4:1002–12. doi: 10.1038/nrn1256[↩]
- Sodhi MSK, Sanders-Bush E. Serotonin and brain development. Int Rev Neurobiol (2004) 59:111–74. doi: 10.1016/S0074-7742(04)59006-2[↩]
- Azouzi S, Santuz H, Morandat S, Pereira C, Côté F, Hermine O, et al. Antioxidant and membrane binding properties of serotonin protect lipids from oxidation. Biophys J (2016) 112:1863–73. doi: 10.1016/j.bpj.2017.03.037[↩]
- The role of serotonin in human mood and social interaction. Insight from altered tryptophan levels. Young SN, Leyton M. Pharmacol Biochem Behav. 2002 Apr; 71(4):857-65. https://www.ncbi.nlm.nih.gov/pubmed/11888576/[↩]
- Serotonin and drug-induced therapeutic responses in major depression, obsessive-compulsive and panic disorders. Blier P, de Montigny C. Neuropsychopharmacology. 1999 Aug; 21(2 Suppl):91S-98S. https://www.ncbi.nlm.nih.gov/pubmed/10432494/[↩]
- The spectrum of behaviors influenced by serotonin. Lucki I. Biol Psychiatry. 1998 Aug 1; 44(3):151-62. https://www.ncbi.nlm.nih.gov/pubmed/9693387/[↩]
- Synaptic and extrasynaptic secretion of serotonin. Cell Mol Neurobiol. 2005 Mar;25(2):297-312. https://link.springer.com/article/10.1007%2Fs10571-005-3061-z[↩][↩]
- A High-Resolution In Vivo Atlas of the Human Brain’s Serotonin System. Beliveau V, Ganz M, Feng L, Ozenne B, Højgaard L, Fisher PM, Svarer C, Greve DN, Knudsen GM. J Neurosci. 2017 Jan 4; 37(1):120-128.[↩]
- Kitson SL. 5-hydroxytryptamine (5-HT) receptor ligands. Curr Pharm Des. 2007;13(25):2621-37. doi: 10.2174/138161207781663000[↩]
- Affective style, psychopathology, and resilience: brain mechanisms and plasticity. Davidson RJ. Am Psychol. 2000 Nov; 55(11):1196-214. http://dx.doi.org/10.1037/0003-066X.55.11.1196[↩]
- Molliver M E . Serotonergic neuronal systems: What their anatomic organization tells us about function. J. Clin. Psychopharmacol. 1987;7 (6 Suppl.):3s–23s.[↩][↩]
- Frazer A, Hensler JG. Serotonin. In: Siegel GJ, Agranoff BW, Albers RW, et al., editors. Basic Neurochemistry: Molecular, Cellular and Medical Aspects. 6th edition. Philadelphia: Lippincott-Raven; 1999. Available from: https://www.ncbi.nlm.nih.gov/books/NBK28150[↩][↩][↩][↩][↩][↩][↩][↩]
- Lovenberg W., Jequier E., Sjoerdsma A. (1967). Tryptophan hydroxylation: measurement in pineal gland, brainstem, and carcinoid tumor. Science 155, 217–219. 10.1126/science.155.3759.217[↩][↩]
- Ichiyama A., Nakamura S., Nishizuka Y., Hayaishi O. (1970). Enzymic studies on the biosynthesis of serotonin in mammalian brain. J. Biol. Chem. 245, 1699–1709. https://www.jbc.org/article/S0021-9258(19)77149-X/pdf[↩]
- Côté F., Thévenot E., Fligny C., Fromes Y., Darmon M., Ripoche M.-A., et al. (2003). Disruption of the nonneuronal tph1 gene demonstrates the importance of peripheral serotonin in cardiac function. Proc. Natl. Acad. Sci. U S A 100, 13525–13530. 10.1073/pnas.2233056100[↩]
- Walther DJ, Peter J-U, Bashammakh S, Hörtnagl H, Voits M, Fink H, et al. Synthesis of serotonin by a second tryptophan hydroxylase isoform. Science (2003) 299:76. doi: 10.1126/science.1078197[↩]
- Zill P., Büttner A., Eisenmenger W., Müller J., Moller H. J., Bondy B. (2009). Predominant expression of tryptophan hydroxylase 1 mRNA in the pituitary: a postmortem study in human brain. Neuroscience 159, 1274–1282. 10.1016/j.neuroscience.2009.01.006[↩]
- Zhang P., He Q., Chen D., Liu W., Wang L., Zhang C., et al. (2015). G protein-coupled receptor 183 facilitates endothelial-to-hematopoietic transition via Notch1 inhibition. Cell Res. 25, 1093–1107. 10.1038/cr.2015.109[↩]
- Lv J., Wang L., Gao Y., Ding Y. Q., Liu F. (2017). 5-hydroxytryptamine synthesized in the aorta-gonad-mesonephros regulates hematopoietic stem and progenitor cell survival. J. Exp. Med. 214, 529–545. 10.1084/jem.20150906[↩][↩]
- Christie A. E., Fontanilla T. M., Roncalli V., Cieslak M. C., Lenz P. H. (2014). Identification and developmental expression of the enzymes responsible for dopamine, histamine, octopamine and serotonin biosynthesis in the copepod crustacean Calanus finmarchicus. Gen. Comp. Endocrinol. 195, 28–39. 10.1016/j.ygcen.2013.10.003[↩]
- Shih J. C., Chen K., Ridd M. J. (1999). Monoamine oxidase: from genes to behavior. Annu. Rev. Neurosci. 22, 197–217. 10.1146/annurev.neuro.22.1.197[↩][↩][↩]
- Singh S., Johnson P. I., Javed A., Gray T. S., Lonchyna V. A., Wurster R. D. (1999). Monoamine- and histamine-synthesizing enzymes and neurotransmitters within neurons of adult human cardiac ganglia. Circulation 99, 411–419. 10.1161/01.cir.99.3.411[↩][↩][↩]
- Heninger GR, Delgado PL, Charney DS. The revised monoamine theory of depression: a modulatory role for monoamines, based on new findings from monoamine depletion experiments in humans. Pharmacopsychiatry. 1996 Jan;29(1):2-11. doi: 10.1055/s-2007-979535[↩]
- Young SN, Gauthier S. Effect of tryptophan administration on tryptophan, 5-hydroxyindoleacetic acid, and indoleacetic acid in human lumbar and cisternal cerebrospinal fluid. J Neurol Neurosurg Psychiatry 1981;44:323-7. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC490955/pdf/jnnpsyc00060-0059.pdf[↩][↩]
- Young SN. The clinical psychopharmacology of tryptophan. In: Wurtman RJ, Wurtman JJ, editors. Food constituents affecting normal and abnormal behaviors. New York: Raven Press; 1986. p. 49-88.[↩][↩][↩]
- aan het Rot M, Moskowitz DS, Pinard G, et al. Social behaviour and mood in everyday life: effects of tryptophan in quarrelsome individuals. J Psychiatry Neurosci 2006;31:253-62. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1488902/[↩][↩]
- Wurtman RJ, Hefti F, Melamed E. Precursor control of neurotransmitter synthesis. Pharmacol Rev 1980;32:315-35. https://www.ncbi.nlm.nih.gov/pubmed/6115400[↩][↩]
- Young SN. How to increase serotonin in the human brain without drugs. Journal of Psychiatry & Neuroscience : JPN. 2007;32(6):394-399. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2077351/[↩][↩][↩][↩][↩]
- Booij L, Merens W, Markus CR, et al. Diet rich in alpha-lactalbumin improves memory in unmedicated recovered depressed patients and matched controls. J Psychopharmacol 2006;20:526-35. https://www.ncbi.nlm.nih.gov/pubmed/16174675[↩][↩]
- Eaton M J , Gudehithlu K P , Quach T , Silvia C P , Hadjiconstantinou M , Neff N H . Distribution of aromatic l-amino acid decarboxylase mRNA in mouse brain by in situ hybridization histology. J. Comp. Neurol. 1993;337:640–654[↩]
- Cortes R , Mengod G , Celada P , Artigas F . p-Chlorophenylalanine increases tryptophan-5-hydroxylase mRNA levels in the rat dorsal raphe: A time course study using in situ hybridization. J. Neurochem. 1993;60:761–764.[↩]
- Boadle-Biber M C . Regulation of serotonin synthesis. Prog. Biophys. Mol. Biol. 1993;60:1–15.[↩]
- Tamir H , Liu K , Hsiung S , Adlersberg M , Gershon M D . Serotonin binding protein: Synthesis, secretion, and recycling. J. Neurochem. 1994;63:97–107.[↩]
- Sivasubramaniam SD, Finch CC, Rodriguez MJ, Mahy N, Billett EE. A comparative study of the expression of monoamine oxidase-a and -b mRNA and protein in non-CNS human tissues. Cell Tissue Res (2003) 313:291–300. doi: 10.1007/s00441-003-0765-6[↩]
- Ramsay RR, Albreht A. Kinetics, mechanism, and inhibition of monoamine oxidase. J Neural Transm (2018) 125:1659–83. doi: 10.1007/s00702-018-1861-9[↩]
- Shih J C . Molecular basis of human MAO A and B. Neuropsychopharmacology. 1991;4:1–3.[↩]
- Cases O , Seif I , Grimsby J . et al. Aggressive behavior and altered amounts of brain serotonin and norepinephrine in mice lacking MAO A. Science. 1995;268:1763–1766.[↩]
- Carhart-Harris R, Nutt D. Serotonin and brain function: a tale of two receptors. Journal of Psychopharmacology (Oxford, England). 2017;31(9):1091-1120. doi:10.1177/0269881117725915. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5606297/[↩][↩][↩][↩]
- Is serotonin an upper or a downer? The evolution of the serotonergic system and its role in depression and the antidepressant response. Andrews PW, Bharwani A, Lee KR, Fox M, Thomson JA Jr. Neurosci Biobehav Rev. 2015 Apr; 51():164-88. https://www.ncbi.nlm.nih.gov/pubmed/25625874/[↩]
- The double edged sword of neural plasticity: increasing serotonin levels leads to both greater vulnerability to depression and improved capacity to recover. Branchi I. Psychoneuroendocrinology. 2011 Apr; 36(3):339-51. https://www.ncbi.nlm.nih.gov/pubmed/20875703/[↩]
- Serotonin in affective control. Dayan P, Huys QJ. Annu Rev Neurosci. 2009; 32:95-126.[↩]
- Serotoninergic regulation of emotional and behavioural control processes. Cools R, Roberts AC, Robbins TW. Trends Cogn Sci. 2008 Jan; 12(1):31-40.[↩]
- International Union of Pharmacology classification of receptors for 5-hydroxytryptamine (Serotonin). Hoyer D, Clarke DE, Fozard JR, Hartig PR, Martin GR, Mylecharane EJ, Saxena PR, Humphrey PP. Pharmacol Rev. 1994 Jun; 46(2):157-203.[↩]
- A comparison of the subsecond dynamics of neurotransmission of dopamine and serotonin. Jennings KA. ACS Chem Neurosci. 2013 May 15; 4(5):704-14.[↩]
- Serotonin revisited. Müller CP, Homberg JR. Behav Brain Res. 2015 Jan 15; 277():1-2. https://www.ncbi.nlm.nih.gov/pubmed/25311281/[↩]
- The origins of ‘5-HT and mechanisms of defence’ by Deakin and Graeff: a personal perspective. Deakin J. J Psychopharmacol. 2013 Dec; 27(12):1084-9.[↩]
- The role of serotonin in the regulation of patience and impulsivity. Miyazaki K, Miyazaki KW, Doya K. Mol Neurobiol. 2012 Apr; 45(2):213-24. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3311865/[↩]
- The double edged sword of neural plasticity: increasing serotonin levels leads to both greater vulnerability to depression and improved capacity to recover. Branchi I. Psychoneuroendocrinology. 2011 Apr; 36(3):339-51.[↩]
- Serotonin and stress coping. Puglisi-Allegra S, Andolina D. Behav Brain Res. 2015 Jan 15; 277():58-67. https://www.ncbi.nlm.nih.gov/pubmed/25108244/[↩]
- Graeff FG, Zangrossi H Jr. The dual role of serotonin in defense and the mode of action of antidepressants on generalized anxiety and panic disorders. Cent Nerv Syst Agents Med Chem 2010; 10: 207–217.[↩]
- Hamm AO, Richter J, Pané-Farré C, Westphal D, Wittchen H-U, Vossbeck-Elsebusch AN et al. Panic disorder with agoraphobia from a behavioral neuroscience perspective: Applying the research principles formulated by the Research Domain Criteria (RDoC) initiative. Psychophysiology 2016; 53: 312–322.[↩]
- American Psychiatric AssociationDSM-5 Task Force. Diagnostic and Statistical Manual of Mental Disorders: DSM-5, 5th edn, American Psychiatric Association: Washington, DC, USA, 2013; 947.[↩]
- Canli T, Congdon E, Gutknecht L, Constable RT, Lesch KP. Amygdala responsiveness is modulated by tryptophan hydroxylase-2 gene variation. J Neural Transm 2005; 112: 1479–1485.[↩]
- Gutknecht L, Jacob C, Strobel A, Kriegebaum C, Muller J, Zeng Y et al. Tryptophan hydroxylase-2 gene variation influences personality traits and disorders related to emotional dysregulation. Int J Neuropsychopharmacol 2007; 10: 309–320.[↩]
- Kim YK, Lee HJ, Yang JC, Hwang JA, Yoon HK. A tryptophan hydroxylase 2 gene polymorphism is associated with panic disorder. Behav Genet 2009; 39: 170–175.[↩]
- Preuss N, Salehi B, van der Veen JW, Shen J, Drevets WC, Hodgkinson C et al. Associations between prefrontal gamma-aminobutyric acid concentration and the tryptophan hydroxylase isoform 2 gene, a panic disorder risk allele in women. Int J Neuropsychopharmacol 2013; 16: 1707–1717.[↩]
- Johnson PL, Molosh A, Fitz SD, Arendt D, Deehan GA, Federici LM et al. Pharmacological depletion of serotonin in the basolateral amygdala complex reduces anxiety and disrupts fear conditioning. Pharmacol Biochem Behav 2015; 138: 174–179.[↩]
- Baratta MV, Kodandaramaiah SB, Monahan PE, Yao J, Weber MD, Lin PA et al. Stress enables reinforcement-elicited serotonergic consolidation of fear memory. Biol Psychiatry 2016; 79: 814–822.[↩]
- Waider J, Popp S, Lange MD, et al. Genetically driven brain serotonin deficiency facilitates panic-like escape behavior in mice. Translational Psychiatry. 2017;7(10):e1246-. doi:10.1038/tp.2017.209. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5682603/[↩]
- Sivolap YP. Antidepressanty: tseli i vozmozhnosti terapii [Antidepressants: the goals and possibilities of therapy]. Zh Nevrol Psikhiatr Im S S Korsakova. 2018;118(12):120-124. Russian. doi: 10.17116/jnevro2018118121120[↩]
- Dale G. L., Friese P., Batar P., Hamilton S. F., Reed G. L., Jackson K. W., et al. (2002). Stimulated platelets use serotonin to enhance their retention of procoagulant proteins on the cell surface. Nature 415, 175–179. 10.1038/415175a[↩]
- Ivetic N, Arnold DM, Smith JW, Huynh A, Kelton JG, Nazy I. A platelet viability assay (PVA) for the diagnosis of heparin-induced thrombocytopenia. Platelets. 2019;30(8):1017-1021. doi: 10.1080/09537104.2018.1562169[↩]
- Liu Y.-S., Yang M. (2006). The effect of 5-hydroxtryptamine on the regulation of megakaryocytopoiesis. Hematology 11, 53–56. 10.1080/10245330500322370[↩]
- Yang M., Srikiatkhachorn A., Anthony M., Chong B. H. (1996). Serotonin stimulates megakaryocytopoiesis via the 5-HT2 receptor. Blood Coagul. Fibrinolysis 7, 127–133. 10.1097/00001721-199603000-00004[↩]
- Ye J. Y., Liang E. Y., Cheng Y. S., Chan G. C., Ding Y., Meng F., et al. (2014). Serotonin enhances megakaryopoiesis and proplatelet formation via p-Erk1/2 and F-actin reorganization. Stem Cells 32, 2973–2982. 10.1002/stem.1777[↩]
- Weaver SR, Xie C, Charles JF, Hernandez LL. In utero and lactational exposure to the Selective Serotonin Reuptake Inhibitor fluoxetine compromises pup bones at weaning. Sci Rep. 2019 Jan 18;9(1):238. doi: 10.1038/s41598-018-36497-8[↩]
- Frost M, Andersen TE, Yadav V, Brixen K, Karsenty G, Kassem M. Patients with high-bone-mass phenotype owing to Lrp5-T253I mutation have low plasma levels of serotonin. J Bone Miner Res. 2010 Mar;25(3):673-5. doi: 10.1002/jbmr.44. Erratum in: J Bone Miner Res. 2010 Aug;25(8):1896.[↩]
- Zainol M, Sidi H, Kumar J, Das S, Ismail SB, Hatta MH, Baharuddin N, Ravindran A. Co-Morbid Erectile Dysfunction (ED) and Antidepressant Treatment in a Patient – A Management Challenge? Curr Drug Targets. 2019;20(2):182-191. doi: 10.2174/1389450118666170315110902[↩]
- Cowen PJ, Browning M. What has serotonin to do with depression? World Psychiatry. 2015;14(2):158-160. doi:10.1002/wps.20229. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4471964/[↩][↩]
- The biochemistry of affective disorders. Coppen A. Br J Psychiatry. 1967 Nov; 113(504):1237-64. https://www.cambridge.org/core/journals/the-british-journal-of-psychiatry/article/biochemistry-of-affective-disorders/1CB6D7B69D1E60F5731D4B8FBC02CE84[↩]
- Smith KA, Fairburn CG, Cowen PJ. Relapse of depression after rapid depletion of tryptophan. Lancet. 1997;349:915–9.[↩]
- Berman RM, Narasimhan M, Miller HL, et al. Transient depressive relapse induced by catecholamine depletion: potential phenotypic vulnerability marker? Arch Gen Psychiatry. 1999;56:395–403.[↩]
- Ruhé HG, Mason NS, Schene AH. Mood is indirectly related to serotonin, norepinephrine and dopamine levels in humans: a meta-analysis of monoamine depletion studies. Mol Psychiatry. 2007;12:331–59.[↩]
- Anderson IM, Parry-Billings M, Newsholme EA, et al. Decreased plasma tryptophan concentration in major depression: relationship to melancholia and weight loss. J Affect Disord. 1990;20:185–91.[↩]
- Wichers MC, Koek GH, Robaeys G, et al. IDO and interferon-α-induced depressive symptoms: a shift in hypothesis from tryptophan depletion to neurotoxicity. Mol Psychiatry. 2005;10:538–44.[↩]
- Uher R, Tansey KE, Dew T, et al. An inflammatory biomarker as a differential predictor of outcome of depression treatment with escitalopram and nortriptyline. Am J Psychiatry. 2014;171:1278–86.[↩]
- Sharp T, Cowen PJ. 5-HT and depression: is the glass half-full? Curr Opin Pharmacol. 2011;11:45–51.[↩]
- Harmer CJ, Goodwin GM, Cowen PJ. Why do antidepressants take so long to work? A cognitive neuropsychological model of antidepressant drug action. Br J Psychiatry. 2009;195:102–8.[↩][↩]
- Beck AT, Rush AJ, Shaw BF. Cognitive therapy of depression. New York: Guildford; 1979.[↩]
- Duman RS, et al. Pathophysiology of depression: the concept of synaptic plasticity. Eur Psychiatry. 2002;17:306–10.[↩]
- Perreau-Linck E, Beauregard M, Gravel P, et al. In vivo measurements of brain trapping of 11C-labelled α-methyl-L-tryptophan during acute changes in mood states. Journal of Psychiatry & Neuroscience : JPN. 2007;32(6):430-434. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2077345/[↩][↩]
- Kjaer TW, Bertelsen C, Piccini P, et al. Increased dopamine tone during meditation-induced change of consciousness. Brain Res Cogn Brain Res 2002;13:255-9. https://www.ncbi.nlm.nih.gov/pubmed/11958969[↩]
- erreau-Linck E, Beauregard M, Gravel P, et al. In vivo measurements of brain trapping of α-[11C]methyl-L-tryptophan during acute changes in mood states. J Psychiatry Neurosci 2007;32:430-4. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2077345/[↩]
- Golden RN, Gaynes BN, Ekstrom RD, et al. The efficacy of light therapy in the treatment of mood disorders: a review and meta-analysis of the evidence. Am J Psychiatry 2005;162:656-62. https://www.ncbi.nlm.nih.gov/pubmedhealth/PMH0021785/[↩]
- Lam RW, Carter D, Misri S, et al. A controlled study of light therapy in women with late luteal phase dysphoric disorder. Psychiatry Res 1999;86:185-92.[↩]
- Epperson CN, Terman M, Terman JS, et al. Randomized clinical trial of bright light therapy for antepartum depression: preliminary findings. J Clin Psychiatry 2004;65:421-5.[↩]
- Carlsson A, Svennerholm L, Winblad B. Seasonal and circadian monoamine variations in human brains examined post mortem. Acta Psychiatr Scand Suppl 1980;280:75-85.[↩]
- Lambert GW, Reid C, Kaye DM, et al. Effects of sunlight and season on serotonin turnover in the brain. Lancet 2002;360:1840-2.[↩]
- Cagampang FRA, Yamazaki S, Otori Y, et al. Serotonin in the raphe nuclei: regulation by light and an endogenous pacemaker. Neuroreport 1993;5:49-52[↩]
- Fite KV, Wu PS, Bellemer A. Photostimulation alters c-Fos expression in the dorsal raphe nucleus. Brain Res 2005;1031:245-52.[↩]
- aan het Rot M, Benkelfat C, Boivin DB, et al. Bright light exposure during acute tryptophan depletion prevents a lowering of mood in mildly seasonal women. Eur Neuropsychopharmacol. Epub 2007 Jun 18 ahead of print.[↩]
- aan het Rot M, Moskowitz DS, Young SN. Exposure to bright light is associated with positive social interaction and good mood over short time periods: a naturalistic study in mildly seasonal people. J Psychiatr Res DOI:10.1016/j.jpsychires.2006.11.010. Epub 2007 Jan 31 ahead of print.[↩]
- Light cafe to beat the winter blues. BBC News. Available: http://news.bbc.co.uk/2/hi/health/4595626.stm[↩]
- Leidig M. Austrian town uses mirrors to tackle lack of daylight. BMJ 2005;331:1162.[↩]
- Salmon P. Effects of physical exercise on anxiety, depression, and sensitivity to stress: a unifying theory. Clin Psychol Rev 2001;21:33-61.[↩]
- Depression. https://www.nice.org.uk/guidance/conditions-and-diseases/mental-health-and-behavioural-conditions/depression[↩]
- Salmon P. Effects of physical exercise on anxiety, depression, and sensitivity to stress: a unifying theory. Clin Psychol Rev 2001;21:33-61. https://www.ncbi.nlm.nih.gov/pubmed/11148895[↩]
- Toups M, Carmody T, Greer T, Rethorst C, Grannemann B, Trivedi MH. Exercise is an effective treatment for positive valence symptoms in major depression. Journal of affective disorders. 2017;209:188-194. doi:10.1016/j.jad.2016.08.058. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6036912/[↩][↩]
- Post RM, Goodwin FK. Simulated behavior states: An approach to specificity in psychobiological research. Biol Psychiatry 1973;7:237-54.[↩]
- Chaouloff F, Elghozi JL, Guezennec Y, et al. Effects of conditioned running on plasma, liver and brain tryptophan and on brain 5-hydroxytryptamine metabolism of the rat. Br J Pharmacol 1985;86:33-41.[↩]
- Gomez-Merino D, Béquet F, Berthelot M, et al. Site-dependent effects of an acute intensive exercise on extracellular 5-HT and 5-HIAA levels in rat brain. Neurosci Lett 2001;301:143-6.[↩]
- Jacobs BL, Fornal CA. Activity of serotonergic neurons in behaving animals. Neuropsychopharmacology 1999;21:9S-15S.[↩]
- Rueter LE, Jacobs BL. A microdialysis examination of serotonin release in the rat forebrain induced by behavioral/environmental manipulations. Brain Res 1996;739:57-69.[↩]
- Chaouloff F, Laude D, Guezennec Y, et al. Motor activity increases tryptophan, 5-hydroxyindoleacetic acid, and homovanillic acid in ventricular cerebrospinal fluid of the conscious rat. J Neurochem 1986;46:1313-6.[↩]
- Blomstrand E. Amino acids and central fatigue. Amino Acids 2001;20:25-34. https://www.ncbi.nlm.nih.gov/pubmed/11310928[↩][↩]
- Pardridge WM. Blood-brain barrier transport of nutrients. Nutr Rev 1986;44(Suppl):15-25. https://www.ncbi.nlm.nih.gov/pubmed/2980844[↩]
- Lambert KG. Rising rates of depression in today’s society: Consideration of the roles of effort-based rewards and enhanced resilience in day-to-day functioning. Neurosci Biobehav Rev 2006;30:497-510. https://www.ncbi.nlm.nih.gov/pubmed/16253328[↩]
- Kerem Z, Lev-Yadun S, Gopher A, et al. Chickpea domestication in the Neolithic Levant through the nutritional perspective. J Archaeol Sci 2007;34:1289-93.[↩][↩]
- Katz SH, Hediger ML, Valleroy LA. Traditional maize processing techniques in the new world: traditional alkali processing enhances the nutritional quality of the maize. Science 1974;184:765-73.[↩]
- Xue-Cun C, Tai-An Y, Xiu-Zhen T, et al. Opaque-2 maize in the prevention and treatment of pellagra. Nutr Res 1983;3:171-80.[↩]
- Morris CE, Sands DC. The breeder’s dilemma — yield or nutrition? Nat Biotechnol 2006;24:1078-80.[↩]
- Mawson AR, Jacobs KW. Corn consumption, tryptophan, and cross-national homicide rates. J Orthomolec Psychiatry 1978;7:227-30.[↩]
- Voracek M, Tran US. Dietary tryptophan intake and suicide rate in industrialized nations. J Affect Disord 2007;98:259-62. [↩]
- United States Department of Agriculture Agricultural Research Service. USDA Food Composition Databases. https://ndb.nal.usda.gov/ndb/search/list[↩]
- Richard D.M., Dawes M.A., Mathias C.W., Acheson A., Hill-Kapturczak N., Dougherty D.M. l-tryptophan: Basic metabolic functions, behavioral research and therapeutic indications. Int. J. Tryptophan Res. IJTR. 2009;2:45–60 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2908021/[↩]
- Chu A, Wadhwa R. Selective Serotonin Reuptake Inhibitors. [Updated 2023 May 1]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK554406[↩][↩][↩][↩][↩]
- Bruggeman C, O’Day CS. Selective Serotonin Reuptake Inhibitor Toxicity. [Updated 2023 Jul 4]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK534815[↩]
- Antosik-Wójcińska AZ, Stefanowski B, Święcicki Ł. Efficacy and safety of antidepressant’s use in the treatment of depressive episodes in bipolar disorder – review of research. Psychiatr Pol. 2015;49(6):1223-39. English, Polish. doi: 10.12740/PP/37914[↩][↩]
- Preskorn SH. Clinically relevant pharmacology of selective serotonin reuptake inhibitors. An overview with emphasis on pharmacokinetics and effects on oxidative drug metabolism. Clin Pharmacokinet. 1997;32 Suppl 1:1-21. doi: 10.2165/00003088-199700321-00003[↩][↩][↩][↩]
- Feighner JP. Mechanism of action of antidepressant medications. J Clin Psychiatry. 1999;60 Suppl 4:4-11; discussion 12-3. https://www.psychiatrist.com/pdf/mechanism-of-action-of-antidepressant-medications-pdf/[↩]
- Delgado PL. How antidepressants help depression: mechanisms of action and clinical response. J Clin Psychiatry. 2004;65 Suppl 4:25-30. https://www.psychiatrist.com/pdf/how-antidepressants-help-depression-mechanisms-of-action-and-clinical-response-pdf/[↩]
- Xue W, Wang P, Li B, Li Y, Xu X, Yang F, Yao X, Chen YZ, Xu F, Zhu F. Identification of the inhibitory mechanism of FDA approved selective serotonin reuptake inhibitors: an insight from molecular dynamics simulation study. Phys Chem Chem Phys. 2016 Jan 28;18(4):3260-71. doi: 10.1039/c5cp05771j[↩]
- DeLucia V, Kelsberg G, Safranek S. Which SSRIs most effectively treat depression in adolescents? J Fam Pract. 2016 Sep;65(9):632-4. https://www.mdedge.com/jfponline/article/111448/mental-health/which-ssris-most-effectively-treat-depression-adolescents[↩]
- Rohde C, Hieronymus F, Østergaard SD. A target trial emulation comparing the antidepressant effectiveness of selective serotonin reuptake inhibitors (SSRIs) highlighting the importance of patent-related confounding by indication. Acta Psychiatr Scand. 2024 Oct;150(4):198-208. doi: 10.1111/acps.13729[↩]
- Coleiro B, Marshall SE, Denton CP, Howell K, Blann A, Welsh KI, Black CM. Treatment of Raynaud’s phenomenon with the selective serotonin reuptake inhibitor fluoxetine. Rheumatology (Oxford). 2001 Sep;40(9):1038-43. doi: 10.1093/rheumatology/40.9.1038[↩]
- Fisher KA, Marwaha R. Paraphilia. [Updated 2023 Mar 6]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK554425[↩]
- Seto MC, Kingston DA, Bourget D. Assessment of the paraphilias. Psychiatr Clin North Am. 2014 Jun;37(2):149-61. doi: 10.1016/j.psc.2014.03.001[↩]
- Seto MC. Pedophilia. Annu Rev Clin Psychol. 2009;5:391-407. doi: 10.1146/annurev.clinpsy.032408.153618[↩]
- PODOLSKY E. Exhibitionism. J Natl Med Assoc. 1960 Sep;52(5):343-4. https://pmc.ncbi.nlm.nih.gov/articles/instance/2641763/pdf/jnma00699-0033.pdf[↩]
- Balon R. Lack of Clarity in the DSM-5 Criteria of Voyeuristic Disorder. J Sex Marital Ther. 2016 Jul 3;42(5):391-2. doi: 10.1080/0092623X.2016.1158760[↩]
- Palermo GB. The various faces of sadism. Int J Offender Ther Comp Criminol. 2013 Apr;57(4):399-401. doi: 10.1177/0306624X13480125[↩]
- Civitarese G. Masochism and its Rhythm. J Am Psychoanal Assoc. 2016 Oct;64(5):885-916. doi: 10.1177/0003065116674442[↩]
- Långström N. The DSM diagnostic criteria for exhibitionism, voyeurism, and frotteurism. Arch Sex Behav. 2010 Apr;39(2):317-24. doi: 10.1007/s10508-009-9577-4[↩]
- Kafka MP. The DSM diagnostic criteria for fetishism. Arch Sex Behav. 2010 Apr;39(2):357-62. doi: 10.1007/s10508-009-9558-7[↩]
- Anupama M, Gangadhar KH, Shetty VB, Dip PB. Transvestism as a Symptom: A Case Series. Indian J Psychol Med. 2016 Jan-Feb;38(1):78-80. doi: 10.4103/0253-7176.175131[↩]
- Susser LC, Sansone SA, Hermann AD. Selective serotonin reuptake inhibitors for depression in pregnancy. Am J Obstet Gynecol. 2016 Dec;215(6):722-730. doi: 10.1016/j.ajog.2016.07.011[↩][↩]
- Hirschfeld RM. Efficacy of SSRIs and newer antidepressants in severe depression: comparison with TCAs. J Clin Psychiatry. 1999 May;60(5):326-35. doi: 10.4088/jcp.v60n0511 https://www.psychiatrist.com/jcp/efficacy-ssris-newer-antidepressants-severe-depression[↩]
- Kocsis JH. Review: SSRIs and TCAs equally effective at treating chronic depression and dysthemia; SSRIs are associated with fewer adverse events than TCAs. Evid Based Ment Health. 2013 Aug;16(3):82. doi: 10.1136/eb-2013-101268[↩]
- David DJ, Gourion D. Antidépresseurs et tolérance : déterminants et prise en charge des principaux effets indésirables [Antidepressant and tolerance: Determinants and management of major side effects]. Encephale. 2016 Dec;42(6):553-561. French. doi: 10.1016/j.encep.2016.05.006[↩]
- Hu XH, Bull SA, Hunkeler EM, Ming E, Lee JY, Fireman B, Markson LE. Incidence and duration of side effects and those rated as bothersome with selective serotonin reuptake inhibitor treatment for depression: patient report versus physician estimate. J Clin Psychiatry. 2004 Jul;65(7):959-65. doi: 10.4088/jcp.v65n0712[↩]
- Funk KA, Bostwick JR. A comparison of the risk of QT prolongation among SSRIs. Ann Pharmacother. 2013 Oct;47(10):1330-41. doi: 10.1177/1060028013501994[↩]
- Beach SR, Kostis WJ, Celano CM, Januzzi JL, Ruskin JN, Noseworthy PA, Huffman JC. Meta-analysis of selective serotonin reuptake inhibitor-associated QTc prolongation. J Clin Psychiatry. 2014 May;75(5):e441-9. doi: 10.4088/JCP.13r08672[↩]
- Laporte S, Chapelle C, Caillet P, Beyens MN, Bellet F, Delavenne X, Mismetti P, Bertoletti L. Bleeding risk under selective serotonin reuptake inhibitor (SSRI) antidepressants: A meta-analysis of observational studies. Pharmacol Res. 2017 Apr;118:19-32. doi: 10.1016/j.phrs.2016.08.017[↩]
- Suicidality in Children and Adolescents Being Treated With Antidepressant Medications. https://www.fda.gov/drugs/postmarket-drug-safety-information-patients-and-providers/suicidality-children-and-adolescents-being-treated-antidepressant-medications[↩]
- Friedman RA. Antidepressants’ black-box warning–10 years later. N Engl J Med. 2014 Oct 30;371(18):1666-8. doi: 10.1056/NEJMp1408480[↩]
- Watson WA, Litovitz TL, Rodgers GC, Jr, Klein-Schwartz W, Youniss J, Rose SR, et al. 2002 annual report of the American Association of Poison Control Centers Toxic Exposure Surveillance System. Am J Emerg Med. 2003;21(5):353–421. http://www.ajemjournal.com/article/S0735-6757(03)00088-3/fulltext[↩][↩]
- Simon LV, Torrico TJ, Keenaghan M. Serotonin Syndrome. [Updated 2024 Mar 2]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK482377[↩][↩][↩][↩]
- Citalopram. https://medlineplus.gov/druginfo/meds/a699001.html[↩]
- Citalopram (oral route). https://www.mayoclinic.org/drugs-supplements/citalopram-oral-route/description/drg-20062980[↩]
- LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]. Bethesda (MD): National Institute of Diabetes and Digestive and Kidney Diseases; 2012-. Citalopram. [Updated 2017 Oct 12]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK548395[↩][↩]
- Citalopram | Escitalopram (Celexa® | Lexapro®). https://mothertobaby.org/fact-sheets/citalopramescitalopram-celexalexapro-pregnancy[↩][↩][↩][↩][↩][↩][↩][↩][↩][↩]
- Escitalopram. https://medlineplus.gov/druginfo/meds/a603005.html[↩]
- LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]. Bethesda (MD): National Institute of Diabetes and Digestive and Kidney Diseases; 2012-. Fluoxetine. [Updated 2018 Feb 2]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK548010[↩][↩]
- Fluoxetine. https://medlineplus.gov/druginfo/meds/a689006.html[↩]
- Fluoxetine (Prozac®). https://mothertobaby.org/fact-sheets/fluoxetine-prozac-pregnancy[↩][↩][↩][↩]
- Baudat M, de Kort AR, van den Hove DLA, Joosten EA. Early-life exposure to selective serotonin reuptake inhibitors: Long-term effects on pain and affective comorbidities. Eur J Neurosci. 2022 Jan;55(1):295-317. doi: 10.1111/ejn.15544[↩]
- LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]. Bethesda (MD): National Institute of Diabetes and Digestive and Kidney Diseases; 2012-. Paroxetine. [Updated 2020 Apr 8]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK548306[↩]
- Paroxetine. https://mothertobaby.org/fact-sheets/paroxetine-paxil-pregnancy/[↩][↩][↩]
- LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]. Bethesda (MD): National Institute of Diabetes and Digestive and Kidney Diseases; 2012-. Sertraline. [Updated 2020 Apr 8]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK548513[↩]
- LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]. Bethesda (MD): National Institute of Diabetes and Digestive and Kidney Diseases; 2012-. Vilazodone. [Updated 2020 Apr 8]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK548223[↩][↩]
- Vilazodone. https://medlineplus.gov/druginfo/meds/a611020.html[↩]
- Vilazodone (Viibryd®). https://mothertobaby.org/fact-sheets/vilazodone-viibryd[↩]
- Boyer EW, Shannon M. The serotonin syndrome. N Engl J Med. 2005;352(11):1112–20. Erratum in: N Engl J Med 2005;356(23):2437. http://www.nejm.org/doi/full/10.1056/NEJMra041867[↩][↩][↩]
- Watson WA, Litovitz TL, Rodgers GC, Jr, Klein-Schwartz W, Youniss J, Rose SR, et al. 2002 annual report of the American Association of Poison Control Centers Toxic Exposure Surveillance System. Am J Emerg Med. 2003;21(5):353–421. https://www.ncbi.nlm.nih.gov/pubmed/14523881[↩]
- Frank C. Recognition and treatment of serotonin syndrome. Canadian Family Physician. 2008;54(7):988-992. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2464814/[↩][↩][↩]
- Graudins A, Stearman A, Chan B. Treatment of the serotonin syndrome with cyproheptadine. J Emerg Med. 1998;16(4):615–9. https://www.ncbi.nlm.nih.gov/pubmed/9696181[↩]
- Boddy R, Ali R, Dowsett R. Use of sublingual olanzapine in serotonin syndrome. J Toxicol Clin Toxicol. 2004;42(5):725.[↩]
- Duggal HS, Fetchko J. Serotonin syndrome and atypical antipsychotics. Am J Psychiatry. 2002;159(4):672–3. https://www.ncbi.nlm.nih.gov/pubmed/11925312[↩]
- Nisijima K, Ishiguro T. Does dantrolene influence central dopamine and serotonin metabolism in the neuroleptic malignant syndrome? A retrospective study. Biol Psychiatry. 1993;33(1):45–8. https://www.ncbi.nlm.nih.gov/pubmed/7678377[↩]
- Scotton WJ, Hill LJ, Williams AC, Barnes NM. Serotonin Syndrome: Pathophysiology, Clinical Features, Management, and Potential Future Directions. Int J Tryptophan Res. 2019 Sep 9;12:1178646919873925. doi: 10.1177/1178646919873925[↩]
- Boyer EW, Shannon M. The serotonin syndrome. N Engl J Med. 2005 Mar 17;352(11):1112-20. doi: 10.1056/NEJMra041867. Erratum in: N Engl J Med. 2007 Jun 7;356(23):2437. Erratum in: N Engl J Med. 2009 Oct 22;361(17):1714[↩]



{kind=link}